
Gene Expression Analysis in Cold Stress Conditions Reveals BBX20 and CLO as Potential Biomarkers for Cold Tolerance in Almond

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cold Treatment
2.2. Phenotype Analysis of the Cold Damages
2.3. RNA Isolation and cDNA Synthesis
2.4. High-Throughput qRT-PCR
2.5. Statistical Analysis
3. Results and Discussion
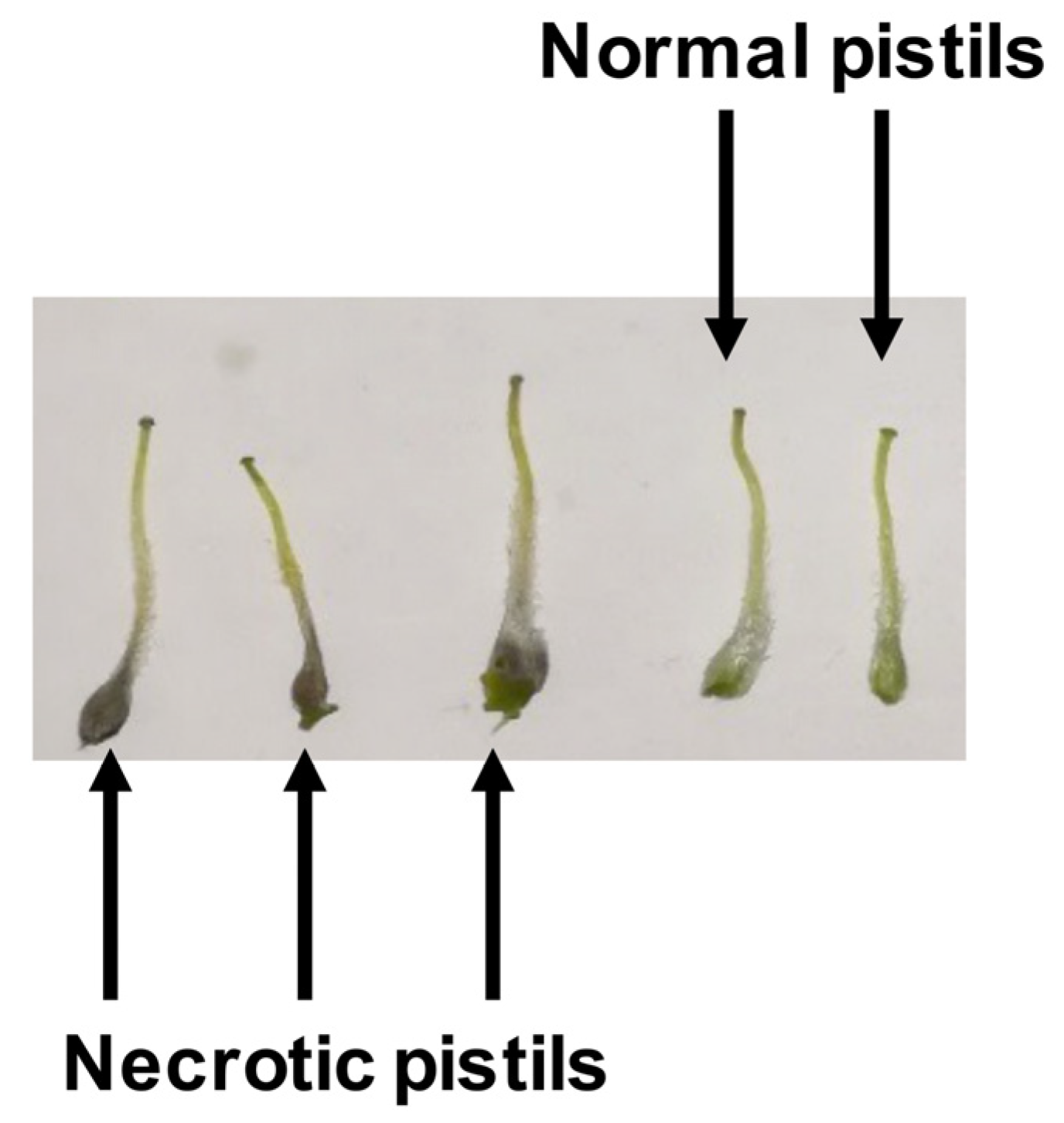
3.1. Necrotic Pistil as a Cold-Sensitiveness Phenotypic Trait
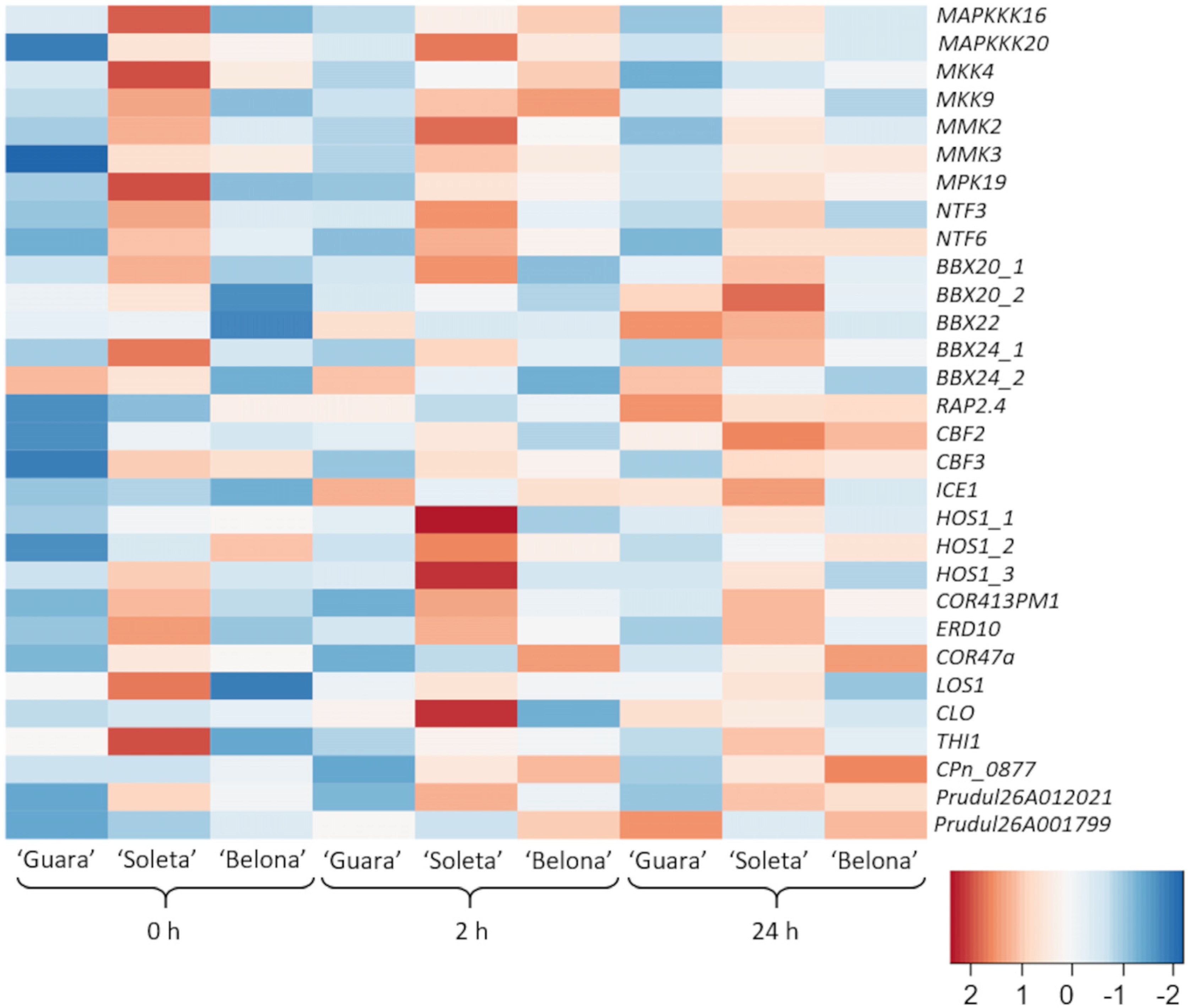
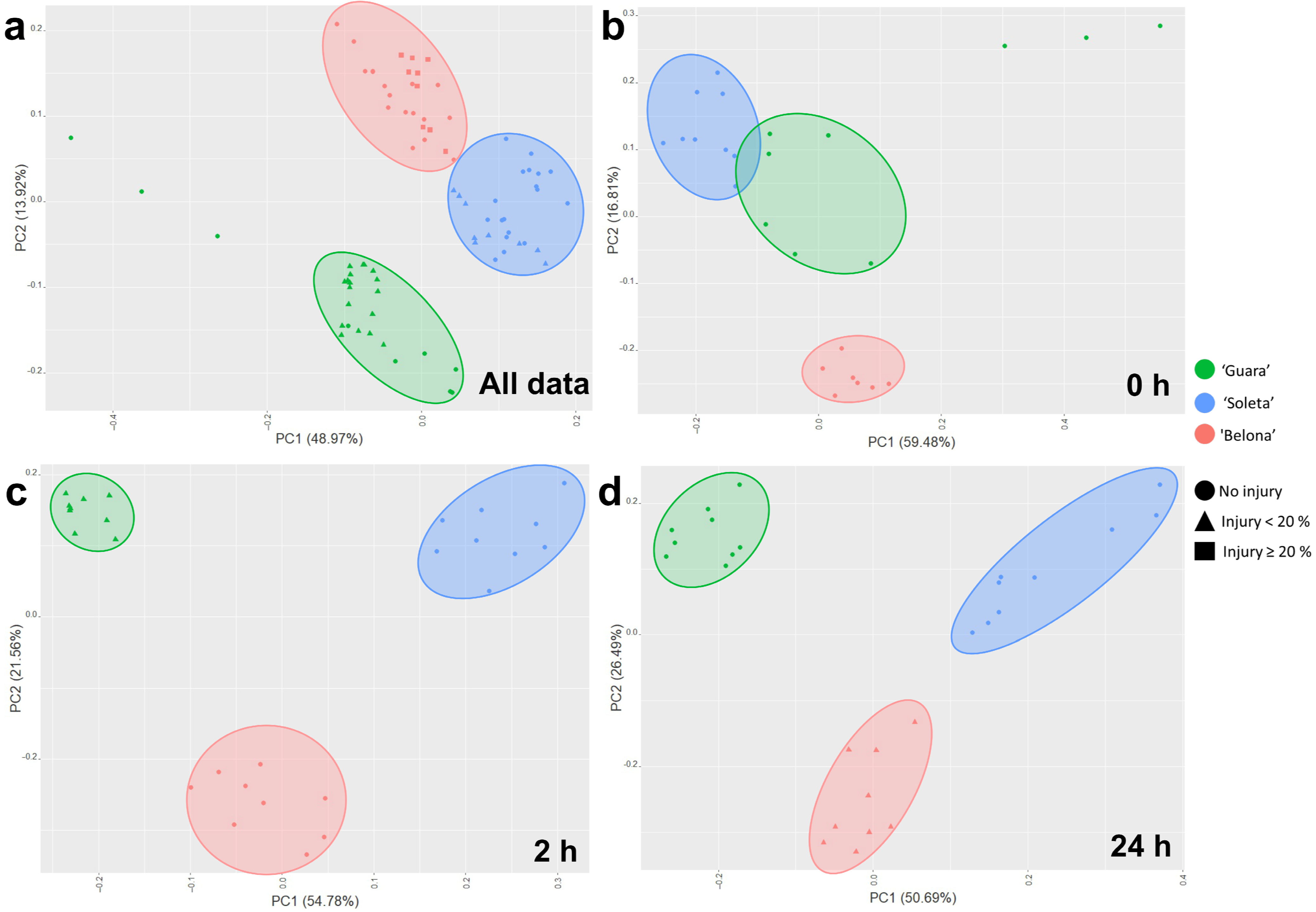
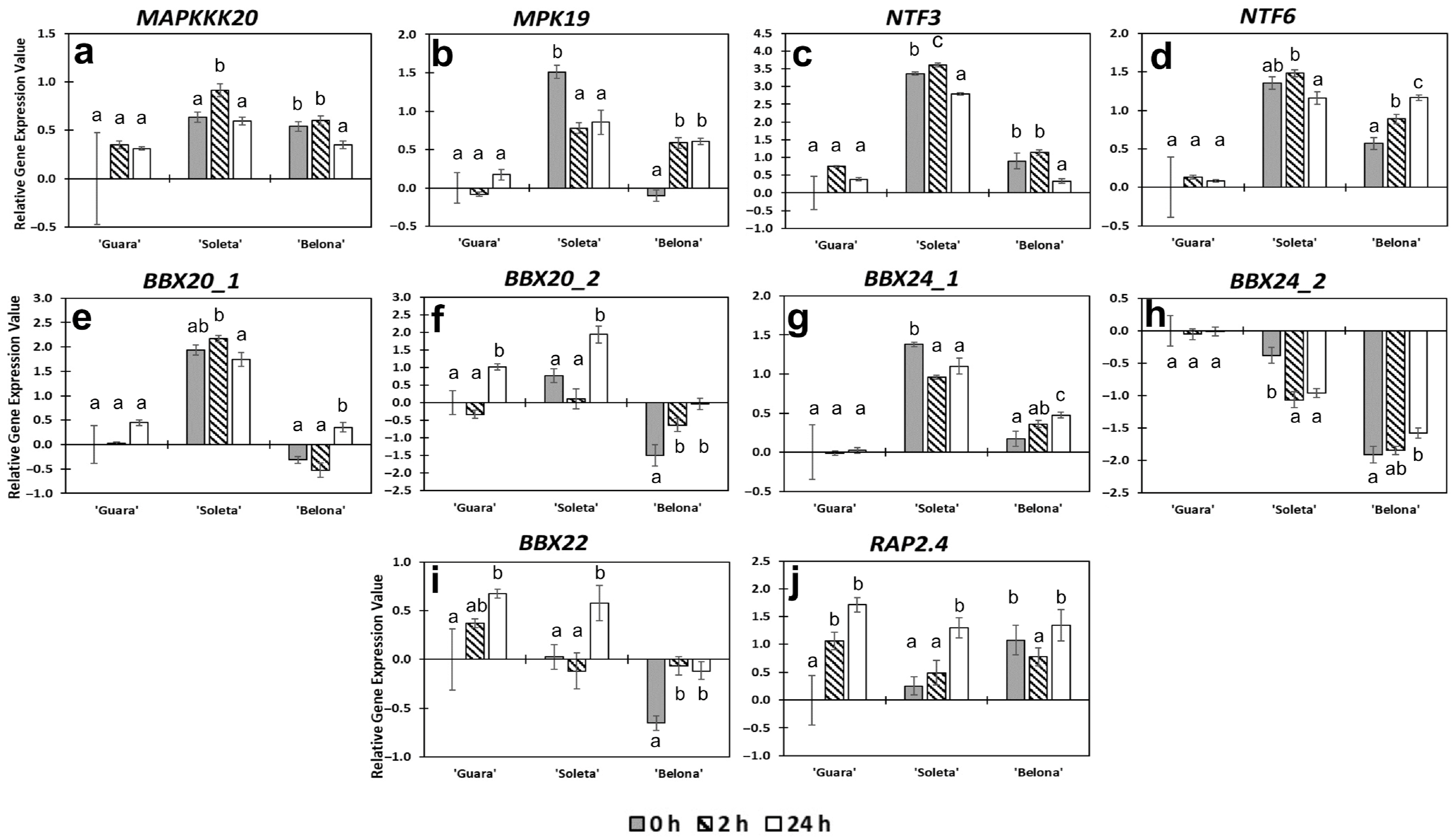
3.2. Changes in Gene Expression as a Response to Cold Stress
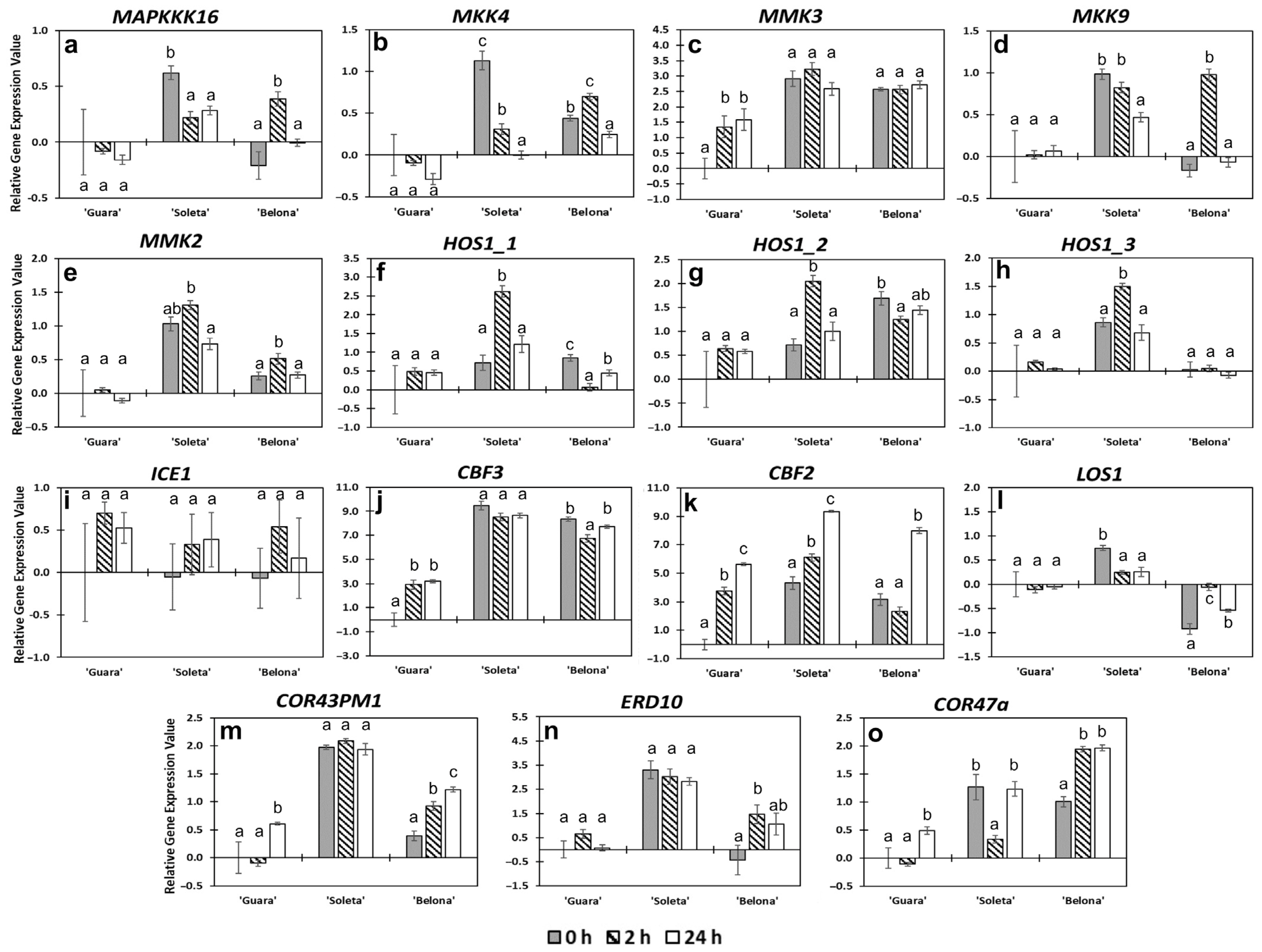
3.2.1. ICE-CBF-COR Pathway
3.2.2. CBF-Independent Responses
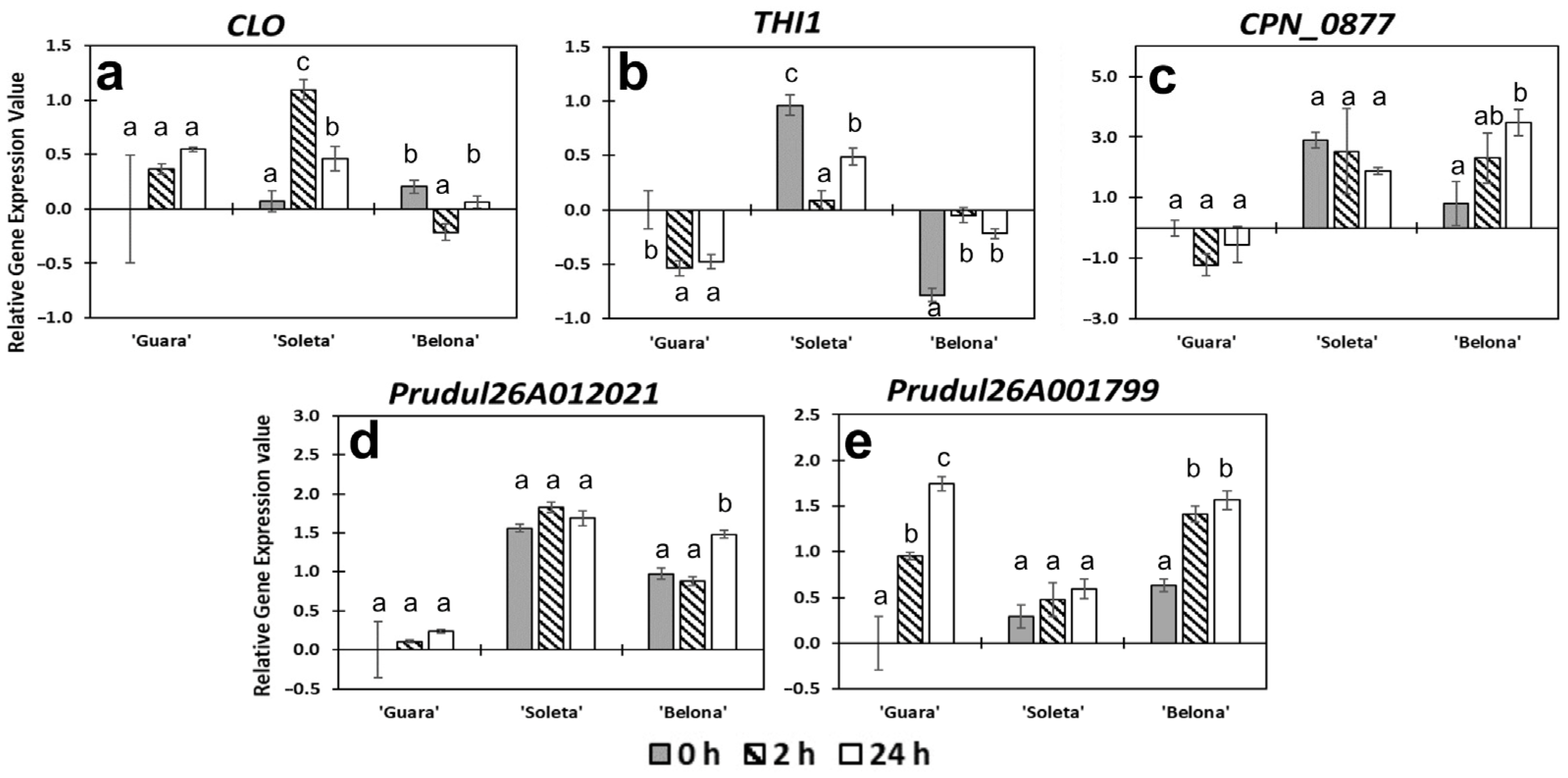
3.2.3. Other Functional and Uncharacterized Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Varela, S.A. Aspectos básicos de la fisiología en respuesta al estrés y el clima como condicionante del mismo en las plantas. INTA EEA Bariloche. Comun. Técnica 2010, 78, 1–23. [Google Scholar]
- Janská, A.; Maršík, P.; Zelenková, S.; Ovesná, J. Cold stress and acclimation—What is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Rodrigo, J. Spring frosts in deciduous fruit trees. Morphological damage and flower hardiness. Sci. Hortic. (Amsterdam) 2000, 85, 155–173. [Google Scholar] [CrossRef]
- Wisniewski, M.; Gusta, L.; Neuner, G. Adaptive mechanisms of freeze avoidance in plants: A brief update. Environ. Exp. Bot. 2014, 99, 133–140. [Google Scholar] [CrossRef]
- Lodolini, E.M.; Alfei, B.; Santinelli, A.; Cioccolanti, T.; Polverigiani, S.; Neri, D. Frost tolerance of 24 olive cultivars and subsequent vegetative re-sprouting as indication of recovery ability. Sci. Hortic. (Amsterdam) 2016, 211, 152–157. [Google Scholar] [CrossRef]
- Urbina, V. Daños por heladas en frutales. Sintomatología y evaluación. In Valoració de Danys Climatoògics i Incendis; Centre de Formació i Estudis Agrorurals, Generalitat de Catalunya—Departament d’Agricultura, Alimentació I Acció Rural: Reus, Spain, 2007; pp. 1–19. [Google Scholar]
- Yu, D.J.; Lee, H.J. Evaluation of freezing injury in temperate fruit trees. Hortic. Environ. Biotechnol. 2020, 61, 787–794. [Google Scholar] [CrossRef]
- Bañuelos, M.; Moreno, L.; Winzerling, J.; Orozco, J.; Gardea, A. Winter metabolism in deciduous trees: Mechanisms, genes and associated proteins. Rev. Fitotec. Mex. 2008, 31, 295–308. [Google Scholar]
- Alonso Segura, J.M.; Socias i Company, R.; Kodad, O. Late-blooming in almond: A controversial objective. Sci. Hortic. (Amsterdam) 2017, 224, 61–67. [Google Scholar] [CrossRef]
- Kodad, O.; Socias i Company, R. Daños diferenciales por heladas en flores y frutos y criterios de selección para la tolerancia a heladas en el almendro. ITEA Inf. Tec. Econ. Agrar. 2005, 101, 349–363. [Google Scholar]
- Lorite, I.J.; Cabezas-Luque, J.M.; Arquero, O.; Gabaldón-Leal, C.; Santos, C.; Rodríguez, A.; Ruiz-Ramos, M.; Lovera, M. The role of phenology in the climate change impacts and adaptation strategies for tree crops: A case study on almond orchards in Southern Europe. Agric. For. Meteorol. 2020, 294, 108142. [Google Scholar] [CrossRef]
- Felipe, J. Almendro. Estados fenológicos. Inf. Tec. Econ. Agrar. 1977, 27, 8–9. [Google Scholar]
- Alisoltani, A.; Shiran, B.; Fallahi, H.; Ebrahimie, E. Gene regulatory network in almond (Prunus dulcis Mill.) in response to frost stress. Tree Genet. Genomes 2015, 11, 1–15. [Google Scholar] [CrossRef]
- Meisel, L.; Fonseca, B.; González, S.; Baeza-Yates, R.; Cambiazo, V.; Campos, R.; González, M.; Orellana, A.; Retamales, J.; Silva, H. A Rapid and Efficient Method for Purifying High Quality Total RNA from Peaches (Prunus persica) for Functional Genomics Analyses. Biol. Res. 2005, 38, 83–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Salzman, R.A.; Fujita, T.; Zhu-Salzman, K.; Hasegawa, P.M.; Bressan, R.A. An Improved RNA Isolation Method for Plant Tissues Containing High Levels of Phenolic Compounds or Carbohydrates. Plant Mol. Biol. Rep. 1999, 17, 11–17. [Google Scholar] [CrossRef]
- Zeng, Y.; Yang, T. RNA Isolation From Highly Viscous Samples Rich in Polyphenols and Polysaccharides. Plant Mol. Biol. Rep. 2002, 20, 417. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Alioto, T.; Alexiou, K.G.; Bardil, A.; Barteri, F.; Castanera, R.; Cruz, F.; Dhingra, A.; Duval, H.; Martí, Á.F.I.; Frias, L.; et al. Transposons played a major role in the diversification between the closely related almond and peach genomes: Results from the almond genome sequence. Plant J. 2019, 101, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Team, R. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Zuther, E.; Lee, Y.P.; Erban, A.; Kopka, J.; Hincha, D.K. Natural variation in freezing tolerance and cold acclimation response in Arabidopsis thaliana and related species. In Survival Strategies in Extreme Cold and Desiccation. Advances in Experimental Medicine and Biology; Iwaya-Inoue, M., Sakurai, M., Uemura, M., Eds.; Springer: Singapore, 2018; Volume 1081, pp. 81–98. ISBN 9789811312441. [Google Scholar]
- Masip, D.; Torguet, L.; Batlle, I.; Alegre, S.; Miarnau, X. Almond fruit tolerance to frost temperatures in new Spanish cultivars. Acta Hortic. 2018, 1219, 67–72. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Paul, S.; Basu, S. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep. 2013, 32, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional Regulatory Networks in Cellular Responses and Tolerance to Dehydration and Cold Stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, M.M.; Hernández, P.F.V.; Rivas, M.R.; López, M.A.V.; Becerra, A.A.; Suárez, S.L.; Cárdenas, F.R. Resistencia y Tolerancia a Estrés Abiótico: Mecanismos Sofisticados de Adaptación de las Plantas ante Distintas Condiciones de Estrés. Available online: https://www.researchgate.net/profile/Pedro-Vera-Hernandez-2/publication/324482888_Resistencia_y_tolerancia_a_estres_abiotico_Mecanismos_sofisticados_de_adaptacion_de_las_plantas_ante_distintas_condiciones_de_estres/links/5ad62b86458515c60f55f5ff/Resistenc (accessed on 14 April 2021).
- Medina, J.; Catalá, R.; Salinas, J. The CBFs: Three arabidopsis transcription factors to cold acclimate. Plant Sci. 2011, 180, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.; Hong, X.; Agarwal, M.; Zhu, J.-K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.H.; Agarwal, M.; Zhang, Y.; Xie, Q.; Zhu, J.K. The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. USA 2006, 103, 8281–8286. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhang, L.; Dong, G.; Xu, Z.; Li, G.; Liu, N.; Wang, A.; Zhu, J. A novel cold-regulated protein isolated from Saussurea involucrata confers cold and drought tolerance in transgenic tobacco (Nicotiana tabacum). Plant Sci. 2019, 289, 110246. [Google Scholar] [CrossRef]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Wang, Y.; Liu, H.; Lei, L.; Yang, H.; Liu, G.; Ren, D. Activation of MAPK kinase 9 induces ethylene and camalexin biosynthesis and enhances sensitivity to salt stress in Arabidopsis. J. Biol. Chem. 2008, 283, 26996–27006. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP Kinase Cascades Regulate the Cold Response by Modulating ICE1 Protein Stability. Dev. Cell 2017, 43, 618–629. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, C.; Zhu, F.; Li, Y. Mild osmotic stress promotes 4-methoxy indolyl-3-methyl glucosinolate biosynthesis mediated by the MKK9–MPK3/MPK6 cascade in Arabidopsis. Plant Cell Rep. 2017, 36, 543–555. [Google Scholar] [CrossRef]
- Li, H.; Ding, Y.; Shi, Y.; Zhang, X.; Zhang, S.; Gong, Z.; Yang, S. MPK3- and MPK6-Mediated ICE1 Phosphorylation Negatively Regulates ICE1 Stability and Freezing Tolerance in Arabidopsis. Dev. Cell 2017, 43, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.G.; Jiang, W.; Chen, S.L.; Mantri, N.; Tao, Z.M.; Jiang, C.X. Insights from the cold transcriptome and metabolome of Dendrobium officinale: Global reprogramming of metabolic and gene regulation networks during cold acclimation. Front. Plant Sci. 2016, 7, 1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.; Raina, S.K.; Sultan, S.M. Arabidopsis MAPK signaling pathways and their cross talks in abiotic stress response. J. Plant Biochem. Biotechnol. 2020, 29, 700–714. [Google Scholar] [CrossRef]
- Wisniewski, M.; Nassuth, A.; Arora, R. Cold hardiness in trees: A mini-review. Front. Plant Sci. 2018, 9, 1394. [Google Scholar] [CrossRef] [PubMed]
- Barros, P.M.; Gonçalves, N.; Saibo, N.J.M.; Oliveira, M.M. Functional characterization of two almond C-repeat-binding factors involved in cold response. Tree Physiol. 2012, 32, 1113–1128. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhang, L.; Zhang, T.; Zhang, H.; Xu, S.; An, L. Transcriptional regulation network of cold-responsive genes in higher plants. Plant Sci. 2005, 169, 987–995. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Creelman, R.A.; Zhu, J.K. From laboratory to field. Using information from Arabidopsis to engineer salt, cold, and drought tolerance in crops. Plant Physiol. 2004, 135, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novillo, F.; Alonso, J.M.; Ecker, J.R.; Salinas, J. CBF2/DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3985–3990. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.G.; Seo, P.J. The MYB96-HHP module integrates cold and abscisic acid signaling to activate the CBF-COR pathway in Arabidopsis. Plant J. 2015, 82, 962–977. [Google Scholar] [CrossRef]
- Park, J.; Lim, C.J.; Shen, M.; Park, H.J.; Cha, J.Y.; Iniesto, E.; Rubio, V.; Mengiste, T.; Zhu, J.K.; Bressan, R.A.; et al. Epigenetic switch from repressive to permissive chromatin in response to cold stress. Proc. Natl. Acad. Sci. USA 2018, 115, E5400–E5409. [Google Scholar] [CrossRef] [Green Version]
- Breton, G.; Danyluk, J.; Frenette Charron, J.B.; Sarhan, F. Expression profiling and bioinformatic analyses of a novel stress-regulated multispanning transmembrane protein family from cereals and Arabidopsis. Plant Physiol. 2003, 132, 64–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.J.; Geng, J.C.; Miao, Y.J.; Xu, Y.M.; Hu, T.M.; Yang, P.Z. Transcriptomic analyses reveal genotype- and organ-specific molecular responses to cold stress in Elymus nutans. Biol. Plant. 2018, 62, 671–683. [Google Scholar] [CrossRef]
- Su, C.; Chen, K.; Ding, Q.; Mou, Y.; Yang, R.; Zhao, M.; Ma, B.; Xu, Z.; Ma, Y.; Pan, Y.; et al. Proteomic analysis of the function of a novel cold-regulated multispanning transmembrane protein COR413-PM1 in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Nam, K.H. Physiological roles of ERD10 in abiotic stresses and seed germination of Arabidopsis. Plant Cell Rep. 2010, 29, 203–209. [Google Scholar] [CrossRef]
- Deng, Z.; Pang, Y.; Kong, W.; Chen, Z.; Wang, X.; Liu, X.; Pi, Y.; Sun, X.; Tang, K. A novel ABA-dependent dehydrin ERD10 gene from Brassica napus. DNA Seq. J. DNA Seq. Mapp. 2005, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Artus, N.N.; Thomashow, M.F. cDNA sequence analysis and expression of two cold-regulated genes of Arabidopsis thaliana. Plant Mol. Biol. 1992, 18, 13–21. [Google Scholar] [CrossRef]
- Guo, Y.; Xiong, L.; Ishitani, M.; Zhu, J.-K. An Arabidopsis mutation in translation elongation factor 2 causes superinduction of CBF/DREB1 transcription factor genes but blocks the induction of their downstream targets under low temperatures. Proc. Natl. Acad. Sci. USA 2002, 99, 7786–7791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustafa, K.; AbuQamar, S.; Jarrar, M.; Al-Rajab, A.J.; Trémouillaux-Guiller, J. MAPK cascades and major abiotic stresses. Plant Cell Rep. 2014, 33, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Xiang, C. Bin The genetic locus At1g73660 encodes a putative MAPKKK and negatively regulates salt tolerance in Arabidopsis. Plant Mol. Biol. 2008, 67, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Woo, D.H.; Kim, S.H.; Lee, S.Y.; Park, H.Y.; Seok, H.Y.; Chung, W.S.; Moon, Y.H. Arabidopsis MKKK20 is involved in osmotic stress response via regulation of MPK6 activity. Plant Cell Rep. 2012, 31, 217–224. [Google Scholar] [CrossRef]
- Zhou, A.; Ma, H.; Liu, E.; Jiang, T.; Feng, S.; Gong, S.; Wang, J. Transcriptome sequencing of Dianthus spiculifolius and analysis of the genes involved in responses to combined cold and drought stress. Int. J. Mol. Sci. 2017, 18, 849. [Google Scholar] [CrossRef]
- Sun, X.; Xu, L.; Wang, Y.; Luo, X.; Zhu, X.; Kinuthia, K.B.; Nie, S.; Feng, H.; Li, C.; Liu, L. Transcriptome-based gene expression profiling identifies differentially expressed genes critical for salt stress response in radish (Raphanus sativus L.). Plant Cell Rep. 2016, 35, 329–346. [Google Scholar] [CrossRef]
- Wilson, C.; Eller, N.; Gartner, A.; Vicente, O.; Heberle-Bors, E. Isolation and characterization of a tobacco cDNA clone encoding a putative MAP kinase. Plant Mol. Biol. 1993, 23, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; Anglmayer, R.; Vicente, O.; Heberlebors, E. Molecular cloning, functional expression in Escherichia coli, and characterization of multiple mitogen-activated-protein kinases From tobacco. Eur. J. Biochem. 1995, 233, 249–257. [Google Scholar] [CrossRef]
- Reyna, N.S.; Yang, Y. Molecular analysis of the rice MAP kinase gene family in relation to Magnaporthe grisea infection. Mol. Plant-Microbe Interact. 2006, 19, 530–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Cheng, T.; Wang, G.; Yan, Y.; Xia, Q. Cloning and evolutionary analysis of tobacco MAPK gene family. Mol. Biol. Rep. 2013, 40, 1407–1415. [Google Scholar] [CrossRef]
- Wang, T.; Ye, C.; Wang, M.; Chu, G. Identification of cold-stress responsive proteins in Anabasis aphylla seedlings via the iTRAQ proteomics technique. J. Plant Interact. 2017, 12, 505–519. [Google Scholar] [CrossRef]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–471. [Google Scholar] [CrossRef]
- Shalmani, A.; Jing, X.Q.; Shi, Y.; Muhammad, I.; Zhou, M.R.; Wei, X.Y.; Chen, Q.Q.; Li, W.Q.; Liu, W.T.; Chen, K.M. Characterization of B-BOX gene family and their expression profiles under hormonal, abiotic and metal stresses in Poaceae plants. BMC Genom. 2019, 20, 27. [Google Scholar] [CrossRef]
- Wei, C.-Q.; Chien, C.-W.; Ai, L.-F.; Zhao, J.; Zhang, Z.; Li, K.; Burlingame, A.; Sun, Y.; Wang, Z.-Y. The Arabidopsis B-BOX Protein BZS1/BBX20 Interacts with HY5 and Mediates Strigolactone Regulation of Photomorphogenesis Chuang-Qi. J. Genet. Genom. 2016, 43, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.Y.; Sun, Y.; Cao, D.M.; Bai, M.Y.; Luo, X.M.; Yang, H.J.; Wei, C.Q.; Zhu, S.W.; Sun, Y.; Chong, K.; et al. BZS1, a B-box protein, promotes photomorphogenesis downstream of both brassinosteroid and light signaling pathways. Mol. Plant 2012, 5, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Dong, Y.; Yue, X.; Hu, J.; Jiang, S.; Xu, H.; Wang, Y.; Su, M.; Zhang, J.; Zhang, Z.; et al. The B-box zinc finger protein MdBBX20 integrates anthocyanin accumulation in response to ultraviolet radiation and low temperature. Plant Cell Environ. 2019, 42, 2090–2104. [Google Scholar] [CrossRef]
- Lin, R.C.; Park, H.J.; Wang, H.Y. Role of Arabidopsis RAP2.4 in regulating lightand ethylene-mediated developmental processes and drought stress tolerance. Mol. Plant 2008, 1, 42–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, N.; Takeda, S.; Matsumoto, N.; Okada, K. VAJ/GFA1/CLO is involved in the directional control of floral Organ growth. Plant Cell Physiol. 2009, 50, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Berger, F.; Twell, D. Germline specification and function in plants. Annu. Rev. Plant Biol. 2011, 62, 461–484. [Google Scholar] [CrossRef]
- Moll, C.; Von Lyncker, L.; Zimmermann, S.; Kägi, C.; Baumann, N.; Twell, D.; Grossniklaus, U.; Groß-Hardt, R. CLO/GFA1 and ATO are novel regulators of gametic cell fate in plants. Plant J. 2008, 56, 913–921. [Google Scholar] [CrossRef]
- Li, C.L.; Wang, M.; Wu, X.M.; Chen, D.H.; Lv, H.J.; Shen, J.L.; Qiao, Z.; Zhang, W. THI1, a Thiamine Thiazole Synthase, Interacts with Ca2+ -Dependent Protein Kinase CPK33 and Modulates the S-Type Anion Channels and Stomatal Closure in Arabidopsis. Plant Physiol. 2016, 170, 1090–1104. [Google Scholar] [CrossRef] [Green Version]
- Khosa, J.; Bellinazzo, F.; Goldstein, R.K.; Macknight, R.; Immink, R.G.H. PEBP proteins: The conductors of dual reproduction in plants with vegetative storage organs. J. Exp. Bot. 2021, 72, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Cui, F.; Hou, L.; Zhao, S.; Xia, H.; Qiu, J.; Li, T.; Zhang, Y.; Wang, X.; et al. Genome-wide analysis of gene expression provides new insights into cold responses in Thellungiella salsuginea. Front. Plant Sci. 2017, 8, 713. [Google Scholar] [CrossRef] [Green Version]
- de Silva, K.; Laska, B.; Brown, C.; Sederoff, H.W.; Khodakovskaya, M. Arabidopsis thaliana calcium-dependent lipid-binding protein (AtCLB): A novel repressor of abiotic stress response. J. Exp. Bot. 2011, 62, 2679–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campo, S.; Baldrich, P.; Messeguer, J.; Lalanne, E.; Coca, M.; San Segundo, B. Overexpression of a Calcium-Dependent Protein Kinase Confers Salt and Drought Tolerance in Rice by Preventing Membrane Lipid Peroxidation. Plant Physiol. 2014, 165, 688–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Peng, H.; Tan, S.; Zhou, J.; Fang, Z.; Hu, Z.; Gao, L.; Li, T.; Zhang, W.; Chen, L. Effects of early cold stress on gene expression in Chlamydomonas reinhardtii. Genomics 2020, 112, 1128–1138. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Percentage of Necrotic Pistils | ||
|---|---|---|---|
| 0 h | 2 h | 24 h | |
| ‘Guara’ | 0 A, a | 2 B, b | 7 A, c |
| ‘Soleta’ | 0 A, a | 0 A, a | 7 A, b |
| ‘Belona’ | 0 A, a | 0 A, a | 62 B, b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bielsa, B.; Ávila-Alonso, J.I.; Fernández i Martí, Á.; Grimplet, J.; Rubio-Cabetas, M.J. Gene Expression Analysis in Cold Stress Conditions Reveals BBX20 and CLO as Potential Biomarkers for Cold Tolerance in Almond. Horticulturae 2021, 7, 527. https://doi.org/10.3390/horticulturae7120527
Bielsa B, Ávila-Alonso JI, Fernández i Martí Á, Grimplet J, Rubio-Cabetas MJ. Gene Expression Analysis in Cold Stress Conditions Reveals BBX20 and CLO as Potential Biomarkers for Cold Tolerance in Almond. Horticulturae. 2021; 7(12):527. https://doi.org/10.3390/horticulturae7120527
Chicago/Turabian StyleBielsa, Beatriz, Jorge Israel Ávila-Alonso, Ángel Fernández i Martí, Jérôme Grimplet, and María José Rubio-Cabetas. 2021. "Gene Expression Analysis in Cold Stress Conditions Reveals BBX20 and CLO as Potential Biomarkers for Cold Tolerance in Almond" Horticulturae 7, no. 12: 527. https://doi.org/10.3390/horticulturae7120527
APA StyleBielsa, B., Ávila-Alonso, J. I., Fernández i Martí, Á., Grimplet, J., & Rubio-Cabetas, M. J. (2021). Gene Expression Analysis in Cold Stress Conditions Reveals BBX20 and CLO as Potential Biomarkers for Cold Tolerance in Almond. Horticulturae, 7(12), 527. https://doi.org/10.3390/horticulturae7120527