
Photosynthetic and Ultrastructural Properties of Eggplant (Solanum melongena) under Salinity Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Phenotyping
2.3. Data Analysis
3. Results
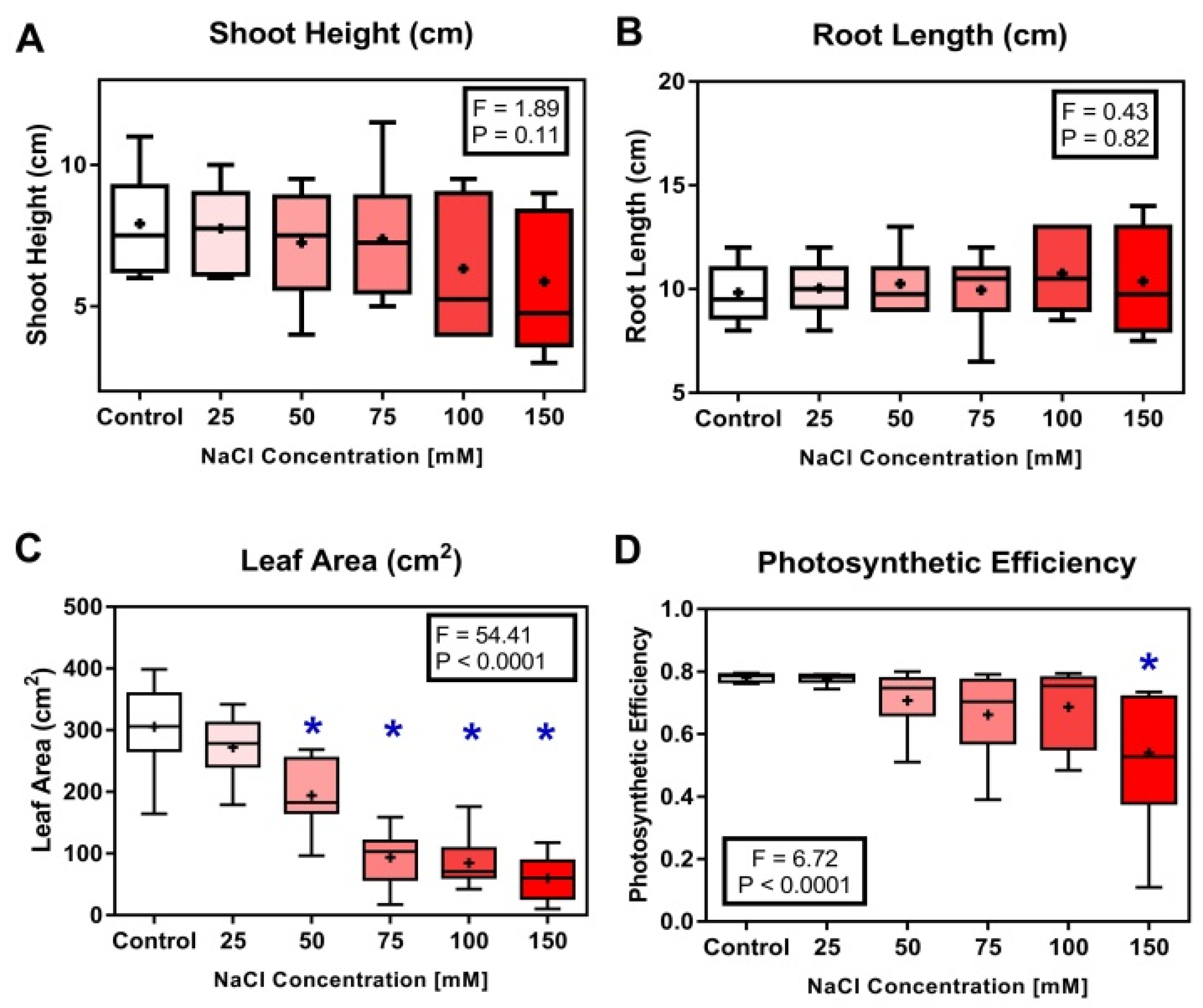
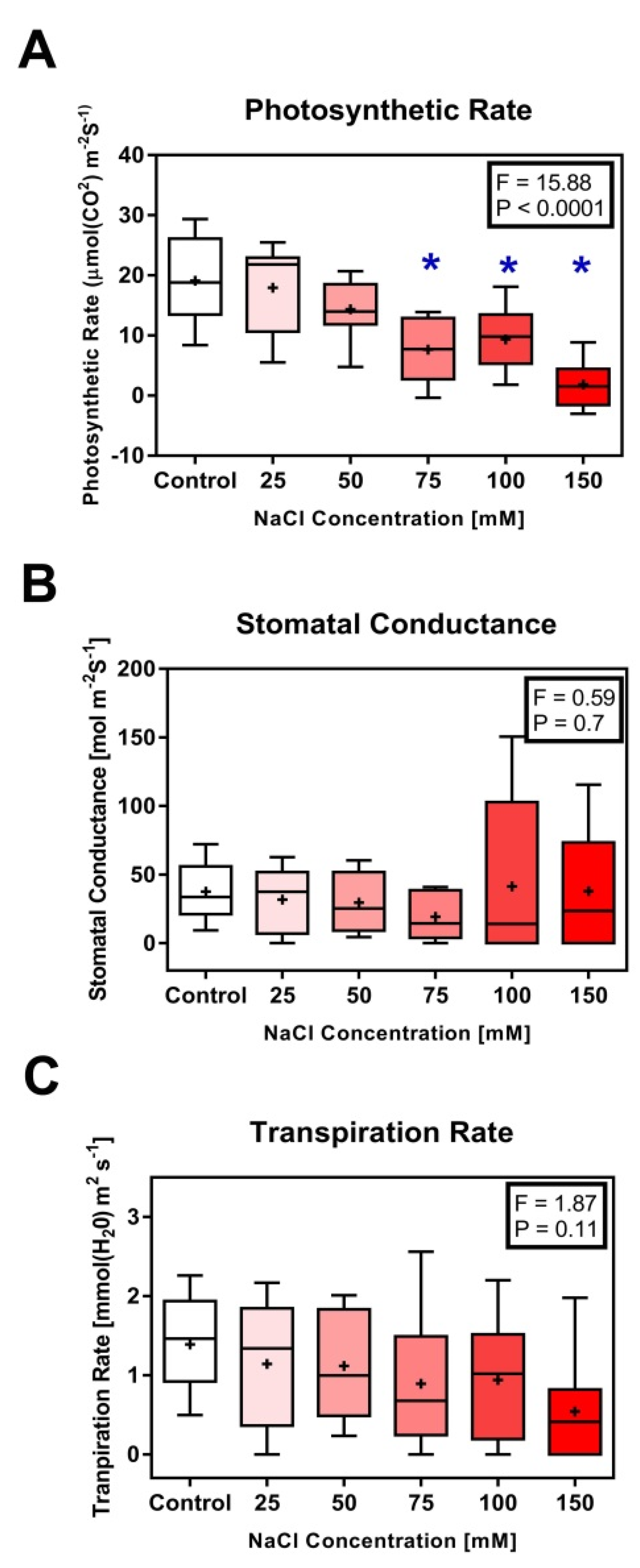
3.1. Morphological and Physiological Characteristics
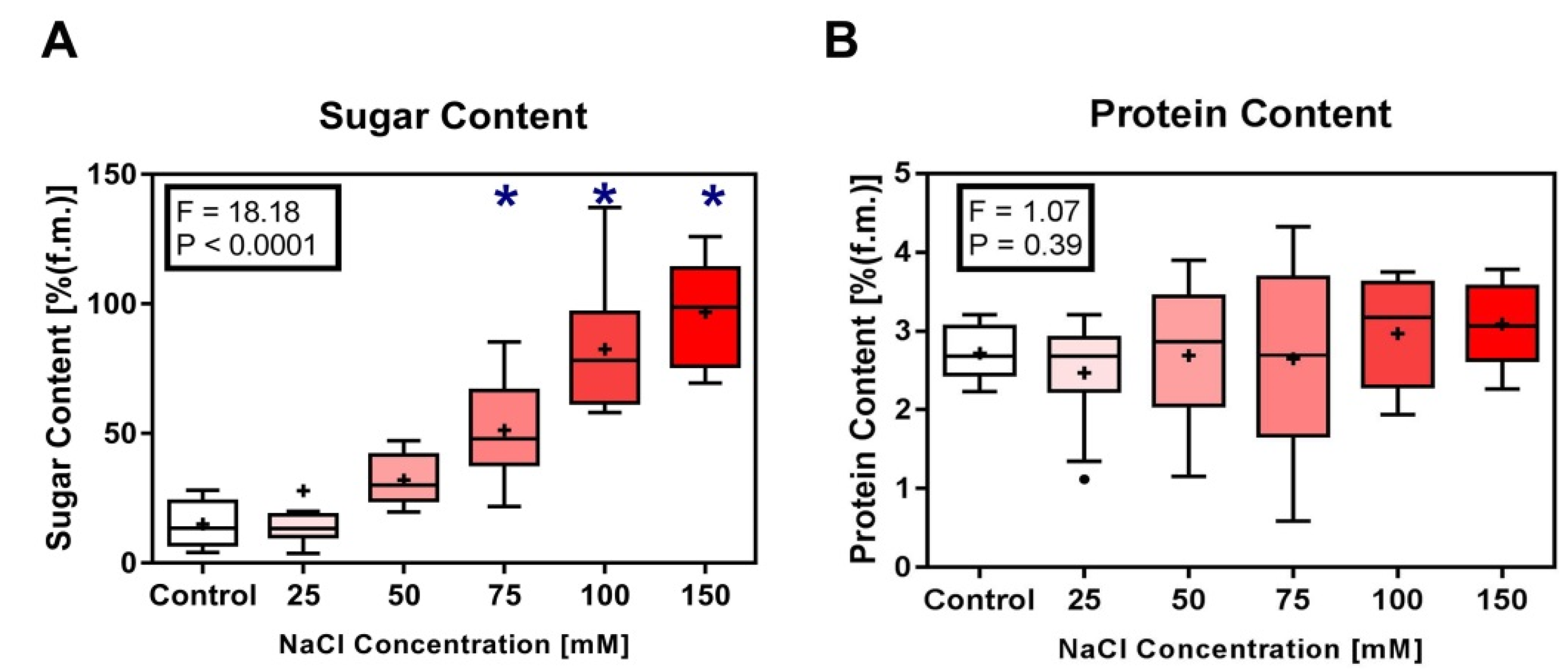
3.2. Biochemical Characteristics
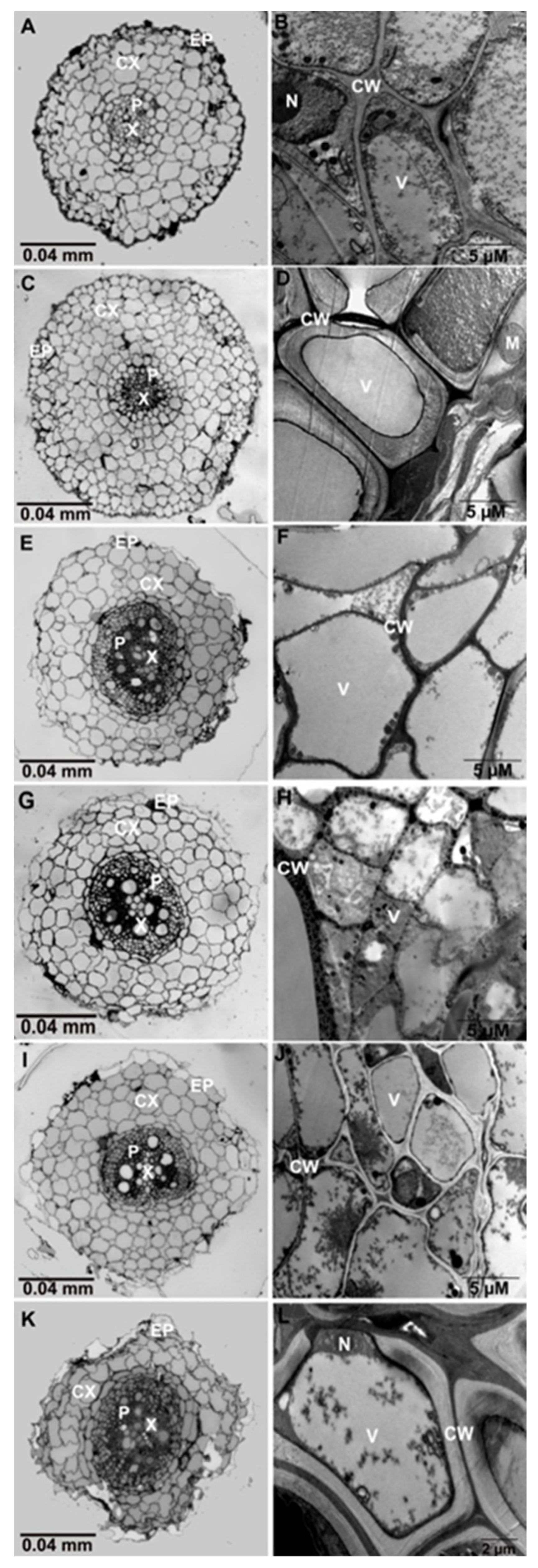
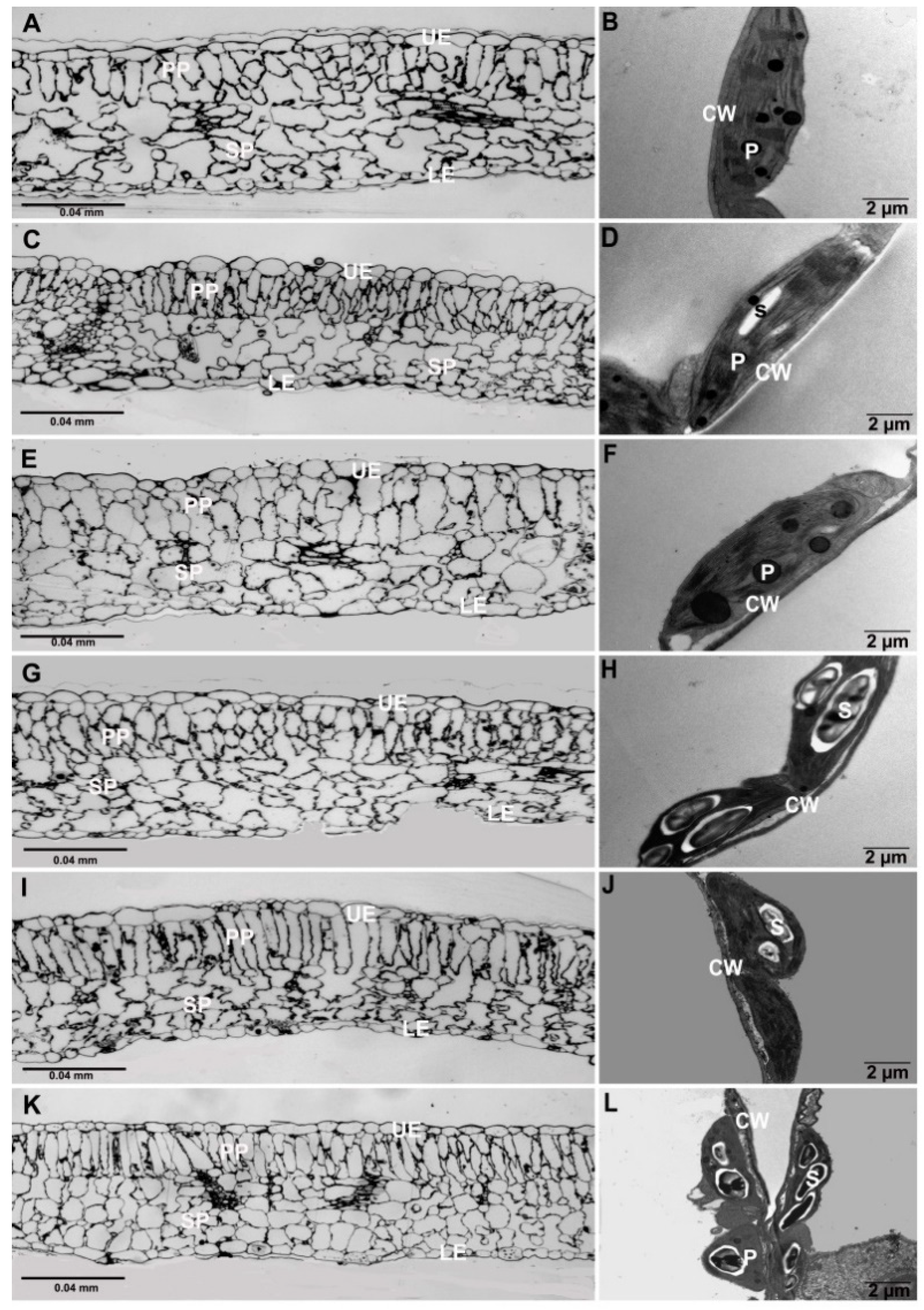
3.3. Microscopic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Unlukara, A.; Kurunc, A.; Semiz, D.G.; Yurtseven, E.; Suarez, L.D. Effects of salinity on eggplant (Solanum melongena L.) growth and evapotranspiration. Irrig. Drain. 2010, 59, 203–214. [Google Scholar]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, P.; Kalaji, M.H.; Baczewska, A.H.; Pawluśkiewicz, B.; Mastalerczuk, G.; Borawska-Jarmułowicz, B.; Paunov, M.; Goltsev, V. Delayed chlorophyll a fluorescence, MR 820, and gas exchange changes in perennial ryegrass under salt stress. J. Lumin. 2017, 183, 322–333. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Al-Qurainy, F.; Harris, P.J.C. Salt tolerance in selected vegetable crops. Crit. Rev. Plant Sci. 2012, 31, 303–320. [Google Scholar] [CrossRef]
- Queiroz, I.S.R.; Leitão, A.R.F.; Ferreira, L.L.; Dias, N.S.; Cosme, C.R.; Mota, A.F. Tolerance of the eggplant salinity cultivated in coconut fiber substrate. Agropecuária Científica No Semi-Árido 2013, 9, 15–20. [Google Scholar]
- Ashraf, M. Some important physiological selection criteria for salt tolerance in plants. Flora 2004, 199, 361–376. [Google Scholar] [CrossRef]
- She, D.; Sun, X.; Gamareldawla, A.H.D.; Nazar, E.A.; Hu, W.; Edith, K.; Yu, S. Benefits of soil biochar amendments to tomato growth under saline water irrigation. Sci. Rep. 2018, 8, 14743. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi. J. Biol. Sci. 2014, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbaspour, H. Effect of salt stress on lipid peroxidation, antioxidative enzymes, and proline accumulation in pistachio plants. J. Med. Plants Res. 2012, 6, 526–529. [Google Scholar]
- Akinci, E.I.; Akinci, S.; Yilmaz, K.; Dikici, H. Response of eggplant varieties (Solanum melongena) to salinity in germination and seedling stages. New Zealand J. Crop Hortic. Sci. 2004, 32, 193–200. [Google Scholar] [CrossRef]
- Nasri, N.; Kaddour, R.; Rebhi, M.; Plassard, C.; Lachal, M. Effect of salinity on germination, phytase activity and phytate content in lettuce seedling. Acta Physiol. Plant. 2011, 33, 935–994. [Google Scholar] [CrossRef]
- Cuatero, J.; Fernandez-Munoz, R. Tomato and salinity. Sci. Hortic. 1999, 78, 83–125. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, S.; Saha, B.; Awasthi, J.; Dey, M.; Panda, S.K.; Sahoo, L. Crosstalk between salt, drought, and cold stress in plants:toward genetic engineering for stress tolerance. In Abiotic Stress Response in Plants: Tuteja/Abiotic Stress Response in Plants; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 55–86. [Google Scholar]
- Turkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Hannachi, S.; Labeke, M.V. Salt stress affects germination, seedling growth and physiological responses differentially in eggplant cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Zhu, X.; Cao, Q.; Sun, L.; Yang, X.; Yang, W.; Zhang, H. Stomatal conductance and morphology of arbuscular mycorrhizal wheat plants response to elevated CO2 and NaCl stress. Front. Plant Sci. 2018, 9, 1363. [Google Scholar] [CrossRef] [PubMed]
- Eller, F.; Lambertini, C.; Nguyen, L.X.; Brix, H. Increased invasive potential of non-native Phragmites australis: Elevated CO2 and temperature alleviate salinity effects on photosynthesis and growth. Glob. Chang. Biol. 2014, 20, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.J.; Sun, L.H.; Fan, N.L.; Yang, Z.M.; Huang, B.R. Physiological factors involved in positive effects of elevated carbon dioxide concentration on Bermuda grass tolerance to salinity stress. Environ. Exp. Bot. 2015, 115, 20–27. [Google Scholar] [CrossRef]
- Oliveira, F.S.; Sá, F.V.S.; Souto, L.S.; Paiva, E.P.; Oliveira, F.A.; Araújo, E.B.G.; Neto, H.T.O.; Mesquita, E.F. Seedling of development and tolerance of eggplant cultivars under saline stress. Afr. J. Agric. Res. 2016, 11, 2310–2315. [Google Scholar]
- Gonçalves, M.C.R.; Diniz, M.F.M.; Dantas, A.H.G.; Borba, J.R.C. Modesto efeito hipolipemiante do extrato seco de berinjela (Solanum melongena L.) em mulheres dislepidemias, sob controle nutricional. Rev. Bras. Farm. 2006, 16, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Abbas, W.; Ashraf, M.; Akram, N.A. Alleviation of salt-induced adverse effects in eggplant (Solanum melongena L.) by glycinebetaine and sugarbeet extracts. Sci. Hortic. 2010, 125, 188–195. [Google Scholar] [CrossRef]
- Alkhatib, R.; Alkhatib, B.; AL-Eitan, L.; Abdo, N.; Tadros, M.; Bsoul, E. Physio-anatomical responses of tobacco under caffeine stress. Photosynthetica 2018, 56, 1140–1146. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parthiban, M.; Thilagavathi, G.; Viju, S. Effect of Solanum tuberosum on Muslin-Gauze Composite for Healthcare Applications. J. Text. Sci. Eng. 2012, 2, 121. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alkhatib, R.; Alkhatib, B.; Abdo, N.; AL-Eitan, L.; Creamer, R. Physio-biochemical and ultrastructural impact of (Fe3O4) nanoparticles on tobacco. BMC Plant Biol. 2019, 19, 253–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, M.A.; Holt, S.J. The localization by electron microscopy of HeLa cell surface enzymes splitting adenosine triphosphate. J. Cell Biol. 1963, 19, 325–336. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Safdarian, M.; Askari, H.; Shariati, J.V.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Wen-Yuan, W.; Xiao-Feng, Y.; Ying, J.; Bo, Q.; Yu-Feng, X. Effects of salt stress on water content and photosynthetic characteristics in Iris lacteal var. Chinensis seedlings. Middle East J. Sci. Res. 2012, 12, 70–74. [Google Scholar]
- Zuo, Z.Y.; Ye, F.; Wang, Z.S.; Li, S.X.; Li, H.; Guo, J.H.; Mao, H.P.; Zhu, X.C.; Li, X.N. Salt acclimation induced salt tolerance in wild-type and chlorophyl b-deficient mutant wheat. Plant Soil Environ. 2021, 67, 26–32. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Sci. J. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- Fu, Q.S.; Yang, R.C.; Wang, H.S.; Zhao, B.; Zhou, C.L.; Ren, S.X.; Guo, Y.D. Leaf morphological and ultrastructural performance of eggplant (Solanum melongena L.) in response to water stress. Photosynthetica 2013, 51, 109–114. [Google Scholar] [CrossRef]
- Shaheen, S.; Naseer, S.; Ashraf, M.; Akram, N.A. Salt stress affects water relations, photosynthesis, and oxidative defense mechanisms in Solanum melongena L. J. Plant Interact. 2013, 8, 85–96. [Google Scholar] [CrossRef]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci. 2004, 44, 806–811. [Google Scholar] [CrossRef]
- Faseela, P.; Sinisha, A.K.; Brestič, M.; Puthur, J.T. Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice. Photosynthetica 2019, 57, 108–115. [Google Scholar]
- Daoud, A.M.; Hemada, M.M.; Saber, N.; El-Araby, A.A.; Moussa, L. Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water. Plants 2018, 7, 29. [Google Scholar]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical and molecular characterization. Int. J. Genom. 2014, 2014, 1–18. [Google Scholar] [CrossRef]
- Ibrahimova, U.; Zivcak, M.; Gasparovic, K.; Rastogi, A.; Allakhverdiev, S.I.; Yang, X.; Brestic, M. Electron and proton transport in wheat exposed to salt stress: Is the increase of the thylakoid membrane proton conductivity responsible for decreasing the photosynthetic activity in sensitive genotypes? Photosynth. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A.; Staden, J.V. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Pattanagul, W.; Thitisaksakul, M. Effect of salinity stress on growth and carbohydrate metabolism in three rice (Oryza sativa L.) cultivars differing in salinity tolerance. Indian J. Exp. Biol. 2008, 46, 736–742. [Google Scholar] [PubMed]
- Koch, K.E. Carbohydrate-modulated gene expression in plants. Annu. Rev. Plant Biol. 1996, 47, 509–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.; Akram, N.A.; Arteca, R.N.; Foolad, M.R. The physiological, biochemical and molecular roles of brassinosteroids and salicylic acid in plant processes and salt tolerance. Crit. Rev. Plant Sci. 2010, 29, 162–190. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Muhling, K.H.; Läuchli, A.; Brown, P.H.; Goldbach, H.E. The interaction between salinity and boron toxicity affects the subcellular distribution of ions and proteins in wheat leaves. Plant Cell Environ. 2003, 26, 1267–1274. [Google Scholar] [CrossRef]
- Ashraf, M.; O’Leary, J.W. Changes in soluble proteins in spring wheat stressed with sodium chloride. Biol. Plant. 1999, 42, 113–117. [Google Scholar] [CrossRef]
- Hajibagheri, M.; Harvey, D.M.; Flowers, T.J. Quantitative ion distribution withir/root cells of salt-sensitive and salt-tolerant maize varieties. New Phytol. 1987, 105, 367–379. [Google Scholar] [CrossRef]
- Xie, Z.; Song, R.; Shao, H.; Song, F.; Xu, H.; Lu, Y. Silicon improves maize photosynthesis in saline-alkaline soils. Sci. World J. 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Jubany-Marí, T.; Alegre, L. Drought induced senescence is characterized by a loss of antioxidant defences in chloroplasts. Plant Cell Environ. 2001, 24, 1319–1327. [Google Scholar] [CrossRef]
- Oi, T.; Enomoto, S.; Nakao, T.; Arai, S.; Yamane, K.; Taniguchi, M. Three-dimensional ultrastructural change of chloroplasts in rice mesophyll cells responding to salt stress. Ann. Bot. 2020, 125, 833–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhatib, R.; Abdo, N.; Mheidat, M. Photosynthetic and Ultrastructural Properties of Eggplant (Solanum melongena) under Salinity Stress. Horticulturae 2021, 7, 181. https://doi.org/10.3390/horticulturae7070181
Alkhatib R, Abdo N, Mheidat M. Photosynthetic and Ultrastructural Properties of Eggplant (Solanum melongena) under Salinity Stress. Horticulturae. 2021; 7(7):181. https://doi.org/10.3390/horticulturae7070181
Chicago/Turabian StyleAlkhatib, Rami, Nour Abdo, and Maha Mheidat. 2021. "Photosynthetic and Ultrastructural Properties of Eggplant (Solanum melongena) under Salinity Stress" Horticulturae 7, no. 7: 181. https://doi.org/10.3390/horticulturae7070181
APA StyleAlkhatib, R., Abdo, N., & Mheidat, M. (2021). Photosynthetic and Ultrastructural Properties of Eggplant (Solanum melongena) under Salinity Stress. Horticulturae, 7(7), 181. https://doi.org/10.3390/horticulturae7070181