Abstract
Breeding for disease resistance has been one of the most important research objectives in melon for the last few decades. Fusarium wilt caused by Fusarium oxysporum f. sp. melonis (Fom) is among the most threatening melon fungal diseases along the Mediterranean coast, affecting yield and quality. Since genetic resistance is one of the best sustainable strategies that can be used to control this pathogen, 27 Tunisian melon accessions collected from local farmers have been tested using phenotypic and molecular approaches to identify new sources of resistance to be used directly as cultivars in affected areas or as resistance donors in breeding programs. The phenotypic evaluations, using artificial inoculations, showed several resistant accessions to the pathogenic races of Fom. Additionally, molecular analysis revealed that 13 out of 27 accessions carried the resistance Fom-1 gene (7 in homozygous state and 6 in heterozygous state), confirming their resistance to races 0 and 2. Two of them were also identified as heterozygous for the Fom-2 gene, being resistant to races 0 and 1. Furthermore, two accessions with a high level of resistance to the most virulent race 1.2 have been also reported. This melon germplasm should be explored as a potential source of resistance genes in breeding programs to develop new resistant melon cultivars.
1. Introduction
Several fungal diseases, such as Fusarium wilt, powdery mildew, Monosporascus root rot, and charcoal root, caused by Macrophomina phaseolina [1,2,3,4,5,6,7,8], affect seriously melon crops (Cucumis melo L.), decreasing yield and fruit quality and forcing intensive use of chemical treatments. Among the diseases affecting melon crops, Fusarium wilt is one of the most threatening in the Mediterranean area [1]. Considerable damages caused by Fusarium wilt have been reported in many parts of this area, including Tunisia. The loss of yield through plant death and the decrease in marketable fruits are considered as the most obvious damages. The world annual loss in melon crops caused by Fusarium wilt has been estimated at 10–30%, and sometimes up to 100% [1].
Fusarium wilt is caused by several soil-borne fungal species and formae speciales belonging to the genus Fusarium [2,3,4,5]. Fusarium oxysporum f. sp. melonis W.C. Snyder and H.N. Hansen (Fom) is the best-known species affecting melon crops, and the four pathogenic races of this fungus (0, 1, 2, and 1.2) [9] have all been found in the Mediterranean basin, including Tunisia [1,6]. Fusarium wilt is one of the most difficult diseases to control primarily because the pathogen has a soil-borne lifestyle and remains viable in the soil as chlamydospores for decades. Agricultural practices have been used to control Fusarium wilt, such as crop rotation that has only provided limited protection, soil disinfection using chloropicrin or methyl bromide, grafting of melon onto resistant rootstocks [7], soil solarization with polyethylene sheet [10], and use of nonpathogenic strains of Fusarium to compete with pathogenic isolates. However, the most effective way to control the disease is the use of varieties with genetic resistance. This control measure also helps to reduce the large number of chemicals normally used in melon crop production, being favorable for the environment and human health.
Risser et al. [9] described two race-specific resistant genes, Fom-1 and Fom-2 in melon lines ‘Doublon’ and ‘CM17187’, respectively. Fom-1 confers resistance to races 0 and 2, whereas Fom-2 determines resistance to races 0 and 1. Oumouloud et al. [11] reported a new recessive gene fom-4 in a Spanish melon accession ‘Tortuga’ that confers resistance to races 0 and 2. Fom isolates classified as race 1.2 are able to cause disease in melon lines carrying the resistance genes described above. Race 1.2 is further divided into pathotype 1.2y, which induces leaf yellowing symptoms before the death of the plants, and 1.2w, which causes wilting and death without prior yellowing symptoms. The harmful race 1.2 has spread around the Mediterranean area and has become a problem for melon cultivation [1,6,12]; the development of genotypes with resistance not only to race 1.2 but to the other races of the pathogen represents a major objective in melon breeding programs. Resistance to race 1.2 seems to be quantitative and polygenically inherited [13,14,15]. Herman and Perl-Treves [16] reported a near-complete resistance to Fom race 1.2 in the melon breeding line BIZ, controlled by two complementary recessive genes, fom 1.2a and fom 1.2b. Chikh-Rouhou et al. [13] reported that resistance to race 1.2 is under complex genetic control since relatively moderate broad-sense heritabilities were obtained and epistatic effects were detected.
Several methods of screening to identify resistance sources have been used, artificial inoculations being the most suitable [17,18]. Combining artificial inoculations with the use of molecular markers would allow the identification of reliable sources of resistance. Several molecular markers linked to the two dominant Fom-1 and Fom-2 genes have been identified [19,20,21,22,23]. Those markers have only application in the specific melon genetic backgrounds where they were developed, and they could not be validated for consistent polymorphism in the Tunisian melon cultivars [20,21]. Oumouloud et al. [24,25] developed functional markers for Fom-1 and Fom-2 based on single nucleotide polymorphism within the coding region of the Fom-1 locus encoding for resistance to Fom and a functional nucleotide polymorphism detected between the susceptible and resistant Fom-2 alleles, respectively. These functional markers [24,25] are highly predictive of the phenotype as they target the functional polymorphism within a desired gene, overcoming the problem of recombination and linkage between marker and trait [26]. Regarding Fom 1.2, Perchepied et al. [27] identified a total of 9 recessive QTLs, associated with resistance, located on 5 linkage different groups, and Herman et al. [28] detected a major QTL for such resistance. However, useful molecular markers for Fom 1.2 race are not available yet.
There are commercial cultivars with resistance to races 0, 1, and 2, mostly belonging to ‘Galia’ and ‘Cantaloup’ types, but only a few hybrids and lines of makuwa and conomon botanical groups are partially resistant to race 1.2, which are mainly used as rootstocks. The spread of Fom races has become a problem for melon cultivation, and the development of genotypes with resistance to multiple races of the pathogen represents a major objective. Furthermore, the search for and utilization of new genes different from the well-known genes conferring resistance to Fusarium wilt are still among the primary objectives in melon breeding in order to strengthen the existing resistances and develop more durable resistant cultivars.
Local landraces represent a valuable genetic resource for breeding in a changing environment. They exhibit fine adaptation to the specific environment in which they have evolved under years of domestication. The Tunisian melon germplasm, unexplored to date for Fusarium wilt resistance, could be of great potential and should be exploited. The agronomic traits and fruit characteristics of the Tunisian accessions are similar to the commercial types demanded by Mediterranean markets, and they also carry the adaptation to the environment and cultivation methods [29,30,31]. All these characteristics would favor the development of varieties or hybrids of commercial interest if any resistance is found in the Tunisian material. The aim of this study was to screen Tunisian germplasm unexplored to date for Fusarium wilt resistance to identify new sources useful in breeding programs by using both phenotypic and molecular marker approaches.
2. Materials and Methods
2.1. Plant Material
The melon material used in this study consisted of 27 Tunisian melon landraces belonging to different horticultural groups, collected from local farmers of the Center-East of the country since 2015 and 2016 (Table 1) and maintained at the Regional Research Centre on Horticulture and Organic Agriculture (CRRHAB, Chott-Mariem, Tunisia). Most accessions have been characterized for their morpho-agronomic traits [29,31].

Table 1.
Horticultural groups and Tunisian melon landraces used in the study.
The melon genotypes ‘Charentais Fom-1’ and ‘Charentais Fom-2’ were used as controls resistant to races 0 and 2 and 0 and 1, respectively, and as controls susceptible to race 1.2. ‘Charentais-T’ was used as a control susceptible to races 0, 1, and 2. The resistant control used for race 1.2 was the hybrid F1 ‘Dinero’ (Syngenta), which is resistant to both pathotypes of race 1.2.
2.2. Phenotypic Evaluation of the Resistance to Fom
In this study, five Fom isolates were used, Fom0122 belonging to race 0, Fom0123 belonging to race 1, Fom0124 belonging to race 2, and Fom24mls and Fom0125 belonging to the pathotypes 1.2y and 1.2w, respectively. All strains were isolated and characterized by our group in previous works [11,12] and kept in the living collection (freeze-dried or deep-frozen at −85 °C) of the Plant Protection Unit of CITA (Zaragoza, Spain).
All Fom races were previously refreshed from lyophilized vials and grown in plates containing potato dextrose agar (PDA) medium (Panreac, Barcelona, Spain) at 25 °C for 5 days in the dark (Figure 1). Then, for each Fom race, a liquid culture containing yeast malt broth (per liter: 5 g of yeast extract (Difco Laboratories, Detroit, MI, USA), 5 g of malt extract (Panreac), 5 g of glucose (Panreac)) inoculated with 3 mycelial plugs (coming from the margins of 5-day-old PDA cultures) was prepared and incubated at room temperature under agitation (120 rpm) with a 16:8h (light/dark) photoperiod. After 3 days of incubation, the fermentation broth was filtered through an autoclaved nylon mesh, and the final conidial concentration of all isolates was determined using a Neubauer hemocytometer (Brand, Wertheim, Germany) and adjusted to 106 conidia/mL by diluting with sterile distilled water.
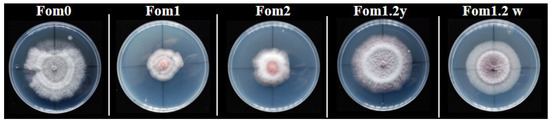
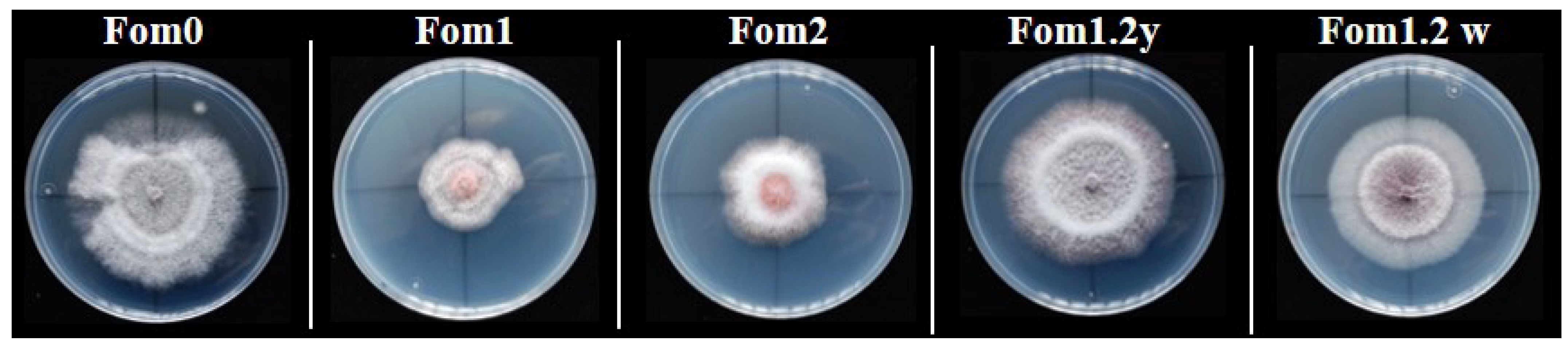
Figure 1.
Isolates of Fusarium oxysporum f.sp. melonis (Fom) races grown on PDA medium at 25 °C for 5 days in the dark.
Seeds of the melon accessions were sown into sterilized sand and grown in a growth chamber at 24 °C for 16 h per day (light) and 16 °C for 8 h per day (dark). At the first true leaf stage, (approximately 15-day-old) seedlings were carefully up-rooted from sand and their roots were washed in tap water. For each melon accession and Fom race, artificial inoculation was carried out by dipping the roots, previously trimmed at 0.5 cm, in conidial suspension (106 conidia/mL) of the appropriate race for 15 min [18]. The inoculated seedlings were transplanted directly into plastic pots containing sterilized substrate (1 part each of soil, sand, and peat) and maintained in a controlled growth chamber, at 28–20 °C (day/night) with a 16:8 h (light/dark) photoperiod, for 1 month. For each isolate inoculation assay, the plants were distributed according to a completely randomized design, with each plant constituting a replicate. For each combination of melon accession (including 27 Tunisian landraces and resistant and susceptible control genotypes) and Fom race, 12 plants were inoculated and another 12 plants per accession were included as uninoculated controls. At the end of this period, the presence of any Fusarium wilt symptom was noted to classify each accession as resistant or susceptible. To evaluate the performance against Fom races, plants were scored for disease severity based on a visual scale ranging from 0 to 4 (0 = no symptoms, 1 = beginning of wilting or yellowing on leaves, 2 = leaves heavily affected by wilting or yellowing, 3 = all leaves completely wilted, and 4 = death of the plant) [11,15,33] (Figure 2). In addition, all plants were also longitudinally cut to observe the presence of necrosis in internal vascular tissue; if the internal tissue was necrosed, the plant was recorded as susceptible [34].

Figure 2.
Typical symptoms of Fusarium wilt disease in melon seedlings (A) symptoms of wilting and (B) symptoms of yellowing. Scale used for symptom evaluation: 0 = no symptoms; 1 = beginning of wilting or yellowing on leaves; 2 = leaves heavily affected by wilting or yellowing; 3 = all leaves completely wilted or yellowed; 4 = death of the plant.
For each accession and Fom race, the mean and standard error of disease severity score (n = 12) were calculated. Melon accessions with a mean disease severity score between 0 and 1 (healthy plants) were classified as resistant (R), whereas those >1 were considered as susceptible (S) [33].
For Fom race 1.2 inoculation experiments, disease severity in each inoculated plant was evaluated at 7, 15, 21, and 30 dpi (days post-inoculation) according to the same visual scale from 0 to 4, and the area under the disease progress curve (AUDPC) was estimated from those scores [13,15]. The AUDPC integrates both the intensity of symptoms and the time taken between inoculation and expression of symptoms. The AUDPC was calculated for each treatment according to the formula AUDPC = ∑i [(xi +xi+1)/2] (ti+1 − ti), where i = 1 to 3 scorings, xi = mean disease score of each plant at date i, xi+1 = mean disease score of each plant at date i+1, and ti+1− ti = number of days between scoring date i and scoring date i + 1.
For the statistical analysis, the values of the AUDPC for each pathotype (1.2y and 1.2w) were used for the analysis of variance (ANOVA). Means were separated using the Tukey’s b test (p < 0.05). Data analysis was performed with the Statistical Analysis System (SAS Institute, Cary, NC, USA).
For races 0, 1, and 2, verification of the phenotypic data evaluation was conducted using molecular markers linked to disease resistance genes Fom-1 and Fom-2. Since there are no available useful molecular markers linked to resistance genes against Fom race 1.2 (pathotypes 1.2w and 1.2y), two artificial inoculation assays were conducted to confirm the resistance found in the accessions.
2.3. Analyses of Molecular Markers for Fom Races 0, 1, and 2
Fresh leaf tissue (2–3 g) at 1–2 true leaf stage was collected from each accession and ground in liquid nitrogen. Genomic DNA was extracted following a modified CTAB method [35]. DNA was quantified using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). For PCR amplification, the DNA concentrations were adjusted to a final concentration of 10 ng/mL with MTE Buffer (10 mM of Tris–HCl, pH 8.0; 1 mM EDTA).
A cleaved amplified polymorphic sequence (CAPS) marker, previously reported to be linked to Fom-1 gene [24], and two SCAR markers, previously reported to be linked to Fom-2 gene [25], were used (Table 2). The utility and robustness of these functional markers have been confirmed [36,37], and they resulted in good identification of the genotypes tested since they permit the distinction between the resistant and susceptible alleles.
Table 2.
Molecular markers, linked to Fom-1 and Fom-2 locus in melon, used in our study.
PCR reactions for Fom-1 and Fom-2 markers were carried out in a total volume of 10 µL containing 40 ng of DNA, 1× PCR buffer (20 mM of Tris–HCl pH 8.4 + 50 mM of KCl), 2.5 mM MgCl2, 400 µM of dNTP mixture in 2.5 mM, 0.1 µM of each primer, and 0.3U of Taq polymerase (Invitrogen, Carlsbad, CA, USA). PCR conditions were 2 min at 94 °C; 30 cycles of 94 °C for 45 s, 62 °C for 1 min for Fom-2 markers or 51 °C for 1 min for Fom-1 marker, and 72 °C for 2 min; and a final extension at 72 °C for 10 min.
Amplification products for Fom-1 PCR reaction were digested with BspCNI restriction endonuclease [24], resulting in 182 and 386 bp fragments in the resistant homozygous genotypes; three fragments, 182, 386, and 568 bp, in the resistant heterozygous genotypes; and uncut 568 bp fragment in the susceptible ones. For Fom-2 PCR reactions, the expected amplification products were 408 bp for the resistant genotypes and 342 bp for the susceptible ones. Amplified fragments were separated in a 1% agarose gel (Sigma Aldrich, St. Louis, MO, USA) in 1× TAE buffer stained with SYBR Safe DNA Gel Stain (Invitrogen, Carlsbad, CA, USA) and visualized using a UV transilluminator G-Box (Syngene, Cambridge, UK).
The melon genotypes ‘Charentais Fom-1’ and ‘Charentais Fom-2’ were used as resistant controls for Fom-1 and Fom-2 genes, respectively. ‘Charentais T’ was used as susceptible control for both genes.
3. Results
3.1. Phenotypic and Molecular Evaluation of the Resistance to Fom Races 0, 1, and 2
Fifteen Tunisian accessions showed resistance to race 0, 3 accessions were resistant to race 1, and 13 accessions were resistant to race 2 (Table 3; Supplementary Material). Typical symptoms of Fusarium wilt were observed in all inoculated plants of the susceptible control ‘Charentais T’ (Figure 3).
Table 3.
Phenotypic evaluation of the resistance to Fom (races 0, 1, and 2) and further molecular marker analysis for Fom-1 and Fom-2 genes in the Tunisian melon accessions.

Figure 3.
Symptom evaluation of yellowing caused by Fusarium wilt disease in some melon accessions inoculated with Fom0124 isolate belonging to race 2 in comparison to the susceptible and resistant controls.
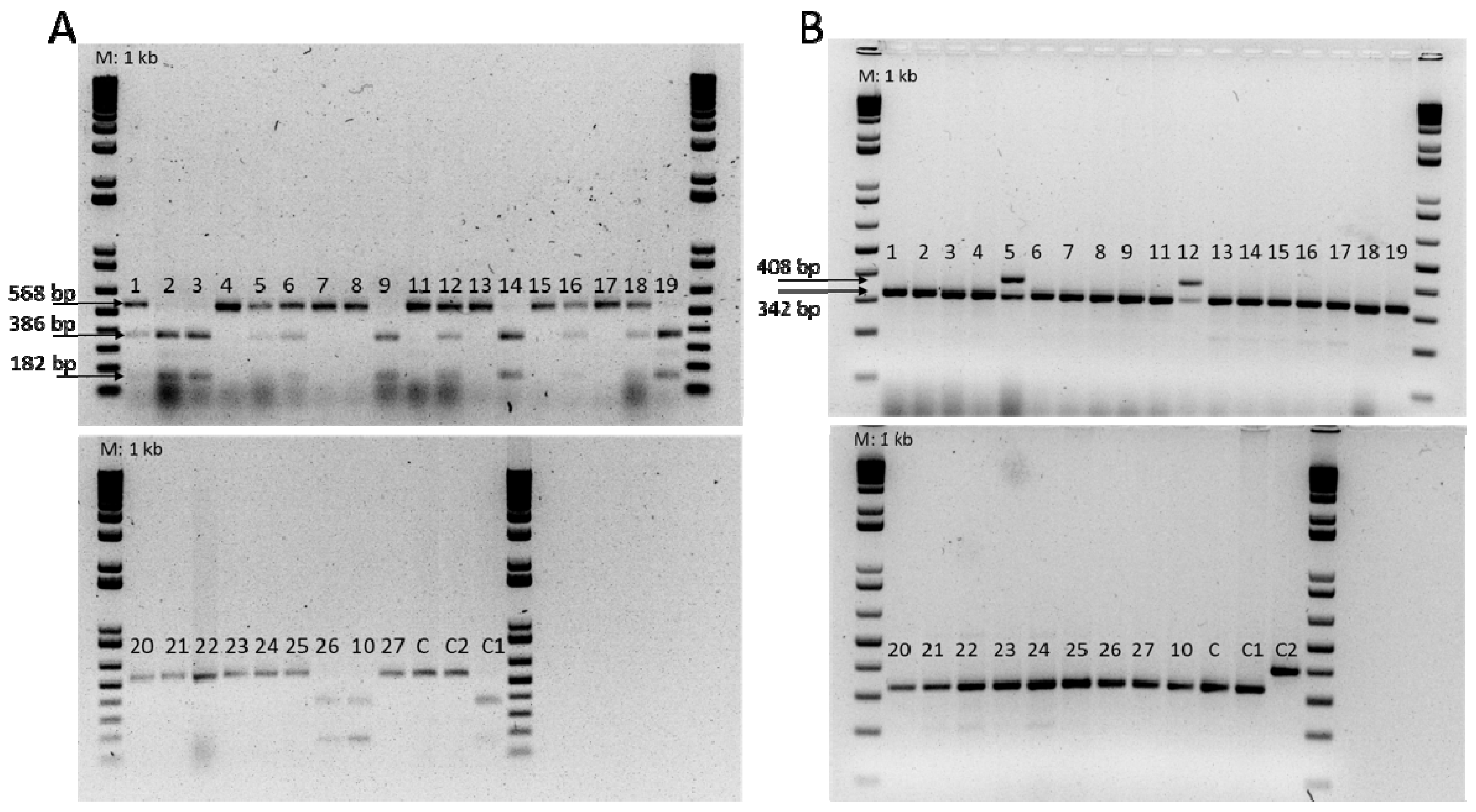
All accessions were then screened for the presence of alleles conferring resistance to Fom. The Fom-1 CAPS marker conditioned the presence (resistant) or absence (susceptible) of the BspCNI restriction endonuclease CTCAG(N)10 site (Figure 4A). The 13 accessions scored as resistant to race 0 and 2 by artificial inoculation contained the Fom-1 resistance gene, including 7 homozygous accessions (TUN-2, TUN-3, TUN-9, TUN-10, TUN-14, TUN-19, and TUN-26) with patterns of 182 and 386 bp digestion bands and 6 heterozygous accessions (TUN-1, TUN-5, TUN-6, TUN-12, TUN-16, and TUN-18) showing three digestion fragments of 182, 386, and 568 bp (Figure 4A). The remaining susceptible accessions showed an uncut 568 bp fragment, indicating the presence of the susceptible alleles of the Fom-1 gene and therefore, confirming the susceptibility to race 2 (Table 3).
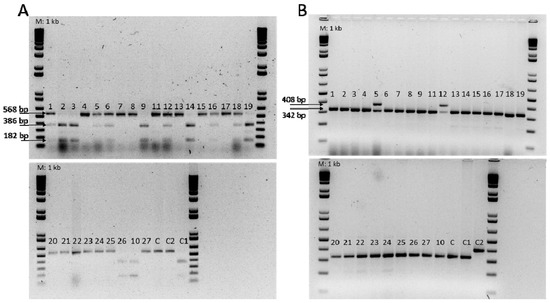
Figure 4.
(A) DNA analysis of Fom-1R CAPS marker. BspCNI digest generates 182 and 386 bp products for resistant genotype Fom-1/Fom-1; 182, 386, and 568 bp products for resistant genotype Fom-1/fom-1; and 568 bp products for susceptible genotype fom-1/fom-1. ‘Charentais T’ and ‘Charentais Fom-2’ (C and C2, respectively) were used as susceptible controls. ‘Charentais Fom-1’ (C1) was used as resistant control for the Fom-1 gene. M: 1 kb Plus DNA ladder (Invitrogen). (B) PCR amplification of the Fom-2 alleles, in melon genotypes, with SCAR primers (Fom2-R408; Fom2-S342). ‘Charentais T’ and ‘Charentais Fom-1’ (C and C1, respectively) were used as susceptible controls and ‘Charentais Fom-2’ (C2) was used as resistant control, for the Fom-2 gene. M: 1 kb Plus DNA ladder (Invitrogen).
Using the codominant Fom-2 SCAR marker, only a single band of 408 bp, corresponding to resistant Fom-2 allele, was amplified in the homozygous resistant genotype ‘Charentais Fom-2’, used as resistant control (Figure 4B). For the accessions TUN-5 and TUN-12, resistant to races 0 and 1 by phenotypic screening, this marker produced two bands of 408 and 342 bp signaling the presence of the Fom-2 resistance gene in the heterozygous state (Figure 4B). All the accessions susceptible to race 1 carried the susceptible alleles of the Fom-2 gene (Table 3).
3.2. Phenotypic Evaluations of the Resistance to Fom Pathotypes 1.2y and 1.2w
The majority of accessions showed clear Fusarium wilt symptoms with severe yellowing and wilting. Typical symptoms of Fusarium wilt were also observed in all the inoculated seedlings of the susceptible control lines, whereas all the inoculated seedlings of the resistant control ‘Dinero’ showed only mild or no symptoms (Supplementary Material). Pathotype 1.2y induced leaf yellowing before plant death, and seedlings inoculated with 1.2w wilted and died without previous yellowing symptoms.
In all the experiments, the AUDPC values for the susceptible controls, ‘Charentais-Fom-1’ and ‘Charentais-Fom-2’, were significantly higher than those for most of the tested accessions, while the AUDPC for ‘Dinero’, resistant control, was low (Table 4).
Table 4.
Mean values for the area under the disease progress curve (AUDPC) ± SD observed for Tunisian melon accessions inoculated with both pathotypes, 1.2y and 1.2w.
Large variability of the AUDPC values was observed among accessions for both pathotypes; thus, it was possible to detect differences in the level of resistance (Table 4). Two accessions, TUN-5 and TUN-26, showed a high level of resistance to both pathotypes (1.2y and 1.2w). They appeared to be highly resistant, although they displayed slightly lower AUDPC values than the resistant control ‘Dinero’ (Figure 5). To confirm the resistance found in those accessions, an additional phenotypic evaluation was performed, including all melon accessions and controls. The results obtained confirmed a high level of resistance of Tun-5 and Tun-26 to Fom 1.2y and 1.2w (Table 4).

Figure 5.
Resistant healthy accessions, Tun-5 and Tun-26, in comparison to the susceptible control (Charentais Fom-1) and resistant control (Dinero).
4. Discussion
One of the most convenient and environmentally safe options to control melon Fusarium wilt disease is the use of resistant varieties. Indeed, new melon lines containing resistance are being developed in backcross programs [38]. Several accessions carrying resistance to races 0, 1, and 2 of Fom have been described; most of the resistances are race-specific and controlled by Fom-1 and Fom-2 genes. To date, Fom-1 and Fom-2 genes have been extensively used in melon breeding programs; they have been introduced into the majority of the modern melon cultivars belonging mostly to Galia and Cantaloupe types [12]. However, due to the ability of adaptation and the evolution of Fusarium populations that could make specific resistance genes ineffective, the availability of resistance sources with different genetic bases is desirable. So, there is a need to screen new germplasm that could lead to the finding of new genes that could reinforce the existing resistance. On the other side, and although the introgression of these known genes is not complicated, the elimination of the nondesirable characteristics introgressed with the resistance makes breeding programs leading to obtaining varieties or hybrids of commercial value enormously difficult. Tunisian accessions are well adapted to the Mediterranean environment, and their fruits are very close to the fruits demanded by the Mediterranean, even European, markets, so the use of these accessions to obtain commercial hybrids or elite cultivars is promising; this is more strongly true in the introduction of resistance to race 1.2, only described in a few Eastern accessions [15,17].
In this study, Tunisian melon accessions were evaluated against Fom races 0, 1, 2, and 1.2 using artificial inoculation; subsequently, they were genotyped by using functional markers for the dominant and specific Fom-1 and Fom-2 genes [24,25] controlling resistance to races 0, 1, and 2. These markers allowed the identification of the resistance gene Fom-1 in 13 accessions, which were resistant to races 0 and 2. In seven of them, the gene Fom-1 was found to be in the homozygous state (TUN-2, TUN-3, TUN-9, TUN-10, TUN-14, TUN-19, and TUN-26), and six accessions were in the heterozygous state for this gene (TUN-1, TUN-5, TUN-6, TUN-12, TUN-16, and TUN-18). The accessions TUN-5 and TUN-12 also carried the gene Fom-2 in the heterozygous state, validating the resistance to races 0 and 1 detected by artificial inoculation. The utility of these allele-specific markers used was also confirmed, allowing the efficient identification of the resistant accessions. Accessions TUN-8 and TUN-25 showed resistance to race 0, but this resistance could not be explained by the resistance alleles of the Fom genes. This discrepancy could be because different levels of resistance are expressed under varying inoculation conditions [16] or because the genetic control of the resistance could be different and a new resistance gene could be found. Further artificial inoculation will be performed to confirm this resistance, and in the case of persistent resistance, additional genetic studies will be needed to determine the nature of such resistance. The same happened for the accession TUN-7, where resistance to race 1 could not be attributed to the presence of the resistance allele of Fom-2.
Resistance to the two pathotypes 1.2y and 1.2w was identified in two accessions (TUN-5 and TUN-26). They appeared to be highly resistant, despite displaying slightly lower resistance levels than the resistant control ‘Dinero’. However, it must be taken into account that the number of the known accessions resistant to this race is very low, and almost all of them belong to the makuwa or conomon groups, far away from the melon groups growing in the Mediterranean basin. So, the identified accessions TUN-5 and TUN-26 should be considered as very promising resistance sources to be used facing the losses caused by Fom 1.2. In both cases, further characterization of the resistance and selection are needed before the resistance can be incorporated into melon breeding programs. Indeed, up to now, resistance to race 1.2 is reported to be complex, controlled by multiple recessive genes with epistatic effects, which make its management difficult [13]. Besides, Herman and Perl-Treves [16] reported that resistance to this race is controlled by two recessive genes in the breeding line BIZ, and unfortunately, the molecular markers linked to those genes were not validated. Partial resistance to race 1.2 was also shown not to be race-specific in accessions belonging to makuwa and conomon groups [13,15]. The resistance herein identified against race 1.2 in TUN-5 and TUN-26 seems not to be race-specific since resistance was not only to race 1.2 but also to races 0, 1, and 2.
With the results obtained, TUN-5 (Fakous) is confirmed as an accession of high interest; it belongs to the flexuosus group, showing resistance to Fom 0, 1, and 2 controlled by the genes Fom-1 and Fom-2, and is highly resistant to Fom 1.2, although the genetic control of this resistance is still unknown. Fruits of the flexuosus group are in high demand for use in salads, not only in Tunisia but also in other Mediterranean countries. The fruit is long, with light to dark green skin and white to creamflesh, without sugar or aroma, and eaten raw before ripeness. This is, to the best of our knowledge, the first report of genetic resistance to this fungus in the flexuosus horticultural group. After its self-pollination and selection to obtain Fom-1 and Fom-2 in homozygosis, TUN-5 could be recommended to be used directly by farmers or to be exploited as a resistance donor in melon breeding programs to develop Fom-resistant cultivars of this type. Crosses between this accession and a Fom-susceptible Tunisian melon have been programmed to better understand the genetics of the resistance to race 1.2.
TUN-26 (Stambouli) is a highly productive Tunisian cultivar belonging to the inodorus group; this accession is resistant to Fom 0 and 2 (Fom-1 gene in homozygosis) and Fom 1.2 (pathotypes w and y). However, its fruit has poor organoleptic quality [29], so its major significance is the resistance shown against Fom race 1.2 that must be studied. The accession could be used either as a resistant melon rootstock or as a source of resistance to introgress the resistance into elite melon lines.
Studies will also continue with these two accessions (TUN-5 and TUN-26) to elucidate their genetic control and to provide useful molecular markers linked to Fom 1.2 resistance. Strategies such as association mapping and QTL mapping represent an opportunity to elucidate the genes or regions of the genome involved in resistance to Fom 1.2
Another remarkable accession is TUN-12 (Horchay), whose resistance to Fom 0, 1, and 2 is also conferred by Fom-1 and Fom-2; it belongs to the chate group. Its fruit is small-sized, round to oval, with ribs and dark green skin. The flesh is white to light orange, without aroma or sugar. It is harvested before maturity and eaten raw in salads. This type is cultivated in the Mediterranean basin and western Asia. Self-pollination and selection to obtain Fom-1 and Fom-2 in homozygosis are recommended before its use.
Multiple resistance to Fom races could be developed as a sustainable management strategy for melon culture in Tunisia and other Mediterranean areas where the races 0, 1, 2, and 1.2 of Fom have been identified [1,6]. Interestingly the resistance to race 1 observed in the accession TUN-7 seems not to be conferred by the Fom-2 gene. Further studies will be necessary to characterize the gene(s) involved in such resistance, as well as the genes involved in the resistance to race 0 found in TUN-8 and TUN-25. As mentioned above, the localization of resistance genes different from the mentioned genes could reinforce Fusarium wilt resistance in melon crops.
So far, most of the described resistances to Fom are race-specific. Many commercial varieties with specific resistance have been developed, but they become susceptible to infection after they are exploited. Therefore, the pyramiding of resistance genes to specific races in a single genotype, or the use of resistance that is not specific to race, should give more comprehensive and durable protection [12,13,39,40]. Consequently, the search for and utilization of new genes that confer resistance to Fusarium wilt is still one of the primary objectives in melon breeding. In this paper, new sources of resistance to Fom races have been found. These accessions belong to inodorus, cantalupensis, flexuosus, and reticulatus horticultural groups, which are of high interest since their agronomic traits and fruit characteristics are similar to commercial types demanded by markets.
Special attention should be given to the accession TUN-19 (Chamem), with resistance to Fom races 0 and 2 conferred by the Fom-1 gene. This accession, carrying the Vat gene [30], was previously reported to be resistant to Aphis gossypii (which causes direct damage to the plant by removing photoassimilates and indirect damage by transmitting pathogenic viruses). Therefore, the results obtained here emphasize the potential value of this melon accession as a donor of Fom disease and aphid resistance in melon breeding programs. This accession belongs to the Ananas type, which has yellow and netted skin and with good fruit quality [29], so TUN-19 is also an interesting and suitable accession as a source of both resistances to develop commercial melons of Ananas type, which are highly appreciated not only in Tunisia but also in other Mediterranean countries.
5. Conclusions
The current study demonstrated the importance of the Tunisian melon germplasm as a valuable genetic reservoir and the need to plan strategies for its conservation and utilization in breeding programs. Several Fusarium wilt-resistant accessions have been identified; some of them could contribute to reinforcing the known Fom 0, 1, and 2 resistances. Two accessions, with a high level of resistance to Fom race 1.2 (pathotypes 1.2y and 1.2w) have been also reported in this melon material. This local germplasm is of great potential and should be exploited since the morphological and organoleptic characteristics are very close to commercial types, highly appreciated not only in Tunisia but also in other Mediterranean countries. Specifically, TUN-5 and TUN-26 showed resistance to races 0, 1, and 2 and a high level of resistance to both pathotypes of race 1.2 and might be exploited either as a resistant melon rootstock or as a source for resistance in commercial flexuosus and inodorus lines, respectively. Further study is also needed to elucidate their genetic control and to provide useful molecular markers linked to Fom 1.2 resistance identified herein.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/horticulturae7080208/s1, Table S1: Scores of disease severity for races 0, 1, 2 and 1.2 for melon accessions.
Author Contributions
Conceptualization, H.C.-R., R.S.-B. and A.G.-C.; methodology, H.C.-R. and A.G.-C.; validation, H.C.-R. and A.G.-C.; formal analysis, H.C.-R. and A.G.-C.; writing—original draft preparation, H.C.-R.; writing—review and editing, H.C.-R., M.L.G.-G., V.G. and A.G.-C.; funding acquisition, A.G.-C., V.G. and M.L.G.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded, in part, by the Spanish Ministry of Economy and Competitivity cofunded with FEDER grant numbers AGL2017-85563-C2-2-R and PID2020-116055RB-C22 and partially financed by Aragon Government Grant for Research Group, A11-20R-PROVESOS. This work was supported in part by the Ministry of Higher Education and Scientific Research of Tunisia (MESRST) through the funding allocated to the Research Laboratory LR21AGR03.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors thank Mouna Gaddes-Chahed (CRRHAB) for inoculum preparation and Mohamed Ferjani (Sahline Station) for preparing plants. We are also grateful to R. González-Torres for his kind help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Luongo, L.; Ferrarini, A.; Haegi, A.; Vitale, S.; Polverari, A.; Belisario, A. Genetic diversity and pathogenicity of Fusarium oxysporum f.sp melonis races from different areas of Italy. J. Phytopathol. 2015, 163, 73–83. [Google Scholar] [CrossRef]
- González, V.; Armengol, J.; Garcés-Claver, A. First Report of Fusarium petroliphilum Causing Fruit Rot of Butternut Squash in Spain. Plant Dis. 2018, 102, 1662. [Google Scholar] [CrossRef]
- González, V.; Armijos, E.; Garcés-Claver, A. FungalEndophytes as Biocontrol Agents against the Main Soil-Borne Diseases of Melon and Watermelon in Spain. Agronomy 2020, 10, 820. [Google Scholar] [CrossRef]
- González, V.; García-Martínez, S.; Flores-León, A.; Ruiz, J.J.; Pico, M.B.; Garces-Claver, A. Neocosmospora keratoplastica, a relevant human fusarial pathogenis found to be associated with wilt and root rot of Muskmelon and Watermelon crops in Spain: Epidemiological and molecular evidences. Eur. J. Plant. Pathol. 2020, 156, 1189–1196. [Google Scholar] [CrossRef]
- González, V.; García-Martínez, S.; Ruiz, J.J.; Flores-León, A.; Picó, B.; Garcés-Claver, A. First Report of Neocosmospora falciformis Causing Wilt and Root Rot of Muskmelon in Spain. Plant Dis. 2020, 104, 1256. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; Sta-Baba, R.; Ayed, C.; Belgacem, S.; Boughalleb, N.; Cherif, M. Physiological races of Fusarium oxysporum f.sp. melonis in Tunisia. Phytoparasitica 2013, 41, 593–596. [Google Scholar] [CrossRef]
- Flores-León, A.; García-Martínez, S.; González, V.; Garcés-Claver, A.; Martí, R.; Julián, C.; Sifres, A.; Pérez-De-Castro, A.; Díez, M.J.; López, C.; et al. Grafting Snake Melon [Cucumis melo L. subsp. melo Var. flexuosus (L.) Naudin] in Organic Farming: Effects on Agronomic Performance; Resistance to Pathogens; Sugar, Acid, and VOC Profiles; and Consumer Acceptance. Front. Plant Sci. 2021, 12, 613845. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, A.; Salem, I.B.; M’hamdi, M.; Boughalleb, N. Relationship study among soils physico-chemical properties and Monosporascus cannonballus ascospores densities for cucurbit fields in Tunisia. Eur. J. Plant Pathol. 2019, 153, 65–78. [Google Scholar] [CrossRef]
- Risser, G.; Banihashemi, Z.; Davis, D.W. A proposed nomenclature of Fusarium oxysporum f.sp. melonis races and resistancegenes in Cucumis melo. Phytopathology 1976, 66, 1105–1106. [Google Scholar] [CrossRef]
- Tamietti, G.; Valentino, D. Soil solarization as an ecological method for the control of Fusarium wilt of melon in Italy. Crop Prot. 2006, 25, 389–397. [Google Scholar] [CrossRef]
- Oumouloud, A.; Arnedo-Andrés, M.S.; González-Torres, R.; Álvarez, J.M. Inheritance of resistance to Fusarium oxysporum f. sp. melonis races 0 and 2 in melon accession Tortuga. Euphytica 2010, 176, 183–189. [Google Scholar] [CrossRef]
- Oumouloud, A.; El-Otmani, M.; Chikh-Rouhou, H.; Garcés-Claver, A.; Torres, R.G.; Perltreves, R.; Alvarez, J.M. Breeding melon for resistance to Fusarium wilt: Recent developments. Euphytica 2013, 192, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Chikh-Rouhou, H.; González-Torres, R.; Oumouloud, A.; Alvarez, J.M. Inheritance of race 1.2 Fusarium wilt resistance in four melon cultivars. Euphytica 2011, 182, 177–186. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; González-Torres, R.; Álvarez, J.M. Characterization of the resistance to Fusarium oxysporum f.sp melonis race 1.2 in Cucumis melo ‘BG-5384’. In Proceedings of the Cucurbitaceae 2008, IXth EUCARPIA Meeting on Genetics and Breeding of Cucurbitaceae, Avignon, France, 21–24 May 2008; pp. 419–422. [Google Scholar]
- Perchepied, L.; Pitrat, M. Polygenic Inheritance of Partial Resistance to Fusarium oxysporum f. sp. melonis Race 1.2 in Melon. Phytopathology 2004, 94, 1331–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, R.; Perl-Treves, R. Characterization and Inheritance of a New Source of Resistance to Fusarium oxysporum f. sp. melonis Race 1.2 in Cucumis melo. Plant Dis. 2007, 91, 1180–1186. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; González-Torres, R.; Alvarez, J.M.; Oumouloud, A. Screening and Morphological Characterization of Melons for Resistance to Fusarium oxysporum f.sp. melonis Race 1.2. HortScience 2010, 45, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Chikh-Rouhou, H.; Alvarez, J.M.; González-Torres, R. Differential interaction between melon cultivars and race 1.2 of Fusarium oxysporum f.sp. melonis. Commun. Agric. Appl. Boil. Sci. 2007, 72, 825–829. [Google Scholar]
- Tezuka, T.; Waki, K.; Yashiro, K.; Kuzuya, M.; Ishikawa, T.; Takatsu, Y.; Miyagi, M. Construction of a linkage map and identification of DNA markers linked to Fom-1, a gene conferring resistance to Fusarium oxysporum f.sp. melonis race 2 in melon. Euphytica 2009, 168, 177–188. [Google Scholar] [CrossRef]
- Oumouloud, A.; Arnedo-Andres, M.S.; Gonzalez-Torres, R.; Alvarez, J.M. Development of molecular markers linked to the Fom-1 locus for resistance to Fusarium race 2 in melon. Euphytica 2008, 164, 347–356. [Google Scholar] [CrossRef]
- Brotman, Y.; Kovalski, I.; Dogimont, C.; Pitrat, M.; Portnoy, V.; Katzir, N.; Perl-Treves, R. Molecular markers linked to papaya ring spot virus resistance and Fusarium race 2 resistance in melon. Theor. Appl. Genet. 2004, 110, 337–345. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Thomas, C.E.; Dean, R.A. Genetic mapping of a Fusarium wilt resistance gene (Fom-2) in melon (Cucumis melo L.). Mol. Breed. 2000, 6, 379–389. [Google Scholar] [CrossRef]
- Wechter, W.P.; Whitehead, M.P.; Thomas, C.E.; Dean, R.A. Identification of a randomly amplified polymorphic DNA marker linked to the Fom-2 Fusarium wilt resistance gene in muskmelon MR-1. Phytopathology 1995, 85, 1245–1249. [Google Scholar] [CrossRef]
- Oumouloud, A.; El Otmani, M.; Alvarez, J.M.A. Molecular characterization of Fom-1 gene and development of functional markers for molecular breeding of resistance to Fusarium race 2 in melon. Euphytica 2015, 205, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Oumouloud, A.; Mokhtari, M.; Chikh-Rouhou, H.; Arnedo-Andrés, M.S.; González-Torres, R.; Álvarez, J.M. Characterization of the Fusarium wilt resistance Fom-2 gene in melon. Mol. Breed. 2012, 30, 325–334. [Google Scholar] [CrossRef]
- Andersen, J.R.; Lübberstedt, T. Functional markers in plants. Trends Plant Sci. 2003, 8, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Perchepied, L.; Dogimont, C.; Pitrat, M. Strain-specific and recessive QTLs involved in the control of partial resistance to Fusarium oxysporum f. sp. melonis race 1.2 in a recombinant inbred line population of melon. Theor. Appl. Genet. 2005, 111, 65–74. [Google Scholar] [CrossRef]
- Herman, R.; Zvirin, Z.; Kovalski, I. Characterization of Fusarium race 1.2 resistance in melon and mapping of a major QTL for this trait near a fruit netting locus. In Proceedings of the IXth Eucarpia Meeting on Genetics and Breeding of Cucurbitaceae 2008, Avignon, France, 21–24 May 2008; pp. 149–151. [Google Scholar]
- Chikh-Rouhou, H.; Tlili, I.; Ilahy, R.; R’Him, T.; Sta-Baba, R. Fruit quality assessment and characterization of melon genotypes. Int. J. Veg. Sci. 2021, 27, 3–19. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; Ben Belgacem, A.M.; Sta-Baba, R.; Tarchoun, N.; Gómez-Guillamón, M.L. New source of resistance to Aphis gossypii in Tunisian melon accessions using phenotypic and molecular marker approaches. Phytoparasitica 2019, 47, 405–413. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; Mezghani, N.; Mnasri, S.; Mezghani, N.; Garcés-Claver, A. Assessing the Genetic Diversity and Population Structure of a Tunisian Melon (Cucumis melo L.) Collection Using Phenotypic Traits and SSR Molecular Markers. Agronomy 2021, 11, 1121. [Google Scholar] [CrossRef]
- Pitrat, M. Melon Genetic Resources: Phenotypic Diversity and Horticultural Taxonomy. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer: Cham, Switzerland, 2016; Volume 20, pp. 25–60. [Google Scholar]
- Álvarez, J.M.; González-Torres, R.; Mallor, C.; Gómez-Guillamón, M.L. Potential sources of resistanse to Fusarium wilt and powdery mildew in melons. HortScience 2005, 40, 1657–1660. [Google Scholar] [CrossRef] [Green Version]
- Mas, P.; Risser, G. Caracterisation, Symptômes et Virulence de Diverses Races de Fusarium oxysporum Schl. f. sp. melonis Sn. et Hans. In Proceedings of the Actes du Premier Congres de l’Union Phytopathologique Mediterraneenne 1966, Bari-Naples, Italy, 23 September 1966. [Google Scholar]
- Rodríguez-Maza, M.J.; Garcés-Claver, A.; Park, S.-W.; Kang, B.-C.; Arnedo-Andrés, M.S. A versatile PCR marker for pungency in Capsicum spp. Mol. Breed. 2012, 30, 889–898. [Google Scholar] [CrossRef]
- Meighan, A.M.; Rabiei, B.; Khodaparast, S.A. Identification of Fusarium wilt resistance sources in melon (Cucumis melo L.) landraces of Iran using marker-assisted selection technique. Australas. Plant Pathol. 2020, 49, 413–423. [Google Scholar] [CrossRef]
- Chikh-Rouhou, H.; Garcés-Claver, A.; Sta-Baba, R.; González, V.; Daami-Remadi, M. Screening for resistance to race 1 of Fusarium oxysporum f.sp melonis in Tunisian melon cultivars using molecular markers. Comm. Agric. Appl. Biol. Sci. 2018, 83, 87–92. [Google Scholar]
- Palomares-Rius, F.; Garcés-Claver, A.; Picó, B.; Esteras, C.; Yuste-Lisbona, F.; Gómez-Guillamón, M.L. ‘Carmen’, a Yellow-Canary melon resistant to Podosphaera xanthii, Aphis gossypii and Cucurbit Yellow Stunting Disorder Virus. HortScience 2018, 53, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Oumouloud, A.; Torres, R.G.; Garcés-Claver, A.; Rouhou, H.C.; Álvarez, J. Differential response of Cucumis melo to Fusarium oxysporum f. sp. melonis race 1.2 isolates. Crop Prot. 2013, 44, 91–94. [Google Scholar] [CrossRef]
- Dhillon, N.P.S.; Monforte, A.J.; Pitrat, M.; Pandey, S.; Singh, P.K.; Reitsma, K.R.; Garcia-Mas, J.; Sharma, A.; McCreight, J.D. Melon landraces of India: Contributions and importance. In Plant Breeding Reviews; Janick, J., Ed.; Wiley: Hoboken, NJ, USA, 2012; Volume 35, pp. 85–150. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).