
The Growth of Leaf Lettuce and Bacterial Communities in a Closed Aquaponics System with Catfish

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fish Materials
2.2. Experimental Design and System
2.3. Measurements of Solution Quality
2.4. Growth Survey of Lettuce
2.5. Analysis of Bacterial Flora
3. Results
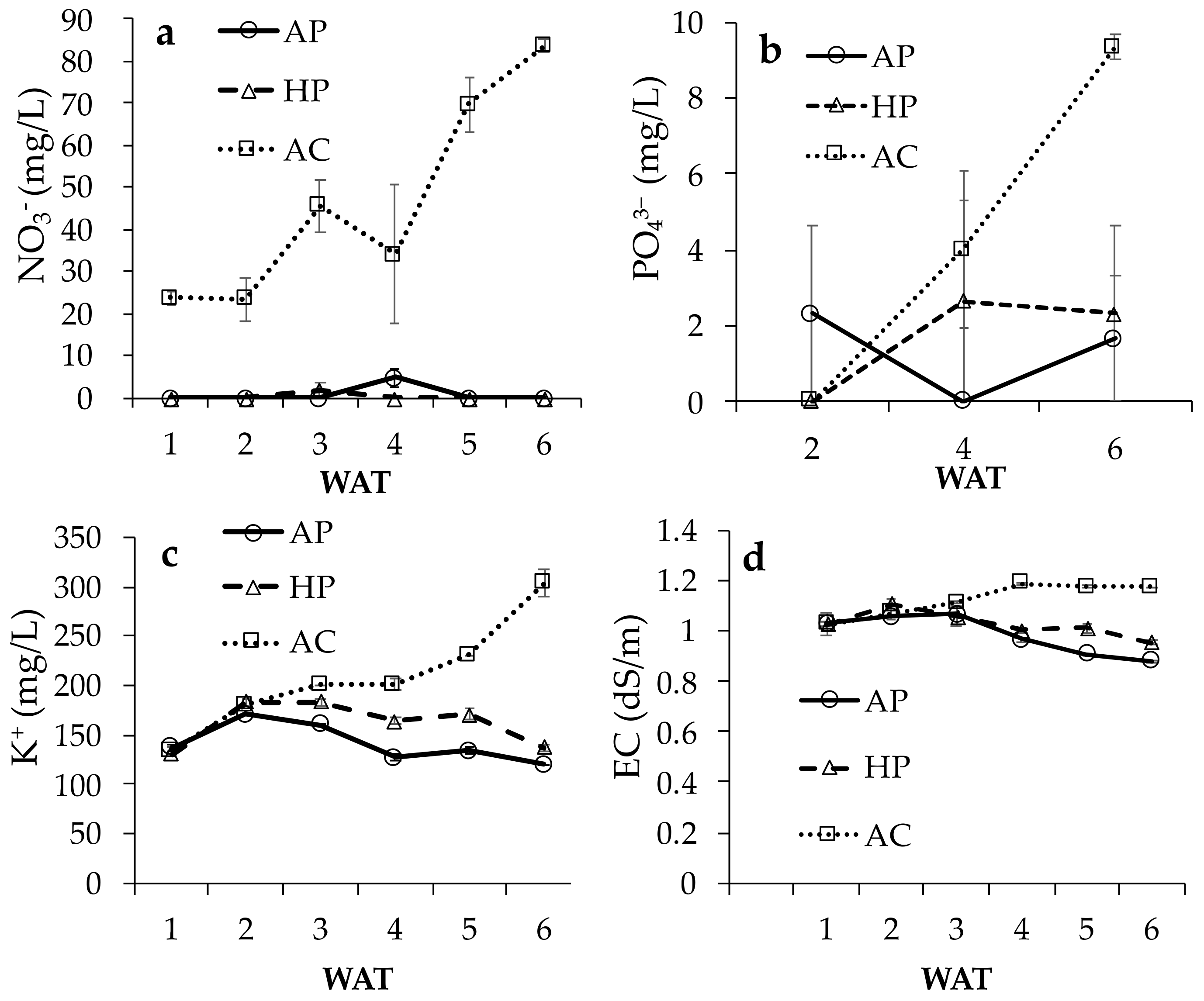
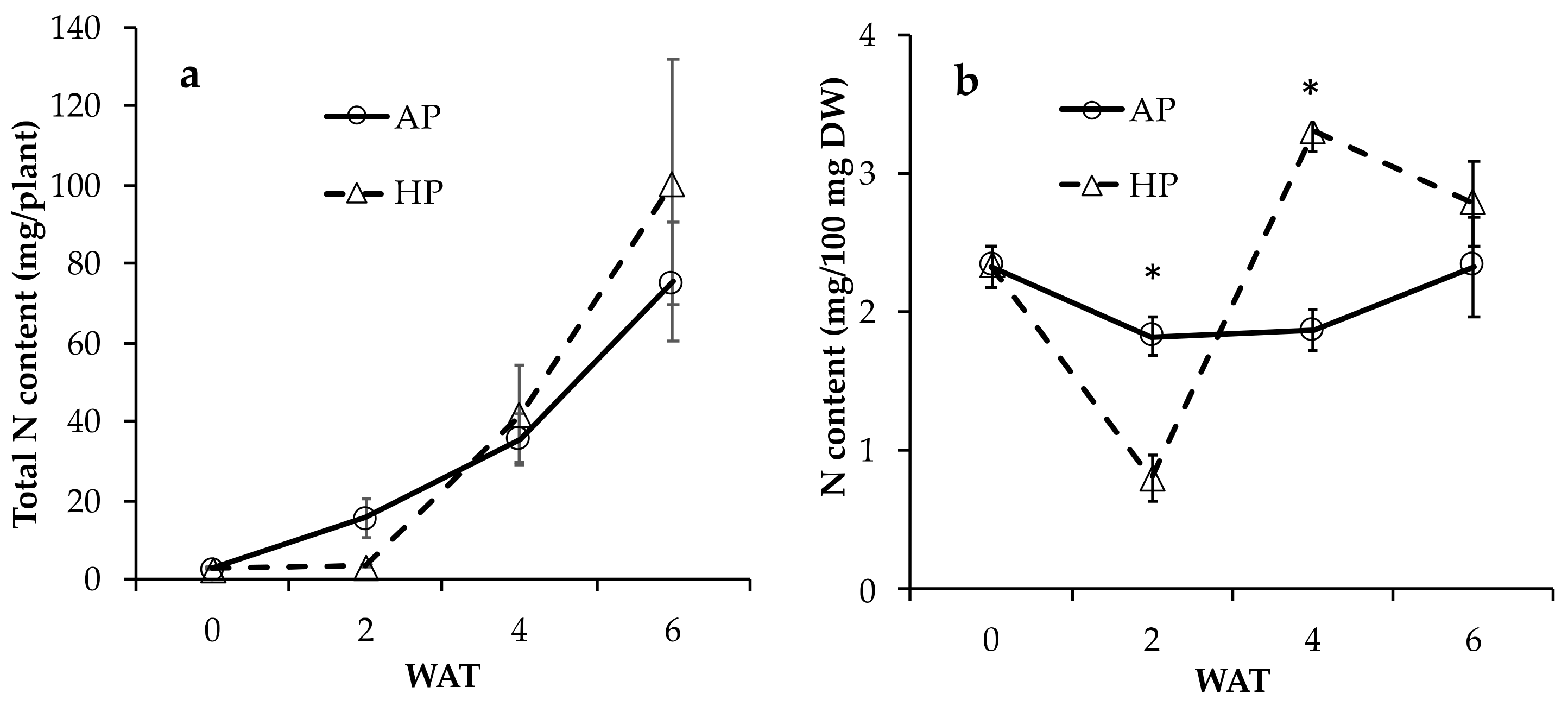
3.1. Nutrition of Solutions and Growth of Lettuce Plants
3.1.1. Nutrition and EC of Solutions
3.1.2. Growth of Lettuce Plants in AC and HP
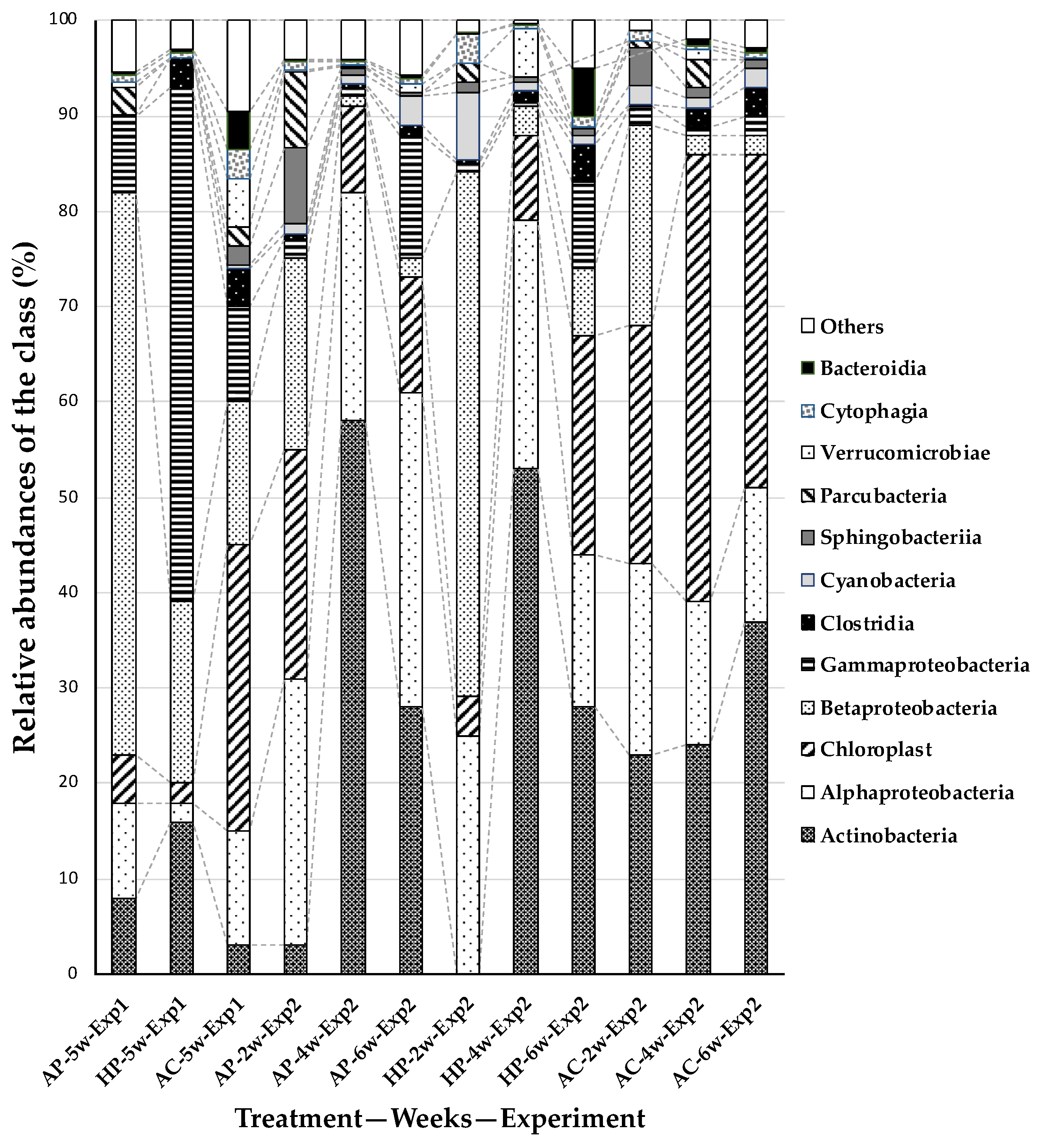
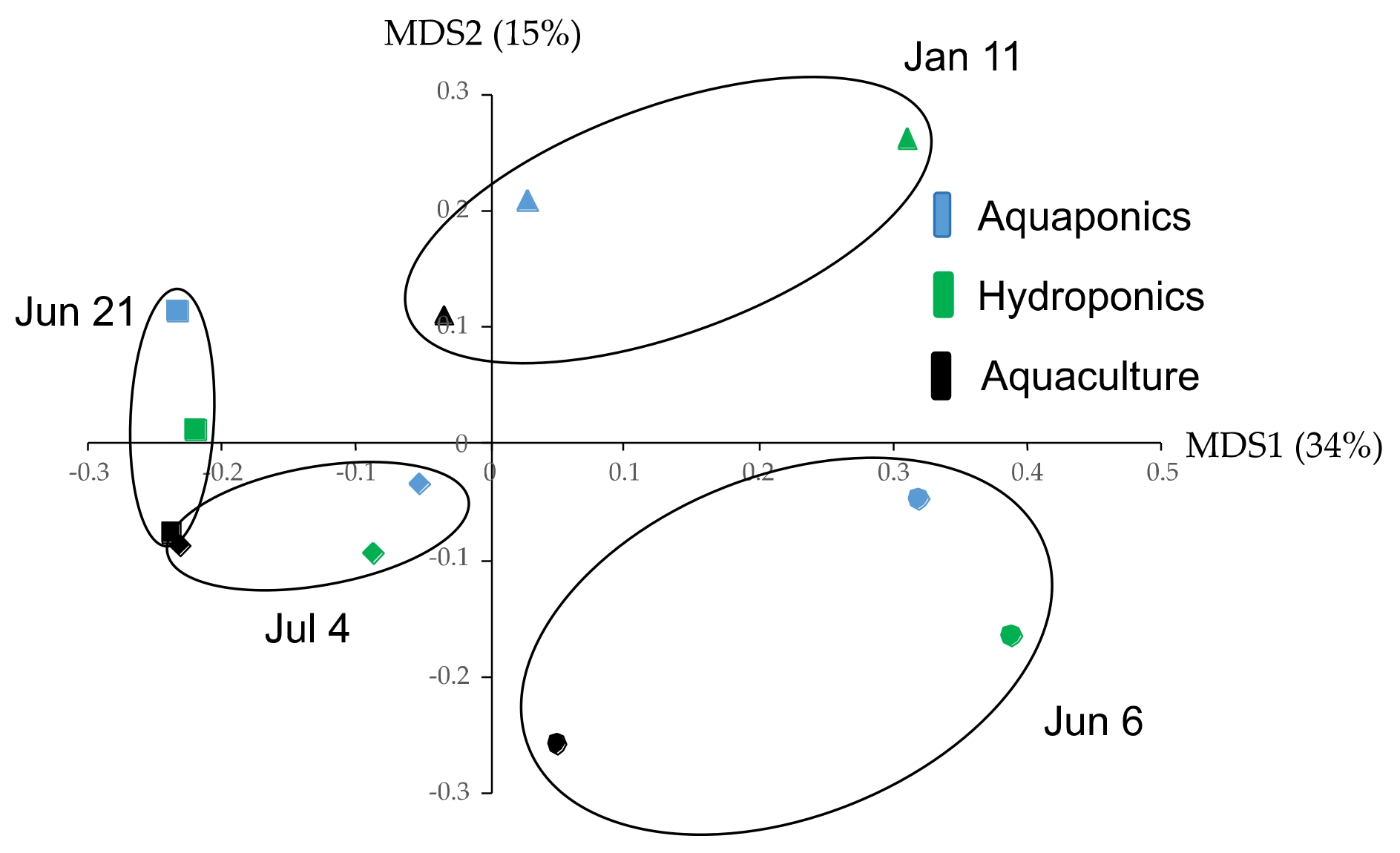
3.2. Bacterial Flora in Solution
3.2.1. Ratio of Bacteria Class
3.2.2. Specific Bacteria OTUs and Existence of Nitrifying Bacteria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yep, B.; Zheng, Y. Aquaponic trends and challenges—A review. J. Clean. Prod. 2019, 228, 1586–1599. [Google Scholar] [CrossRef]
- Frincu, M.; Dumitrache, C. Study regarding nitrification in experimental aquaponics system. J. Young Sci. 2016, 4, 27–32. [Google Scholar]
- Tyson, R.V.; Treadwell, D.D.; Simonne, E.H. Opportunities and Challenges to Sustainability in Aquaponic Systems. HortTechnology 2011, 21, 6–13. [Google Scholar] [CrossRef]
- Endut, A.; Lananan, F.; Jusoh, A.; Nik, W.N.W.; Ali, N. Aquaponics recirculation system: A sustainable food source for the future water conserves and resources. Malays. J. Appl. Sci. 2016, 1, 1–12. [Google Scholar]
- Hargreaves, J.A. Nitrogen biogeochemistry of aquaculture ponds. Aquaculture 1998, 166, 181–212. [Google Scholar] [CrossRef]
- Yang, T.; Kim, H.-J. Characterizing Nutrient Composition and Concentration in Tomato-, Basil-, and Lettuce-Based Aquaponic and Hydroponic Systems. Water 2020, 12, 1259. [Google Scholar] [CrossRef]
- Sanchez, F.A.; Vivian-Rogers, V.R.; Urakawa, H. Tilapia recirculating aquaculture systems as a source of plant growth promoting bacteria. Aquac. Res. 2019, 50, 2054–2065. [Google Scholar] [CrossRef]
- Rakocy, J.E.; Masser, M.P.; Losordo, T.M. Recirculating Aquaculture Tank Production Systems: Aquaponics-Integrating Fish and Plant Culture; SRAC: Stoneville, MS, USA, 2006. [Google Scholar]
- Lennard, W.A.; Leonard, B.V. A comparison of three different hydroponic sub-systems (gravel bed, floating and nutrient film technique) in an aquaponic test system. Aquac. Int. 2006, 14, 539–550. [Google Scholar] [CrossRef]
- Sace, C.F.; Fitzsimmons, K.M. Vegetable production in a recirculating aquaponic system using Nile tilapia (Oreochromis niloticus) with and without freshwater prawn (Macrobrachium rosenbergii). Acad. J. Agric. Res. 2013, 1, 236–250. [Google Scholar]
- Danaher, J.J.; Shultz, R.C.; Rakocy, J.E.; Bailey, D.S. Alternative Solids Removal for Warm Water Recirculating Raft Aquaponic Systems. J. World Aquac. Soc. 2013, 44, 374–383. [Google Scholar] [CrossRef]
- Seawright, D.E.; Stickney, R.R.; Walker, R.B. Nutrient dynamics in integrated aquaculture–hydroponics systems. Aquaculture 1998, 160, 215–237. [Google Scholar] [CrossRef]
- Liang, J.-Y.; Chien, Y.-H. Effects of feeding frequency and photoperiod on water quality and crop production in a tilapia–water spinach raft aquaponics system. Int. Biodeterior. Biodegrad. 2013, 85, 693–700. [Google Scholar] [CrossRef]
- Bartelme, R.P.; Oyserman, B.O.; Blom, J.E.; Sepulveda-Villet, O.J.; Newton, R.J. Stripping Away the Soil: Plant Growth Promoting Microbiology Opportunities in Aquaponics. Front. Microbiol. 2018, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmautz, Z.; Graber, A.; Jaenicke, S.; Goesmann, A.; Junge, R.; Smits, T.H. Microbial diversity in different compartments of an aquaponics system. Arch. Microbiol. 2017, 199, 613–620. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J. Beneficial bacteria and fungi in hydroponic systems: Types and characteristics of hydroponic food production methods. Sci. Hortic. 2015, 195, 206–215. [Google Scholar] [CrossRef]
- Khalil, S. Growth performance, nutrients and microbial dynamic in aquaponics systems as affected by water temperature. Eur. J. Hortic. Sci. 2018, 83, 388–394. [Google Scholar] [CrossRef]
- Hüpeden, J.; Wegen, S.; Off, S.; Lucker, S.; Bedarf, Y.; Daims, H.; Kuhn, C.; Spieck, E. Relative Abundance of Nitrotoga spp. in a Biofilter of a Cold-Freshwater Aquaculture Plant Appears To Be Stimulated by Slightly Acidic pH. Appl. Environ. Microbiol. 2016, 82, 1838–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelme, R.P.; Smith, M.C.; Sepulveda-Villet, O.J.; Newton, R.J. Component microenvironments and system biogeography structure microorganism distributions in recirculating aquaculture and aquaponic systems. mSphere 2019, 4, e00143-19. [Google Scholar] [CrossRef] [Green Version]
- Eck, M.; Sare, A.; Massart, S.; Schmautz, Z.; Junge, R.; Smits, T.; Jijakli, M. Exploring Bacterial Communities in Aquaponic Systems. Water 2019, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Ebihara, N.; Washio, S.; Sugita, H. Nitrite-oxidizing bacteria, Nitrospira, distribution in the outer layer of the biofilm from filter materials of a recirculating water system for the goldfish Carassius auratus. Aquaculture 2007, 264, 297–308. [Google Scholar] [CrossRef]
- Rurangwa, E.; Verdegem, M.C.J. Microorganisms in recirculating aquaculture systems and their management. Rev. Aquac. 2015, 7, 117–130. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Wongkiew, S.; Park, M.R.; Chandran, K.; Khanal, S.K. Aquaponic systems for sustainable resource recovery: Linking nitrogen transformations to microbial communities. Environ. Sci. Technol. 2018, 52, 12728–12739. [Google Scholar] [CrossRef]
- Espinosa Moya, E.A.; Angel Sahagún, C.A.; Mendoza Carrillo, J.M.; Albertos Alpuche, P.J.; Álvarez-González, C.A.; Martínez-Yáñez, R. Herbaceous plants as part of biological filter for aquaponics system. Aquac. Res. 2016, 47, 1716–1726. [Google Scholar] [CrossRef]
- Goddek, S.; Körner, O. A fully integrated simulation model of multi-loop aquaponics: A case study for system sizing in different environments. Agric. Syst. 2019, 171, 143–154. [Google Scholar] [CrossRef]
- Gruda, N. Increasing Sustainability of Growing Media Constituents and Stand-Alone Substrates in Soilless Culture Systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Körner, O.; Bisbis, M.B.; Baganz, G.F.M.; Baganz, D.; Staaks, G.B.O.; Monsees, H.; Goddek, S.; Keesman, K.J. Environmental impact assessment of local decoupled multi-loop aquaponics in an urban context. J. Clean. Prod. 2021, 313, 127735. [Google Scholar] [CrossRef]
- Wongkiew, S.; Hu, Z.; Chandran, K.; Lee, J.W.; Khanal, S.K. Nitrogen transformations in aquaponic systems: A review. Aquac. Eng. 2017, 76, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Clavijo-Herrera, J.; van Santen, E.; Gómez, C. Growth, Water-Use Efficiency, Stomatal Conductance, and Nitrogen Uptake of Two Lettuce Cultivars Grown under Different Percentages of Blue and Red Light. Horticulturae 2018, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Kasozi, N.; Abraham, B.; Kaiser, H.; Wilhelmi, B. The complex microbiome in aquaponics: Significance of the bacterial ecosystem. Ann. Microbiol. 2021, 71, 1–13. [Google Scholar] [CrossRef]
- Urakawa, H.; Dettmar, D.L.; Thomas, S. The uniqueness and biogeochemical cycling of plant root microbial communities in a floating treatment wetland. Ecol. Eng. 2017, 108, 573–580. [Google Scholar] [CrossRef]
- Tsuchiya, C.; Sakata, T.; Sugita, H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 2008, 46, 43–48. [Google Scholar] [CrossRef]
- Suarez, C.; Ratering, S.; Schafer, J.; Schnell, S. Ancylobacter pratisalsi sp. nov. with plant growth promotion abilities from the rhizosphere of Plantago winteri Wirtg. Int. J. Syst. Evol. Microbiol. 2017, 67, 4500–4506. [Google Scholar] [CrossRef] [PubMed]
- Day, J.A.; Otwell, A.E.; Diener, C.; Tams, K.E.; Bebout, B.; Detweiler, A.M.; Lee, M.D.; Scott, M.T.; Ta, W.; Ha, M.; et al. Negative plant-microbiome feedback limits productivity in aquaponics. bioRxiv 2019, 709162. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Ishikawa, D.; Saeki, G.; Aoi, Y.; Tsuneda, S. Microbial population dynamics and community structure during the formation of nitrifying granules to treat ammonia-rich inorganic wastewater. Microbes Environ. 2010, 25, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Saijai, S.; Ando, A.; Inukai, R.; Shinohara, M.; Ogawa, J. Analysis of microbial community and nitrogen transition with enriched nitrifying soil microbes for organic hydroponics. Biosci. Biotechnol. Biochem. 2016, 80, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gichana, Z.M.; Liti, D.; Waidbacher, H.; Zollitsch, W.; Drexler, S.; Waikibia, J. Waste management in recirculating aquaculture system through bacteria dissimilation and plant assimilation. Aquac. Int. 2018, 26, 1541–1572. [Google Scholar] [CrossRef]
- Chen, P.; Zhu, G.; Kim, H.-J.; Brown, P.B.; Huang, J.-Y. Comparative life cycle assessment of aquaponics and hydroponics in the Midwestern United States. J. Clean. Prod. 2020, 275, 122888. [Google Scholar] [CrossRef]
- Arsin, N.E.; Estim, A.; Mustafa, S. Behavior and response of Japanese catfish (Silurus asotus) in captivity provided with an artificial microhabitat mosaic. Aquat. Res. 2018, 1, 136–139. [Google Scholar] [CrossRef]
- Higuchi, K.; Kazeto, Y.; Ozaki, Y.; Yamaguchi, T.; Shimada, Y.; Ina, Y.; Soma, S.; Sakakura, Y.; Goto, R.; Matsubara, T.; et al. Targeted mutagenesis of the ryanodine receptor by Platinum TALENs causes slow swimming behaviour in Pacific bluefin tuna (Thunnus orientalis). Sci. Rep. 2019, 9, 13871. [Google Scholar] [CrossRef] [Green Version]
- Hochmuth, G.J. Fertilizer management for greenhouse vegetables. In Florida Greenhouse Vegetable Production Handbook; Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2001; Volume 3. [Google Scholar]
- Zou, Y.; Hu, Z.; Zhang, J.; Xie, H.; Guimbaud, C.; Fang, Y. Effects of pH on nitrogen transformations in media-based aquaponics. Bioresour. Technol. 2016, 210, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Suhl, J.; Dannehl, D.; Kloas, W.; Baganz, D.; Jobs, S.; Scheibe, G.; Schmidt, U. Advanced aquaponics: Evaluation of intensive tomato production in aquaponics vs. conventional hydroponics. Agric. Water Manag. 2016, 178, 335–344. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treat. | Number of Leaves | Maximum Length of Leaf (cm) | Fresh Weight of Shoots (g) | Fresh Weight of Roots (g) | SPAD Value |
|---|---|---|---|---|---|
| 0 weeks after transplanting (WAT) | |||||
| 8.0 ± 0.5 1 | 6.5 ± 0.4 | 0.46 ± 0.06 | 0.025 ± 0.009 | 22.4 ± 2.1 | |
| 2 WAT | |||||
| AP | 10.7 ± 0.3 | 12.1 ± 0.3 | 9.9 ± 1.7 | 2.1 ± 0.4 | 27.6 ± 0.3 |
| HP | 9.0 ± 0.6 | 8.4 ± 0.7 | 5.2 ± 1.4 | 3.3 ± 1.0 | 23.3 ± 1.3 |
| T test | NS 2 | ** | NS | NS | * |
| 4 WAT | |||||
| AP | 15.0 ± 0.0 | 26.3 ± 0.4 | 79.4 ± 2.6 | 14.8 ± 2.0 | 30.1 ± 1.1 |
| HP | 13.3 ± 0.3 | 14.9 ± 0.8 | 36.6 ± 1.2 | 7.9 ± 1.0 | 26.4 ± 1.0 |
| T test | ** | *** | *** | * | NS |
| 6 WAT | |||||
| AP | 16.2 ± 1.7 | 29.9 ± 0.9 | 88.9 ± 17.0 | 11.6 ± 2.9 | 28.9 ± 1.0 |
| HP | 15.8 ± 1.2 | 21.5 ± 1.5 | 68.0 ± 9.8 | 12.9 ± 2.4 | 27.1 ± 1.2 |
| T test | NS | ** | NS | NS | NS |
| Treat. | Number of Leaves | Maximum Length of Leaf (cm) | Fresh Weight of Shoots (g) | Fresh Weight of Roots (g) | SPAD Value |
|---|---|---|---|---|---|
| 0 weeks after transplanting (WAT) | |||||
| 4.5 ± 0.2 1 | 7.5 ± 0.6 | 1.0 ± 0.2 | 0.2 ± 0.0 | 25.0 ± 1.8 | |
| 2 WAT | |||||
| AP | 9.8 ± 1.5 | 10.7 ± 0.6 | 10.5 ± 2.5 | 6.6 ± 1.6 | 23.9 ± 0.6 |
| HP | 6.0 ± 0.4 | 8.4 ± 0.5 | 2.7 ± 0.3 | 2.6 ± 0.2 | 18.1 ± 0.9 |
| T test | NS 2 | * | * | * | ** |
| 4 WAT | |||||
| AP | 14.5 ± 0.9 | 15.2 ± 1.1 | 29.7 ± 4.0 | 19.3 ± 3.5 | 24.8 ± 1.4 |
| HP | 11.3 ± 1.5 | 11.5 ± 2.0 | 23.2 ± 7.5 | 12.7 ± 4.4 | 24.4 ± 0.8 |
| T test | NS | NS | NS | NS | NS |
| 6 WAT | |||||
| AP | 21.0 ± 2.4 | 16.0 ± 1.5 | 56.4 ± 19.8 | 28.6 ± 6.1 | 26.8 ± 2.1 |
| HP | 18.3 ± 3.1 | 20.8 ± 1.4 | 72.1 ± 16.3 | 26.8 ± 6.4 | 28.0 ± 0.5 |
| T test | NS | NS | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamane, K.; Kimura, Y.; Takahashi, K.; Maeda, I.; Iigo, M.; Ikeguchi, A.; Kim, H.-J. The Growth of Leaf Lettuce and Bacterial Communities in a Closed Aquaponics System with Catfish. Horticulturae 2021, 7, 222. https://doi.org/10.3390/horticulturae7080222
Yamane K, Kimura Y, Takahashi K, Maeda I, Iigo M, Ikeguchi A, Kim H-J. The Growth of Leaf Lettuce and Bacterial Communities in a Closed Aquaponics System with Catfish. Horticulturae. 2021; 7(8):222. https://doi.org/10.3390/horticulturae7080222
Chicago/Turabian StyleYamane, Kenji, Yuuki Kimura, Keita Takahashi, Isamu Maeda, Masayuki Iigo, Atsuo Ikeguchi, and Hye-Ji Kim. 2021. "The Growth of Leaf Lettuce and Bacterial Communities in a Closed Aquaponics System with Catfish" Horticulturae 7, no. 8: 222. https://doi.org/10.3390/horticulturae7080222
APA StyleYamane, K., Kimura, Y., Takahashi, K., Maeda, I., Iigo, M., Ikeguchi, A., & Kim, H.-J. (2021). The Growth of Leaf Lettuce and Bacterial Communities in a Closed Aquaponics System with Catfish. Horticulturae, 7(8), 222. https://doi.org/10.3390/horticulturae7080222