
Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants

 ,
,  ,
,  , and
, and
Abstract
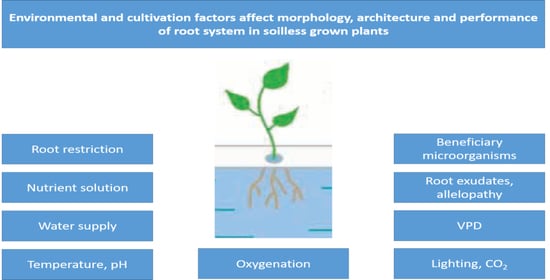
:
1. Introduction
2. Root Restriction
3. Nutrient Solution

{kind=link}
| Plant Response | Crop | SCS | Additional Information | Reference |
|---|---|---|---|---|
| No increase in plant yield by increasing N fertilization rates | Lettuce | Washed sand; 2.5 L (no confinement, the control); 1.0 L (moderate) and 0.4 L (severe root restriction) | Total nitrogen concentrations in mM L1, 5.55, 8.05, 10.55, 13.05 and 15.55. | [50] |
| Spinach | Styrofoam trays floated into 80 cm × 44 cm × 19 cm (52 L) plastic basins | “Full dose” nutrient solution (mg L−1: N 150, P 50, K 150, Ca 150, Mg 50, Fe 5.0, Mn 0.50, Zn 0.05, B 0.50, Cu 0.03, Mo 0.02), “half dose” (with macro elements reduced by 50%) | [51] | |
| Baby leaf lettuce | Styrofoam trays floated into 135 cm × 125 cm × 20 cm a flotation bed | Nutrient solutions with 12 and 4 mM L−1 N | [52] | |
| Primary root growth inhibition, increase in lateral roots and root hairs | Various crops | Various production system | Limited P supply | [39,41,46] |
| Increase in vertical, deep roots | Various crops | Various production systems | Limited N supply | [40,43,47] |
| Increased root dry weight, specific root length, root tissue density, and root length density due to increased irrigation intervals | Tomato, Zinnia | 450 mL plastic pots containing either Metromix 360 (MM360) or Ball Professional Growing Mix (BPGM) | 24-h, 48-h, and 96-h irrigation intervals | [53] |
| Chili pepper | 31 cm × 15 cm × 60 cm container filled with sandy-loamy soil | 1-, 3- and 5-day irrigation intervals | [54] |
4. Water Supply and Irrigation
5. Rootzone Temperature
| Plant Response | Crop | Production System | Additional Information | Reference |
|---|---|---|---|---|
| Increased root length | Cucumber | Plastic pots filled with sand | 12 °C vs. 20 °C | [87] |
| Garden pea | Foam trays filled with peat | 12 °C vs. 20 °C | [88] | |
| Increased root branching | Oilseed rape | Petri dishes filled with agar | 10, 15 and 20 °C | [83] |
| Several species | Transparent cylinders, filled with a growth medium made from half-strength Hoagland solution and 0.2% Phytagel | 18–34 °C | [88] | |
| Increased root density | Oilseed rape | Petri dishes filled with agar | 10, 15 and 20 °C | [83] |
| Garden pea | Foam trays filled with peat | 12 °C vs. 20 °C | [89] | |
| Reduced yield | Lettuce | DFT hydroponic system with | 25 °C and 30 °C vs. 10 °C | [90] |
| Tomato | Rockwool, cubs and slabs | 16–27 °C vs. 10 °C | [91] | |
| Tomato | NFT hydroponic system | 20.3 vs. 16.6 °C and14.2 vs. 5.8 °C | [92] | |
| Baby leaves of lettuce and rocket | Floating system | 30 vs. 21.9 °C | [93] |
6. Oxygenation
| Plant Response | Crop | Production System | Additional Information | Reference |
|---|---|---|---|---|
| Alterations in formation, elongation, and growth angle of roots. | Various crops | Various systems | O2 deficiency | [109] |
| Adventitious root formation, aerenchyma, and radial oxygen-loss barrier development | Cucumber | Floating system | O2 deficiency | [20] |
| Tomato | Flow-through hydroponic culture system (FTS) | O2 deficiency | [21] | |
| Various crops | Various systems | O2 deficiency | [113] | |
| Increased yield | Melon | Rockwool, cubs, and slabs | Oxygen enrichment | [116] |
| Melon | Rockwool, cubs, and slabs | Oxygen enrichment | [117] | |
| Lettuce | Nutrient solution | Oxygen enrichment | [118] |
7. Water Pressure Deficit
8. Lighting
9. CO2
10. Rootzone pH
11. Root Exudates and Allelopathy
12. Root–Microbial Relationships
13. Conclusions, Trends and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Nibau, C.; Gibbs, D.; Coates, J.C. Branching out in new directions: The control of root architecture by lateral root formation. New Phytol. 2008, 179, 595–614. [Google Scholar] [CrossRef]
- Teixeira, J.S.; Tusscher, K.T. The Systems Biology of Lateral Root Formation: Connecting the Dots. Mol. Plant 2019, 12, 784–803. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Rasmussen, A.; Traini, R.; Voss, U.; Sturrock, C.; Mooney, S.J.; Wells, D.; Bennett, M.J. Branching Out in Roots: Uncovering Form, Function, and Regulation. Plant Physiol. 2014, 166, 538–550. [Google Scholar] [CrossRef] [Green Version]
- Lavenus, J.; Goh, T.; Roberts, I.; Guyomarc’H, S.; Lucas, M.; De Smet, I.; Fukaki, H.; Beeckman, T.; Bennett, M.; Laplaze, L. Lateral root development in Arabidopsis: Fifty shades of auxin. Trends Plant Sci. 2013, 18, 450–458. [Google Scholar] [CrossRef]
- Vives-Peris, V.; López-Climent, M.F.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Root Involvement in Plant Responses to Adverse Environmental Conditions. Agronomy 2020, 10, 942. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; Forde, B. Quantitative Analysis of Lateral Root Development: Pitfalls and How to Avoid Them. Plant Cell 2012, 24, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tüzel, Y.; Balliu, A. Advances in liquid -and solid-medium soilless culture systems. In Advances in Horticultural Soilless Culture; Gruda, N.S., Ed.; Burleigh Dodds Science: Cambridge, UK, 2021; ISBN 978 1 78676 292 4. Available online: www.bdspublishing.com (accessed on 10 July 2021). [CrossRef]
- Sabatino, L. Increasing Sustainability of Growing Media Constituents and Stand-Alone Substrates in Soilless Culture Systems—An Editorial. Agronomy 2020, 10, 1384. [Google Scholar] [CrossRef]
- Savvas, D.; Gruda, N. Application of soilless culture technologies in the modern greenhouse industry—A review. Eur. J. Hortic. Sci. 2018, 83, 280–293. [Google Scholar] [CrossRef]
- Gruda, N. Soilless Culture Systems and Growing Media in Horticulture: An Overview. In Advances in Horticultural Soilless Culture; Gruda, N.S., Ed.; Burleigh Dodds Science: Cambridge, UK, 2021; pp. 1–20. ISBN 978 1 78676 292 4. Available online: https://shop.bdspublishing.com/store/bds/detail/workgroup/3-190-89123 (accessed on 10 July 2021). [CrossRef]
- Raviv, M.; Lieth, J.H.; Bar-Tal, A.; Silber, A. Growing plants in soilless culture: Operational conclusions. In Soilless Culture; Elsevier BV: Alpharetta, GA, USA, 2008; pp. 545–571. [Google Scholar]
- Maclellan, L. Effect of Hydroculture Methods on Tomato Root Morphology and Anatomy. Honor. Coll. 2019, 567. Available online: https://digitalcommons.library.umaine.edu/honors/567 (accessed on 11 April 2021).
- Shi, K.; Hu, W.-H.; Dong, D.-K.; Zhou, Y.-H.; Yu, J.-Q. Low O2 supply is involved in the poor growth in root-restricted plants of tomato (Lycopersicon esculentum Mill.). Environ. Exp. Bot. 2007, 61, 181–189. [Google Scholar] [CrossRef]
- Kafkafi, U. Root Temperature, Concentration and the Ratio NO3− / NH4+ Effect on Plant Development. J. Pl. Nutr. 1990, 13, 1291–1306. [Google Scholar] [CrossRef]
- Jones, J.B. Growing Plants Hydroponically; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Bláha, L. Crop Science Institute Importance of Root-Shoot Ratio for Crops Production. Agron. Agric. Sci. 2019, 2, 1–7. [Google Scholar] [CrossRef]
- Zakaria, N.I.; Ismail, M.R.; Awang, Y.; Wahab, P.E.M.; Berahim, Z. Effect of Root Restriction on the Growth, Photosynthesis Rate, and Source and Sink Relationship of Chilli (Capsicum annuum L.) Grown in Soilless Culture. BioMed Res. Int. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, T.; Wheeler, R. Root restriction: A tool for improving volume utilization efficiency in bioregenerative life-support systems. Life Sci. Space Res. 2016, 9, 62–68. [Google Scholar] [CrossRef]
- Kharkina, T.G.; Ottosen, C.-O.; Rosenqvist, E. Effects of root restriction on the growth and physiology of cucumber plants. Physiol. Plant. 1999, 105, 434–441. [Google Scholar] [CrossRef]
- Peterson, T.A.; Reinsel, M.D.; Krizek, D.T. Tomato (Lycopersicon esculentum Mill., cv. ‘Better Bush’) Plant Response to Root Restriction. I. Alteration of plant morphology. J. Exp. Bot. 1991, 42, 1233–1240. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Effect of pot volume on the growth of sweetpotato cultivated in the new hydroponic system. Sustain. Agric. Res. 2018, 77, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Tuzel, I.; Tüzel, Y.; Gül, A.; Altunlu, H.; Eltez, R. Effect of different irrigation schedules, substrates and substrate volumes on fruit quality and yield of greenhouse tomato. Acta Hortic. 2001, 548, 285–292. [Google Scholar] [CrossRef]
- Saito, T.; Fukuda, N.; Iikubo, T.; Inai, S.; Fujii, T.; Konishi, C.; Ezura, H. Effects of Root-volume Restriction and Salinity on the Fruit Yield and Quality of Processing Tomato. J. Jpn. Soc. Hortic. Sci. 2008, 77, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Pires, R.C.D.M.; Furlani, P.R.; Ribeiro, R.V.; Junior, D.B.; Sakai, E.; Lourenção, A.L.; Neto, A.T. Irrigation frequency and substrate volume effects in the growth and yield of tomato plants under greenhouse conditions. Sci. Agricola 2011, 68, 400–405. [Google Scholar] [CrossRef]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of Waterlogging Tolerance in Plants: Research Progress and Prospects. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef]
- Steffens, B.; Rasmussen, A. The Physiology of Adventitious Roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Ayi, Q.; Zeng, B.; Liu, J.; Qiaoli, A.; van Bodegom, P.; Cornelissen, J.H.C. Oxygen absorption by adventitious roots promotes the survival of completely submerged terrestrial plants. Ann. Bot. 2016, 118, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Gonin, M.; Bergougnoux, V.; Nguyen, T.D.; Gantet, P.; Champion, A. What Makes Adventitious Roots? Plants 2019, 8, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Wolf, S.; Kafkafi, U. Interactive effect of nutrient concentration and container volume on flowering, fruiting, and nutrient uptake of sweet pepper. J. Plant Nutr. 2001, 24, 479–501. [Google Scholar] [CrossRef]
- Bar-Tal, A.; Feigin, A.; Sheinfeld, S.; Rosenberg, R.; Sternbaum, B.; Rylski, I.; Pressman, E. Root restriction and N-NO3 solution concentration effects on nutrient uptake, transpiration and dry matter production of tomato. Sci. Hortic. 1995, 63, 195–208. [Google Scholar] [CrossRef]
- Massa, D.; Magán, J.J.; Montesano, F.F.; Tzortzakis, N. Minimizing water and nutrient losses from soilless cropping in southern Europe. Agric. Water Manag. 2020, 241, 106395. [Google Scholar] [CrossRef]
- Shi, K.; Ding, X.-T.; Dong, D.-K.; Zhou, Y.-H.; Yu, J.-Q. Root restriction-induced limitation to photosynthesis in tomato (Lycopersicon esculentum Mill.) leaves. Sci. Hortic. 2008, 117, 197–202. [Google Scholar] [CrossRef]
- Nishizawa, T.; Saito, K. Effects of Rooting Volume Restriction on the Growth and Carbohydrate Concentration in Tomato Plants. J. Am. Soc. Hortic. Sci. 1998, 123, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Pezeshki, S.; Santos, M. Relationships among Rhizosphere Oxygen Deficiency, Root Restriction, Photosynthesis, and Growth in Baldcypress (Taxodium distichum L.) Seedlings. Photosynthrtica 1998, 35, 381–390. [Google Scholar] [CrossRef]
- Osorio, S.; Ruan, Y.-L.; Fernie, A.R. An update on source-to-sink carbon partitioning in tomato. Front. Plant Sci. 2014, 5, 516. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [Green Version]
- Bar-Yosef, B. Fertigation management and crop response to solution recycling in semi-closed greenhouses. In Soilless Culture: Theory and Practice; Raviv, M., Lieth, J.H., Eds.; Elsevier: Alpharetta, GA, USA, 2008; pp. 545–573. [Google Scholar]
- Lynch, J. Root Architecture and Plant Productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2015, 58, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forde, B.; Lorenzo, H. The nutritional control of root development. Plant Soil 2001, 232, 51–68. [Google Scholar] [CrossRef]
- Shahzad, Z.; Amtmann, A. Food for thought: How nutrients regulate root system architecture. Curr. Opin. Plant Biol. 2017, 39, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Awika, H.O.; Mishra, A.K.; Gill, H.; DiPiazza, J.; Avila, C.A.; Joshi, V. Selection of nitrogen responsive root architectural traits in spinach using machine learning and genetic correlations. Sci. Rep. 2021, 11, 9536. [Google Scholar] [CrossRef]
- Marzec, M.; Melzer, M. Regulation of Root Development and Architecture by Strigolactones under Optimal and Nutrient Deficiency Conditions. Int. J. Mol. Sci. 2018, 19, 1887. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2012, 112, 391–408. [Google Scholar] [CrossRef]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Gruda, N.; Schnitzler, W.H. Transplants Influence of Wood Fiber Substrates and N Application Rates on the Growth of Tomato Transplants. Adv. Hortic. Sci. 1999, 13, 20–24. [Google Scholar]
- Kellermeier, F.; Armengaud, P.; Seditas, T.J.; Danku, J.; Salt, D.E.; Amtmann, A. Analysis of the Root System Architecture of Arabidopsis Provides a Quantitative Readout of Crosstalk between Nutritional Signals. Plant Cell 2014, 26, 1480–1496. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.L.; Andriolo, J.L.; Picio, M.D.; Piccin, M.; Souza, J.M. Nitrogen on growth and yield of lettuce plants grown under root confinement. Hortic. Bras. 2015, 33, 422–427. [Google Scholar] [CrossRef]
- Öztekin, G.B.; Uludağ, T.; Tüzel, Y. Growing spinach (Spinacia oleracea L.) in a floating system with different concentrations of nutrient solution. Appl. Ecol. Environ. Res. 2018, 16, 3333–3350. [Google Scholar] [CrossRef]
- Balanza, V.; Martínez, J.; Conesa, E.; Egea-Gilabert, C.; Niñirola, D.; Marin, J.L.; González, A.; Fernández, J.A. Effect of pgpr application and nitrogen doses on baby leaf lettuce grown in a floating system. Acta Hortic. 2012, 952, 679–687. [Google Scholar] [CrossRef]
- Roberts, B.R.; Wolverton, C.; Janowicz, L. The impact of substrate and irrigation interval on the post-transplant root growth of container-grown zinnia and tomato1. J. Environ. Hortic. 2017, 35, 1–5. [Google Scholar] [CrossRef]
- Ismail, S.M.; Ozawa, K. Effect of Irrigation Interval on Growth Characteristics, Plant Water Stress Tolerance and Water Use Efficiency for Chile Pepper. In Proceedings of the Thirteenth International Water Technology Conference (IWTC 130), Hurghada, Egypt, 12–15 March 2009; pp. 545–556. [Google Scholar]
- Silber, A.; Xu, G.; Wallach, R. High irrigation frequency: The effect on plant growth and on uptake of water and nutrients. Acta Hortic. 2003, 627, 89–96. [Google Scholar] [CrossRef]
- Grewal, H.S.; Maheshwari, B.; Parks, S. Water and nutrient use efficiency of a low-cost hydroponic greenhouse for a cucumber crop: An Australian case study. Agric. Water Manag. 2011, 98, 841–846. [Google Scholar] [CrossRef]
- Yang, T.; Kim, H.-J. Comparisons of nitrogen and phosphorus mass balance for tomato-, basil-, and lettuce-based aquaponic and hydroponic systems. J. Clean. Prod. 2020, 274, 122619. [Google Scholar] [CrossRef]
- LE Bot, J.; Jeannequin, B.; Fabre, R. Growth and Nitrogen Status of Soilless Tomato Plants Following Nitrate Withdrawal from the Nutrient Solution. Ann. Bot. 2001, 88, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.; Savvas, D.; Colla, G.; Rouphael, Y. Impacts of genetic material and current technologies on product quality of selected greenhouse vegetables—A review. Eur. J. Hortic. Sci. 2018, 83, 319–328. [Google Scholar] [CrossRef]
- Conversa, G.; Bonasia, A.; Lazzizera, C.; La Rotonda, P.; Elia, A. Reduction of Nitrate Content in Baby-Leaf Lettuce and Cichorium endivia Through the Soilless Cultivation System, Electrical Conductivity and Management of Nutrient Solution. Front. Plant Sci. 2021, 12, 645671. [Google Scholar] [CrossRef]
- Lieth, J.H.; Oki, L.R. Irrigation in soilless production. In Soilless Culture: Theory and Practice Theory and Practice; Elsevier BV: Edinburgh, UK, 2019. [Google Scholar] [CrossRef]
- Nikolaou, G.; Neocleous, D.; Katsoulas, N.; Kittas, C. Irrigation of Greenhouse Crops. Horticulturae 2019, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Silber, A. Fertigation frequency and nutrient uptake by plants: Benefits and constraints. Int. Fertil. Soc. Proc. 2005, 571, 1–36. [Google Scholar]
- Taweesak, V.; Abdullah, T.L.; Hassan, S.A.; Kamarulzaman, N.H.; Yusoff, W.A.W. Growth and Flowering Responses of Cut Chrysanthemum Grown under Restricted Root Volume to Irrigation Frequency. Sci. World J. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Ortega, W.; Martinez, V.; Rivero, R.M.; Cámara-Zapata, J.-M.; Mestre, T.; Garcia-Sanchez, F. Use of a smart irrigation system to study the effects of irrigation management on the agronomic and physiological responses of tomato plants grown under different temperatures regimes. Agric. Water Manag. 2017, 183, 158–168. [Google Scholar] [CrossRef]
- Gruda, N.; Schnitzler, W.H. The effect of water supply on bio-morphological and plant-physiological parameters of tomato transplants cultivated in wood fiber substrate. J. Appl. Bot. 2000, 74, 233–239. [Google Scholar] [CrossRef]
- Leskovar, D.I. Root and Shoot Modification by Irrigation. HortTechnology 1998, 8, 510–514. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.; Schnitzler, W. Suitability of wood fiber substrates for production of vegetable transplants II: The effect of wood fiber substrates and their volume weights on the growth of tomato transplants. Sci. Hortic. 2004, 100, 333–340. [Google Scholar] [CrossRef]
- Gruda, N.; Schnitzler, W. Suitability of wood fiber substrate for production of vegetable transplants: I. Physical properties of wood fiber substrates. Sci. Hortic. 2004, 100, 309–322. [Google Scholar] [CrossRef]
- Machado, R.; Alves-Pereira, I.; Ferreira, R.; Gruda, N. Coir, an Alternative to Peat—Effects on Plant Growth, Phytochemical Accumulation, and Antioxidant Power of Spinach. Horticulturae 2021, 7, 127. [Google Scholar] [CrossRef]
- Wallach, R. Physical Characteristics of Soilless Media. In Soilless Culture: Theory and Practice; Raviv, M., Lieth, J.H., Eds.; Elsevier: Alpharetta, GA, USA, 2008; pp. 41–116. [Google Scholar]
- Xu, G.; Levkovitch, I.; Soriano, S.; Wallach, R.; Silber, A. Integrated effect of irrigation frequency and phosphorus level on lettuce: P uptake, root growth and yield. Plant Soil 2004, 263, 297–309. [Google Scholar] [CrossRef]
- Scagel, C.F.; Bi, G.; Fuchigami, L.H.; Regan, R.P. Effects of Irrigation Frequency and Nitrogen Fertilizer Rate on Water Stress, Nitrogen Uptake, and Plant Growth of Container-grown Rhododendron. HortScience 2011, 46, 1598–1603. [Google Scholar] [CrossRef] [Green Version]
- Nikolaou, G.; Neocleous, D.; Katsoulas, N.; Kittas, C. Effect of irrigation frequency on growth and production of a cucumber crop under soilless culture. Emir. J. Food Agric. 2017, 29, 863. [Google Scholar] [CrossRef] [Green Version]
- Heller, H.; Bar-Tal, A.; Assouline, S.; Narkis, K.; Suryano, S.; De La Forge, A.; Barak, M.; Alon, H.; Bruner, M.; Cohen, S.; et al. The effects of container geometry on water and heat regimes in soilless culture: Lettuce as a case study. Irrig. Sci. 2014, 33, 53–65. [Google Scholar] [CrossRef]
- Fields, J.S.; Owen, J.; Scoggins, H.L. The Influence of Substrate Hydraulic Conductivity on Plant Water Status of an Ornamental Container Crop Grown in Suboptimal Substrate Water Potentials. HortScience 2017, 52, 1419–1428. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.; Rau, B.J.; Wright, R.D. Laboratory bioassay and greenhouse evaluation of a pine tree substrate used as a container substrate. Eur. J. Hortic. Sci. 2009, 74, 73–78. [Google Scholar]
- Ortega, M.; Moreno, M.T.; Ordovas, J.; Aguado, M. Behaviour of different horticultural species in phytotoxicity bioassays of bark substrates. Sci. Hortic. 1996, 66, 125–132. [Google Scholar] [CrossRef]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots Withstanding their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N. Impact of Environmental Factors on Product Quality of Greenhouse Vegetables for Fresh Consumption. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, K.-X.; Wang, W.-S.; Gong, W.; Liu, W.-C.; Chen, H.-G.; Xu, H.-H.; Lu, Y.-T. Low Temperature Inhibits Root Growth by Reducing Auxin Accumulation via ARR1/12. Plant Cell Physiol. 2014, 56, 727–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Song, F.; Liu, F.; Tian, C.; Liu, S.; Xu, H.; Zhu, X. Effect of Different Arbuscular Mycorrhizal Fungi on Growth and Physiology of Maize at Ambient and Low Temperature Regimes. Sci. World J. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nagel, K.A.; Kastenholz, B.; Jahnke, S.; van Dusschoten, D.; Aach, T.; Mühlich, M.; Truhn, D.; Scharr, H.; Terjung, S.; Walter, A.; et al. Temperature responses of roots: Impact on growth, root system architecture and implications for phenotyping. Funct. Plant Biol. 2009, 36, 947–959. [Google Scholar] [CrossRef]
- Du, Y.; Scheres, B. Lateral root formation and the multiple roles of auxin. J. Exp. Bot. 2017, 69, 155–167. [Google Scholar] [CrossRef]
- Waidmann, S.; Sarkel, E.; Kleine-Vehn, J. Same same, but different: Growth responses of primary and lateral roots. J. Exp. Bot. 2020, 71, 2397–2411. [Google Scholar] [CrossRef]
- Torres-Martínez, H.H.; Rodriguez-Alonso, G.; Shishkova, S.; Dubrovsky, J.G. Lateral Root Primordium Morphogenesis in Angiosperms. Front. Plant Sci. 2019, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.-Y.; Duan, Z.-Q.; Mao, J.-D.; Li, X.; Dong, F. Low Root Zone Temperature Limits Nutrient Effects on Cucumber Seedling Growth and Induces Adversity Physiological Response. J. Integr. Agric. 2013, 12, 1450–1460. [Google Scholar] [CrossRef]
- Luo, H.; Xu, H.; Chu, C.; He, F.; Fang, S. High Temperature can Change Root System Architecture and Intensify Root Interactions of Plant Seedlings. Front. Plant Sci. 2020, 11, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balliu, A.; Sallaku, G. The environment temperature affects post-germination growth and root system architecture of pea (Pisum sativum L.) plants. Sci. Hortic. 2021, 278, 109858. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Effect of Root-Zone Temperature on Growth and Quality of Hydroponically Grown Red Leaf Lettuce (Lactuca sativa L. cv. Red Wave). Am. J. Plant Sci. 2015, 6, 2350–2360. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Thiele, B.; Watt, M.; Kraska, T.; Ulbrich, A.; Kuhn, A.J. Effects of Root Cooling on Plant Growth and Fruit Quality of Cocktail Tomato during Two Consecutive Seasons. J. Food Qual. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Matsuo, S.; Kanayama, Y.; Kanahama, K. Effect of Root-zone Heating on Root Growth and Activity, Nutrient Uptake, and Fruit Yield of Tomato at Low Air Temperatures. J. Jpn. Soc. Hortic. Sci. 2014, 83, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Karnoutsos, P.; Karagiovanidis, M.; Bantis, F.; Chatzistathis, T.; Koukounaras, A.; Ntinas, G.K. Controlled root-zone temperature effect on baby leaf vegetables yield and quality in a floating system under mild and extreme weather conditions. J. Sci. Food Agric. 2020, 101, 3933–3941. [Google Scholar] [CrossRef]
- Qin, L.; He, J.; Lee, S.K.; Dodd, I.C. An assessment of the role of ethylene in mediating lettuce (Lactuca sativa) root growth at high temperatures. J. Exp. Bot. 2007, 58, 3017–3024. [Google Scholar] [CrossRef] [Green Version]
- Pardales, J.R.J.; Banoc, D.M.; Yamauchi, A.; Iijima, M.; Kono, Y. Root System Development of Cassava and Sweetpotato during Early Growth Stage as Affected by High Root Zone Temperature. Plant Prod. Sci. 1999, 2, 247–251. [Google Scholar] [CrossRef]
- MacDuff, J.H.; Wild, A.; Hopper, M.J.; Dhanoa, M.S. Effects of temperature on parameters of root growth relevant to nutrient uptake: Measurements on oilseed rape and barley grown in flowing nutrient solution. Plant Soil 1986, 94, 321–332. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Miao, Y.; Gao, L. Root-Zone Warming Differently Benefits Mature and Newly Unfolded Leaves of Cucumis sativus L. Seedlings under Sub-Optimal Temperature Stress. PLoS ONE 2016, 11, e0155298. [Google Scholar] [CrossRef] [Green Version]
- Aroca, R.; Porcel, R.; Lozano, J.M.R. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2011, 63, 43–57. [Google Scholar] [CrossRef]
- Kafkafi, U. Functions of the Root System. In Soilless Culture: Theory and Practice; Raviv, M., Lieth, J.H., Eds.; Elsevier: Alpharetta, GA, USA, 2008; pp. 13–40. [Google Scholar]
- Poiré, R.; Schneider, H.; Thorpe, M.R.; Kuhn, A.J.; Schurr, U.; Walter, A. Root cooling strongly affects diel leaf growth dynamics, water and carbohydrate relations inRicinus communis. Plant Cell Environ. 2010, 33, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.; He, J.; Turnbull, C.; Lee, S.; Critchley, C. The influence of supra-optimal root-zone temperatures on growth and stomatal conductance in Capsicum annuum L. J. Exp. Bot. 2000, 51, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, S. Effect of Root Temperature on the Rate of Water and Nutrient Absorption in Cucumber Cultivars and Figleaf Gourd. J. Jpn. Soc. Hortic. Sci. 1987, 55, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, Y.; Matsuo, S.; Suzuki, K.; Kanayama, Y.; Kanahama, K. Root-zone Cooling at High Air Temperatures Enhances Physiological Activities and Internal Structures of Roots in Young Tomato Plants. J. Jpn. Soc. Hortic. Sci. 2013, 82, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Gruda, N.; Dong, J.; Duan, Z. Greenhouse soil warmed by capillary network and its effect on the growth of cucumber. Acta Hortic. 2020, 1296, 149–158. [Google Scholar] [CrossRef]
- Masaru, S.; Uenishi, M.; Miyamoto, K.; Suzuki, T. Effect of Root-Zone Temperature on the Growth and Fruit Quality of Hydroponically Grown Strawberry Plants. J. Agric. Sci. 2016, 8, 122. [Google Scholar] [CrossRef]
- Falah, M.A.F.; Wajima, T.; Yasutake, D.; Sago, Y.; Kitano, M. Responses of root uptake to high temperature of tomato plants (Lycopersicon esculentum Mill.) in soil-less culture. J. Agric. Technol. 2010, 6, 543–558. [Google Scholar]
- Giri, A.; Heckathorn, S.; Mishra, S.; Krause, C. Heat Stress Decreases Levels of Nutrient-Uptake and -Assimilation Proteins in Tomato Roots. Plants 2017, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Suzuki, T. Elevated Root-Zone Temperature Modulates Growth and Quality of Hydroponically Grown Carrots. Agric. Sci. 2015, 06, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, O.; Sauter, M.; Colmer, T.D.; Nakazono, M. Regulation of root adaptive anatomical and morphological traits during low soil oxygen. New Phytol. 2021, 229, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Niu, W.; Cao, X.; Wang, J.; Zhang, M.; Duan, X.; Zhang, Z. Effect of soil aeration on root morphology and photosynthetic characteristics of potted tomato plants (Solanum lycopersicum) at different NaCl salinity levels. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Morard, P.; Silvestre, J. Plant injury due to oxygen deficiency in the root environment of soilless culture: A review. Plant Soil 1996, 184, 243–254. [Google Scholar] [CrossRef]
- Nicola, S.; Egea-Gilabert, C.; Niñirola, D.; Conesa, E.; Pignata, G.; Fontana, E.; Fernández, J.A. Nitrogen and aeration levels of the nutrient solution in soilless cultivation systems as important growing conditions affecting inherent quality of baby leaf vegetables: A review. Acta Hortic. 2015, 1099, 167–177. [Google Scholar] [CrossRef]
- Shaw, R.E.; Meyer, W.S.; McNeill, A.; Tyerman, S.D. Waterlogging in Australian agricultural landscapes: A review of plant responses and crop models Waterlogging in Australian agricultural landscapes: A review of plant responses and crop models. Crop. Pasture Sci. 2013, 64, 549–562. [Google Scholar] [CrossRef]
- Lara, L.J.; Egea-Gilabert, C.; Niñirola, D.; Conesa, E.; Fernández, J.A. Effect of aeration of the nutrient solution on the growth and quality of purslane (Portulaca oleracea). J. Hortic. Sci. Biotechnol. 2011, 86, 603–610. [Google Scholar] [CrossRef]
- Jackson, M.B. Long-distance signalling from roots to shoots assessed: The flooding story. J. Exp.Bot. 2002, 53, 175–181. [Google Scholar] [CrossRef]
- Acuña, R.; Gil, I.; Bonachela, S.; Magán, J. Oxyfertigation of a greenhouse melon crop grown in rockwool slabs in a mediterranean area. Acta Hortic. 2008, 779, 447–454. [Google Scholar] [CrossRef]
- Bonachela, S.; Acuña, R.A.; Magan, J.J.; Marfa, O. Oxygen enrichment of nutrient solution of substrate-grown vegetable crops under Mediterranean greenhouse conditions: Oxygen content dynamics and crop response. Span. J. Agric. Res. 2010, 8, 1231. [Google Scholar] [CrossRef]
- Öztekin, G.B.; Tüzel, Y. Effects of oxyfertigation and plant growth promoting rhizobacteria on greenhouse lettuce grown in perlite. Acta Sci. Pol. Hortorum. Cultus 2020, 19, 97–105. [Google Scholar] [CrossRef]
- Schroeder, F.-G.; Lieth, J. Gas composition and oxygen supply in the root environment of substrates in closed hydroponic systems. Acta Hortic. 2004, 644, 299–305. [Google Scholar] [CrossRef]
- Suhl, J.; Oppedijk, B.; Baganz, D.; Kloas, W.; Schmidt, U.; van Duijn, B. Oxygen consumption in recirculating nutrient film technique in aquaponics. Sci. Hortic. 2019, 255, 281–291. [Google Scholar] [CrossRef]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Annu. Rev. Plant Physiol.Plant Mol. Biol. 1997, 48, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Blok, C.; Jackson, B.E.; Guo, X.; de Visser, P.H.B.; Marcelis, L.F.M. Maximum Plant Uptakes for Water, Nutrients, and Oxygen Are Not Always Met by Irrigation Rate and Distribution in Water-based Cultivation Systems. Front. Plant Sci. 2017, 8, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morard, P.; Lacoste, L.; Silvestre, J. Effect of oxygen deficiency on uptake of water and mineral nutrients by tomato plants in soilless culture. J. Plant Nutr. 2000, 23, 1063–1078. [Google Scholar] [CrossRef]
- Alvarado-Camarillo, D.; Valdez-Aguilar, L.A.; González-Fuentes, J.A.; Rascón-Alvarado, E.; Peña-Ramos, F.M. Response of hydroponic lettuce to aeration, nitrate and potassium in the nutrient solution. Acta Agric. Scand. Sect. B—Plant Soil Sci. 2020, 70, 341–348. [Google Scholar] [CrossRef]
- Marfà, O.; Cáceres, R.; Guri, S. Oxyfertigation: A new technique for soilless culture under mediterranean conditions. Acta Hortic. 2005, 697, 65–72. [Google Scholar] [CrossRef]
- Ehret, D.L.; Edwards, D.; Helmer, T.; Lin, W.; Jones, G.; Dorais, M.; Papadopoulos, A.P. Effects of oxygen-enriched nutrient solution on greenhouse cucumber and pepper production. Sci. Hortic. 2010, 125, 602–607. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, B.S.; Kang, J.G.; Bae, J.H.; Ku, Y.G.; Gorinstein, S.; Lee, J.H. Effect of root zone aeration on the growth and bioactivity of cucumber plants cultured in perlite substrate. Biologia 2014, 69, 610–617. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, L.; Dixon, M. An upper limit for elevated root zone dissolved oxygen concentration for tomato. Sci. Hortic. 2007, 113, 162–165. [Google Scholar] [CrossRef]
- Zhang, J.; Jiao, X.; Du, Q.; Song, X.; Ding, J.; Li, J. Effects of Vapor Pressure Deficit and Potassium Supply on Root Morphology, Potassium Uptake, and Biomass Allocation of Tomato Seedlings. J. Plant Growth Regul. 2021, 40, 509–518. [Google Scholar] [CrossRef]
- Arve, L.E.; Torre, S. Ethylene is involved in high air humidity promoted stomatal opening of tomato (Lycopersicon esculentum) leaves. Funct. Plant Biol. 2015, 42, 376–386. [Google Scholar] [CrossRef]
- Lihavainen, J.; Ahonen, V.; Keski-Saari, S.; Kontunen-Soppela, S.; Oksanen, E.; Keinänen, M. Low vapour pressure deficit affects nitrogen nutrition and foliar metabolites in silver birch. J. Exp. Bot. 2016, 67, 4353–4365. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Shimizu, H. Plant growth and water use efficiency of four Chinese conifer tree species under different air humidity. Phyton-Horn 2005, 45, 575–582. [Google Scholar]
- Adams, P. Crop nutrition in hydroponics. Acta Hortic. 1993, 323, 289–306. [Google Scholar] [CrossRef]
- Christiaens, A.; Gobin, B.; Van Labeke, M. Light quality and adventitious rooting: A mini-review. Acta Hortic. 2016, 1134, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Gil, C.S.; Jung, H.Y.; Lee, C.; Eom, S.H. Blue light and NAA treatment significantly improve rooting on single leaf-bud cutting of Chrysanthemum via upregulated rooting-related genes. Sci. Hortic. 2020, 274, 109650. [Google Scholar] [CrossRef]
- Rasool, A.; Graham, T.; Dayboll, C.; Stoochnoff, J.; Zheng, Y.; Dixon, M. Improving adventitious rooting in Kalanchoe blossfeldiana cuttings using sole source targeted LED lighting spectra. Acta Hortic. 2020, 1296, 629–636. [Google Scholar] [CrossRef]
- Navidad, H.; Fløistad, I.S.; Olsen, J.E.; Torre, S. Subalpine Fir (Abies laciocarpa) and Norway Spruce (Piceaabies) Seedlings Show Different Growth Responses to Blue Light. Agronomy 2020, 10, 712. [Google Scholar] [CrossRef]
- Olschowski, S.; Geiger, E.-M.; Herrmann, J.; Sander, G.; Grüneberg, H. Effects of red, blue, and white LED irradiation on root and shoot development of Calibrachoa cuttings in comparison to high pressure sodium lamps. Acta Hortic. 2016, 1134, 245–250. [Google Scholar] [CrossRef]
- Barickman, T.; Kopsell, D.; Sams, C.; Morrow, R. Sole-Source LED Lighting and Fertility Impact Shoot and Root Tissue Mineral Elements in Chinese Kale (Brassica oleracea var. alboglabra). Horticulturae 2020, 6, 40. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, X.; Wei, H.-X.; Lv, J.; Chen, C.; Liu, X.-Y.; Wen, Q.; Jia, L.-M. Nutrient uptake and utilization in Prince Rupprecht’s larch (Larix principis-rupprechtii Mayr.) seedlings exposed to a combination of light-emitting diode spectra and exponential fertilization. Soil Sci. Plant Nutr. 2019, 65, 358–368. [Google Scholar] [CrossRef]
- Zheng, Y. Advances in understanding plant root behaviour and rootzone management in soilless culture system. In Advances in Horticultural Soilless Culture; Gruda, N.S., Ed.; Burleigh Dodds Science: Cambridge, UK, 2021; pp. 23–44. [Google Scholar]
- Li, X.; Dong, J.; Gruda, N.; Chu, W.; Duan, Z. Does the short-term fluctuation of mineral element concentrations in the closed hydroponic experimental facilities affect the mineral concentrations in cucumber plants exposed to elevated CO2? Plant Soil 2021, 1–17. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Li, X.; Tang, Y.; Zhang, P.; Duan, Z. Sustainable vegetable production under changing climate: The impact of elevated CO2 on yield of vegetables and the interactions with environments-A review. J. Clean. Prod. 2020, 253, 119920. [Google Scholar] [CrossRef]
- Li, X.; Dong, J.; Chu, W.; Chen, Y.; Duan, Z. The relationship between root exudation properties and root morphological traits of cucumber grown under different nitrogen supplies and atmospheric CO2 concentrations. Plant Soil 2018, 425, 415–432. [Google Scholar] [CrossRef]
- Dyśko, J.; Kaniszewski, S.; Kowalczyk, W. The effect of nutrient solution on phosphoros availability in soilless culture of tomato. J. Elementol. 2008, 13, 189–198. [Google Scholar]
- Dyśko, J.; Kowalczyk, W.; Kaniszewski, S. The Influence of pH of Nutrient Solution on Yield and Nutritional Status of Tomato Plants Grown in Soilless Culture System. Veg. Crop. Res. Bull. 2009, 70, 59–69. [Google Scholar] [CrossRef]
- Savvas, D.; Karagianni, V.; Kotsiras, A.; Demopoulos, V.; Karkamisi, I.; Pakou, P. Interactions between ammonium and pH of the nutrient solution supplied to gerbera (Gerbera jamesonii) grown in pumice. Plant Soil 2003, 254, 393–402. [Google Scholar] [CrossRef]
- Huang, J.-S.; Nelson, P.V.; Lee, J.-W. Seedling effect on root substrate pH. J. Plant Nutr. 2001, 24, 1133–1147. [Google Scholar] [CrossRef]
- Zheng, Y.; Dixon, M.; Saxena, P. Greenhouse production of Echinacea purpurea (L.) and E. angustifolia using different growing media, NO3−/NH4+ ratios and watering regimes. Can. J. Plant Sci. 2006, 86, 809–815. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, F.; Zwiazek, J.J. Responses of jack pine (Pinus banksiana) seedlings to root zone pH and calcium. Environ. Exp. Bot. 2015, 111, 32–41. [Google Scholar] [CrossRef]
- Strehmel, N.; Böttcher, C.; Schmidt, S.; Scheel, D. Profiling of secondary metabolites in root exudates of Arabidopsis thaliana. Phytochemistry 2014, 108, 35–46. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. Allelochemical stress inhibits growth, leaf water relations, PSII photochemistry, non-photochemical fluorescence quenching, and heat energy dissipation in three C3 perennial species. J. Exp. Bot. 2011, 62, 4533–4545. [Google Scholar] [CrossRef]
- Talukder, R.; Asaduzzaman, M.; Ueno, M.; Tanaka, H.; Asao, T. Alleviation of allelochemical stress-induced growth inhibition and oxidative damage in lettuce under closed hydroponics through electro-degradation. Hortic. Sci. 2020, 47, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, S.; Kumar, P. Rhizosphere microbiome: Revisiting the synergy of plant-microbe interactions. Ann. Microbiol. 2019, 69, 307–320. [Google Scholar] [CrossRef]
- Nazir, N.; Kamili, A.N.; Zargar, M.Y.; Khan, I.; Shah, D.; Parray, J.A.; Tyub, S. Effect of Root Exudates on Rhizosphere Soil Microbial Communities. J. Res. Dev. 2016, 16, 88–95. [Google Scholar]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O.; Ayangbenro, A. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Weir, T.L.; Perry, L.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Baum, C.; El-Tohamy, W.; Gruda, N. Increasing the productivity and product quality of vegetable crops using arbuscular mycorrhizal fungi: A review. Sci. Hortic. 2015, 187, 131–141. [Google Scholar] [CrossRef]
- Berkelmann, B.; Wohanka, W.; Wolf, G. Characterization of the bacterial flor in circulating nutrient solutions of a hydroponic system with rockwool. Acta Hortic. 1994, 372–381. [Google Scholar] [CrossRef]
- Stanghellini, M.E. Hydroponics: A Solution for Zoosporic Pathogens. Plant Dis. 1994, 78, 1129–1138. [Google Scholar] [CrossRef]
- Chave, M.; Dabert, P.; Brun, R.; Godon, J.-J.; Poncet, C. Dynamics of rhizoplane bacterial communities subjected to physicochemical treatments in hydroponic crops. Crop. Prot. 2008, 27, 418–426. [Google Scholar] [CrossRef]
- Koohakan, P.; Ikeda, H.; Jeanaksorn, T.; Tojo, M.; Kusakari, S.-I.; Okada, K.; Sato, S. Evaluation of the indigenous microorganisms in soilless culture: Occurrence and quantitative characteristics in the different growing systems. Sci. Hortic. 2004, 101, 179–188. [Google Scholar] [CrossRef]
- Khalil, S.; Alsanius, B.W. Dynamics of the indigenous microflora inhabiting the root zone and the nutrient solution of tomato in a commercial closed greenhouse system. Gartenbauwissenschaft 2001, 66, 188–198. [Google Scholar]
- Waechter-Kristensen, B.; Sundin, P.; Gertsson, U.; Hultberg, M.; Khalil, S.; Jensén, P.; Berkelmann-Loehnertz, B.; Wohanka, W. Management of microbial factors in the rhizosphere and nutrient solution of hydroponically grown tomato. Acta Hortic. 1997, 335–342. [Google Scholar] [CrossRef]
- Babaj, I.; Sallaku, G.; Balliu, A. The effects of endogenous mycorrhiza (Glomus spp.) on plant growth and yield of grafted cucumber (Cucumis sativum L.) under common commercial greenhouse conditions. Albanian J. Agric. Sci. 2014, 13, 24–28. [Google Scholar]
- Vuksani, A.; Sallaku, G.; Balliu, A. The Effects of Endogenous Mycorrhiza (Glomus spp.) on Stand Establishment Rate and Yield of Open Field Tomato Crop. Albanian J. Agric. Sci. 2015, 14, 25–30. [Google Scholar]
- Unal, N.; Tuzel, Y.; Tuzel, I.; Oztekin, G. Effects of mycorrhiza and irrigation programs on strawberry production in substrate culture. Acta Hortic. 2020, 1273, 177–182. [Google Scholar] [CrossRef]
- Balliu, A.; Sallaku, G.; Rewald, B. AMF Inoculation Enhances Growth and Improves the Nutrient Uptake Rates of Transplanted, Salt-Stressed Tomato Seedlings. Sustainability 2015, 7, 15967–15981. [Google Scholar] [CrossRef] [Green Version]
- Meça, E.; Sallaku, G.; Balliu, A. Artificial Inoculation of AM Fungi Improves Nutrient Uptake Efficiency in Salt Stressed Pea (Pissum Sativum L.) Plants. J. Agric. Stud. 2016, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.; Ellouze, W.; Howard, R.J. Utility of Arbuscular Mycorrhizal Fungi for Improved Production and Disease Mitigation in Organic and Hydroponic Greenhouse Crops. J. Hortic. 2018, 5, 1–10. [Google Scholar] [CrossRef]
- Shao, Y.-D.; Hu, X.-C.; Wu, Q.-S.; Yang, T.-Y.; Srivastava, A.; Zhang, D.-J.; Gao, X.-B.; Kuča, K. Mycorrhizas promote P acquisition of tea plants through changes in root morphology and P transporter gene expression. S. Afr. J. Bot. 2021, 137, 455–462. [Google Scholar] [CrossRef]
- Nurbaity, A.; Istifadah, N.; A Haryantini, B.; Ilhami, M.F.; I Habibullah, M.; Arifin, M. Optimization of hydroponic technology for production of mycorrhiza biofertilizer. IOP Conf. Series: Earth Environ. Sci. 2019, 347, 012017. [Google Scholar] [CrossRef]
- Utkhede, R. Increased Growth and Yield of Hydroponically Grown Greenhouse Tomato Plants Inoculated with Arbuscular Mycorrhizal Fungi and Fusarium oxysporum f. sp. radicis-lycopersici. BioControl 2006, 51, 393–400. [Google Scholar] [CrossRef]
- Postma, J.; Klein, M.J.E.I.M.W.-D.; Van Elsas, J.D. Effect of the Indigenous Microflora on the Development of Root and Crown Rot Caused by Pythium aphanidermatum in Cucumber Grown on Rockwool. Phytopathology 2000, 90, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Minuto, A.; Clematis, F.; Gullino, M.L.; Garibaldi, A. Induced suppressiveness to Fusarium oxysporumf.sp. radicis lycopersici in rockwool substrate used in closed soilless systems. Phytoparasitica 2007, 35, 77–85. [Google Scholar] [CrossRef]
- Garibaldi, A.; Minuto, A.; Grasso, V.; Gullino, M. Application of selected antagonistic strains against Phytophthora cryptogea on gerbera in closed soilless systems with disinfection by slow sand filtration. Crop. Prot. 2003, 22, 1053–1061. [Google Scholar] [CrossRef]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6, 786. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Glick, B.R. Mechanisms used by plant Growth-Promoting Bacteria. In Bacteria in Agrobiology: Plant Nutrient Management; Maheshwari, D.K., Ed.; Springer: Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of Salt Stress by Plant Growth-Promoting Bacteria in Hydroponic Leaf Lettuce. Agronomy 2020, 10, 1523. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J. Beneficial bacteria and fungi in hydroponic systems: Types and characteristics of hydroponic food production methods. Sci. Hortic. 2015, 195, 206–215. [Google Scholar] [CrossRef]
- Kalozoumis, P.; Savvas, D.; Aliferis, K.; Ntatsi, G.; Marakis, G.; Simou, E.; Tampakaki, A.; Karapanos, I. Impact of Plant Growth-Promoting Rhizobacteria Inoculation and Grafting on Tolerance of Tomato to Combined Water and Nutrient Stress Assessed via Metabolomics Analysis. Front. Plant Sci. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Aini, N.; Yamika, W.S.D.; Pahlevi, R.W. The effect of nutrient concentration and inoculation of PGPR and AMF on the yield and fruit quality of hydroponic cherry tomatoes (Lycopersicum esculentum Mill. var. cerasiforme). J. Appl. Hortic. 2019, 21, 116–122. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR Mediated Alterations in Root Traits: Way toward Sustainable Crop Production. Front. Sustain. Food Syst. 2021, 4, 1–28. [Google Scholar] [CrossRef]
- Veselaj, E.; Sallaku, G.; Balliu, A. Combined application of arbuscular mycorrhizae fungi and plant growth promoting bacteria improves growth and nutrient uptake efficiency of pea (Pisum sativum L.) plants. Acta Sci. Pol. Hortorum. Cultus 2018, 17, 73–86. [Google Scholar] [CrossRef]
- Veselaj, E.; Sallaku, G.; Balliu, A. Tripartite Relationships in Legume Crops Are Plant-Microorganism-Specific and Strongly Influenced by Salinity. Agriculture 2018, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- Ikiz, O.; Abak, K.; Dasgan, H.; Ortas, I. Effects of mycorrhizal inoculation in soilless culture on pepper plant growth. Acta Hortic. 2009, 807, 533–540. [Google Scholar] [CrossRef]
- Shao, Y.; Zhang, D.; Hu, X.; Wu, Q.; Jiang, K.; Xia, T.; Gao, X.; KUČA, K. Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 2018, 64, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.-N.; Zhang, D.-J.; Liu, C.-Y.; Wu, Q.-S. Relationships between mycorrhizas and root hairs. Pak. J. Bot. 2019, 51, 727–733. [Google Scholar] [CrossRef]
- Abeer, H.; Ef, A.; Alqarawi, A.A.; Mona, A. Arbuscular mycorrhizal fungi mitigates NaCl inducedects on Solanum lycopersicum L. Pak. J. Bot. 2015, 47, 327–340. [Google Scholar]
- Jahromi, F.; Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Influence of Salinity on the In Vitro Development of Glomus intraradices and on the In Vivo Physiological and Molecular Responses of Mycorrhizal Lettuce Plants. Microb. Ecol. 2007, 55, 45–53. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Jabborova, D.; Wirth, S. Alleviation of Salt Stress in Legumes by Co-inoculation with Pseudomonas and Rhizobium. In Plant Microbe Symbiosis: Fundamentals and Advances; Springer Science and Business Media LLC: New Delhi, India, 2013; pp. 291–303. [Google Scholar]
- Sallaku, G.; Sandén, H.; Babaj, I.; Kaciu, S.; Balliu, A.; Rewald, B. Specific nutrient absorption rates of transplanted cucumber seedlings are highly related to RGR and influenced by grafting method, AMF inoculation and salinity. Sci. Hortic. 2019, 243, 177–188. [Google Scholar] [CrossRef]
- Senberga, A.; Dubova, L.; Alsina, I. Germination and growth of primary roots of inoculated bean (Vicia faba) seeds under different temperatures. Agron. Res. 2018, 16, 243–253. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
| Plant Response | Crop | Production System | Additional Information | Reference |
|---|---|---|---|---|
| Reduced dry matter of roots | Chili pepper | Polyvinyl-chloride (PVC) columns, filled with a mixture of coconut coir dust and empty fruit bunch compost (70:30, v:v) | 9570 mL (control) vs. 2392 mL (root-restricted) columns | [18] |
| Pepper | Plastic pots (three seeds per pot) containing Fafard 2B mix (Sun Gro Horticulture, Agawam, MA) | 500 mL (control) vs. 60 mL (restricted) containers | [19] | |
| Cucumber | Floating system (F.S.) | Control vs. 40 mL (restricted) vessels | [20] | |
| AR formation | Cucumber | Floating system (F.S.) | Control vs. 40 mL (restricted) vessels | [20] |
| Tomato | Flow-through hydroponic culture system (FTS) | 1500 mL (control) vs. 25 mL (restricted) containers | [21] | |
| Dense mat of roots | Cucumber | Floating system (F.S.) | Control vs. 40 mL (restricted) vessels | [20] |
| Tomato | Flow-through hydroponic culture system (FTS) | 1500 mL (control) vs. 25 mL (restricted) containers | [21] | |
| Yield reduction | Sweet potato | A mixed system of solid media and nutrient solution | 4.5 L, 3.0 L, and 1.6 L pots | [22] |
| Tomato | Different alternatives of solid growing media (perlite, pumice, volcanic ash, perlite + peat, pumice + peat, volcanic ash + peat) | 8 L and 4 L pots | [23] | |
| Processing tomato | Solid growing media (Metro-Mix 350, Sun Gro Horticulture) | 26 L, 16, 6, and 1 L pots | [24] | |
| Non-significant yield reduction | Pepper | Growth media (Fafard 2B mix; Sun Gro Horticulture, and Turface clay) mixed in a 3:1 ratio | 1500 mL, 500 mL, and 250 mL plastic pots | [19] |
| Tomato | Coconut fiber substrate | 10, 7.5 and 5 L pots | [25] | |
| Increased harvest index | Pepper | Growth media (Fafard 2B mix; Sun Gro Horticulture, and Turface clay) mixed in a 3:1 ratio | 1500 mL, 500 mL, and 250 mL plastic pots | [19] |
| Chili pepper | Polyvinyl-chloride (PVC) columns, filled with a mixture of coconut coir dust and empty fruit bunch compost (70:30, v:v) | 9570 mL (control) vs. 2392 mL (root-restricted) columns | [18] |
| Plant Response | Crop | SCS | Additional Information | Reference |
|---|---|---|---|---|
| Increased irrigation frequency increases plant yield | Chrysanthemum | Seedling tray contained coconut peat | Irrigation frequencies of 4, 6, and 8 times/day | [64] |
| Tomato | 40-L (15 cm × 18 cm × 120 cm) bags containing expanded perlite | Irrigation applied when the plants had consumed 0.4-, 0.8-, or 1.2-L of water | [65] | |
| Vertical root-density distribution mimics container moisture content. Denser at the lower part of the container. | Tomato | Wood fiber substrate | [66] | |
| Chili pepper | 31 × 15 × 60 cm container filled with sandy-loamy soil | 1-, 3-, and 5-day irrigation intervals | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balliu, A.; Zheng, Y.; Sallaku, G.; Fernández, J.A.; Gruda, N.S.; Tuzel, Y. Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants. Horticulturae 2021, 7, 243. https://doi.org/10.3390/horticulturae7080243
Balliu A, Zheng Y, Sallaku G, Fernández JA, Gruda NS, Tuzel Y. Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants. Horticulturae. 2021; 7(8):243. https://doi.org/10.3390/horticulturae7080243
Chicago/Turabian StyleBalliu, Astrit, Youbin Zheng, Glenda Sallaku, Juan A. Fernández, Nazim S. Gruda, and Yuksel Tuzel. 2021. "Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants" Horticulturae 7, no. 8: 243. https://doi.org/10.3390/horticulturae7080243
APA StyleBalliu, A., Zheng, Y., Sallaku, G., Fernández, J. A., Gruda, N. S., & Tuzel, Y. (2021). Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants. Horticulturae, 7(8), 243. https://doi.org/10.3390/horticulturae7080243