
Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms

 ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Trial
2.2. Metabolite Extraction
2.3. Isolation of Volatile Fraction
2.4. LC-MS Analysis
2.5. GC-MS Analysis
2.6. Statistical Analysis of Metabolomics Data
3. Results
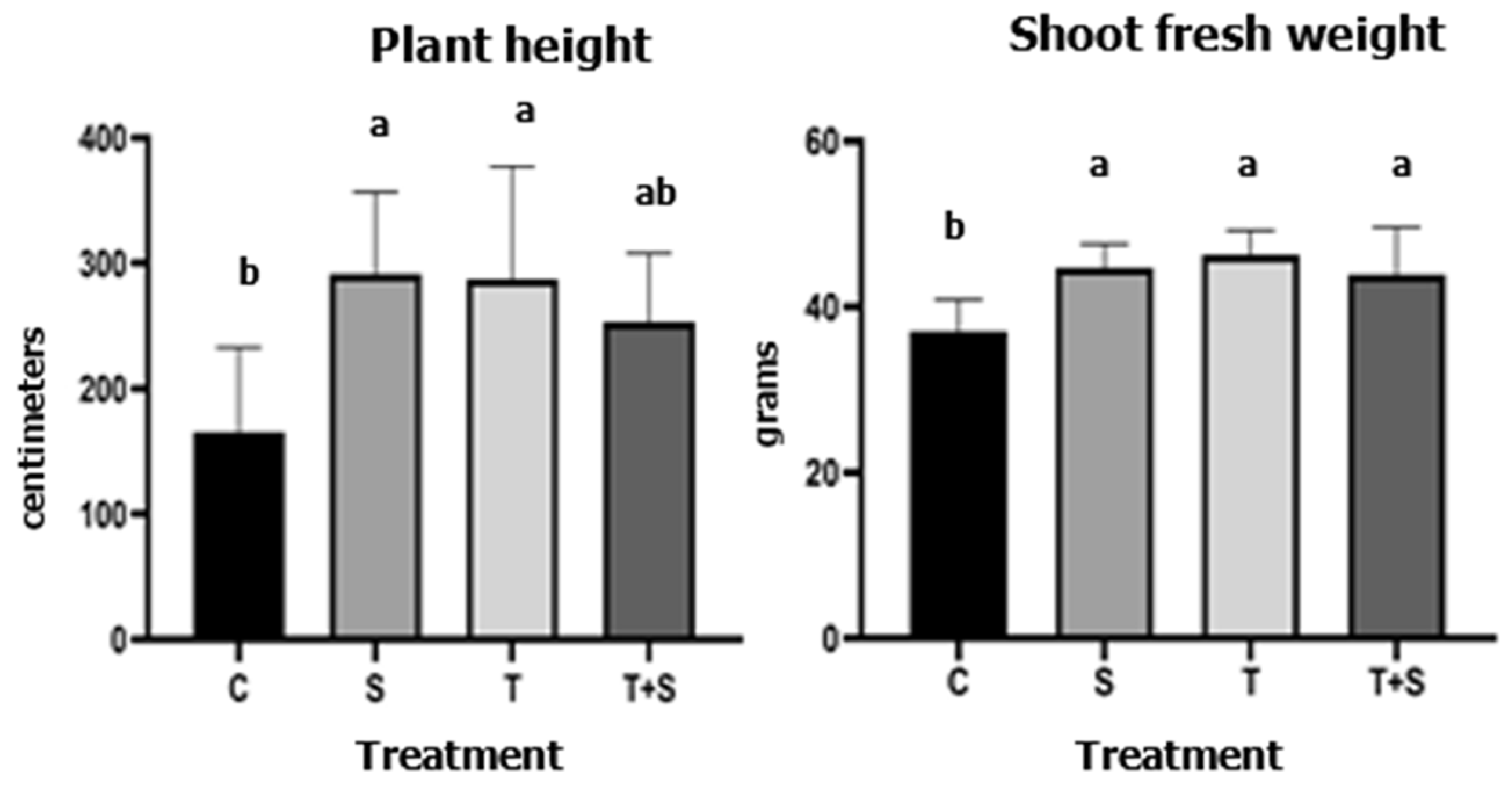
3.1. Plant Growth
3.2. Metabolomic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahmood, S.; Hussain, S.; Malik, F. Critique of medicinal conspicuousness of Parsley (Petroselinum crispum): A culinary herb of Mediterranean region. Pak. J. Pharm. Sci. 2014, 27, 193–202. [Google Scholar]
- Farzaei, M.H.; Abbasabadi, Z.; Ardekani, M.R.S.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Dini, I. Chapter 14—Spices and Herbs as Therapeutic Foods. In Handbook of Food Bioengineering, Food Quality: Balancing Health and Disease; Holban, A.M., Mihai Grumezescu, A., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 433–469. [Google Scholar]
- Dini, I.; Laneri, S. Nutricosmetics: A brief overview. Phytother. Res. 2019, 33, 3054–3063. [Google Scholar] [CrossRef] [PubMed]
- Dobricev, N.; Zabkur, J.; Voca, S.; Pliestic, S.; Galic, A.; Delic, A.; Fabek Uher, S. Bioactive compounds content and nutritional potential of different parsley parts (Petroselinum crispum Mill.). J. Cent. Eur. Agric. 2019, 20, 900–910. [Google Scholar] [CrossRef] [Green Version]
- Agyare, C.; Appiah, T.; Boakye, Y.D.; Apenteng, J.A. Petroselinum crispum: A review. In Medicinal Spices and Vegetables from Africa, Therapeutic Potential Against Metabolic, Inflammatory, Infectious and Systemic Diseases, 1st ed.; Kuete, V., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 527–547. [Google Scholar]
- Liberal, A.; Fernandes, A.; Polyzos, N.; Petropoulos, S.A.; Dias, M.I.; Pinela, J.; Petrović, J.; Soković, M.; Ferreira, I.C.F.R.; Barros, L. Bioactive properties and phenolic compound profiles of turnip-rooted, plain-leafed and curly-leafed parsley cultivars. Molecules 2020, 25, 5606. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Marra, R.; Cavallo, P.; Pironti, A.; Sepe, I.; Troisi, J.; Scala, G.; Lombari, P.; Vinale, F. Trichoderma Strains and Metabolites selectively increase the production of volatile organic compounds (VOCs) in Olive trees. Metabolites 2021, 11, 213. [Google Scholar] [CrossRef]
- Dini, I.; Graziani, G.; Gaspari, A.; Fedele, F.L.; Sicari, A.; Vinale, F.; Cavallo, P.; Lorito, M.; Ritieni, A. New strategies in the cultivation of Olive trees and repercussions on the nutritional value of the Extra Virgin Olive oil. Molecules 2020, 25, 2345. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Bubici, G. Streptomyces spp. as biocontrol agents against Fusarium species. CAB Rev. 2018, 13, 50. [Google Scholar] [CrossRef]
- Sivalingam, P.; Hong, K.; Pote, J.; Prabakar, K. Extreme environment Streptomyces: Potential sources for new antibacterial and anticancer drug leads? Int. J. Microbiol. 2019, 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteca, Á.; Yagüe, P. Streptomyces Differentiation in liquid cultures as a trigger of secondary metabolism. Antibiotics 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubici, G.; Marsico, A.D.; D’Amico, M.; Amenduni, M.; Cirulli, M. Evaluation of Streptomyces spp. for the biological control of corky root of tomato and Verticillium wilt of eggplant. Appl. Soil Ecol. 2013, 72, 128–134. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi-Zarandi, M.; Bonjar, G.H.S.; Riseh, R.S.; El-Shetehy, M.; Saadoun, I.; Barka, E.A. Exploring two Streptomyces species to control Rhizoctonia solani in tomato. Agronomy 2021, 11, 1384. [Google Scholar] [CrossRef]
- Bonaldi, M.; Chen, X.; Kunova, A.; Pizzatti, C.; Saracchi, M.; Cortesi, P. Colonization of lettuce rhizosphere and roots by tagged Streptomyces. Front. Microbiol. 2015, 6, 25. [Google Scholar] [CrossRef]
- Dini, I.; Graziani, G.; Fedele, F.L.; Sicari, A.; Vinale, F.; Castaldo, L.; Ritieni, A. Effects of Trichoderma biostimulation on the phenolic profile of Extra-Virgin Olive oil and Olive oil by-products. Antioxidants 2020, 9, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barroso, F.M.; Pereira Costa Muniz, P.H.; Milan, M.D.; Silva dos Santos, W.; Cássia de Faria Ferreira, N.; Rodriguez, F.; Carvalho, D.D.C. Growth promotion of Parsley (Petroselinum crispum L.) using commercial strains of Trichoderma spp. J. Agric. Sci. 2019, 11, 493–499. [Google Scholar] [CrossRef]
- Saraylou, M.; Nadian Ghomsheh, H.; Enayatizamir, N.; Rangzan, N.; St. Clair Senn, S. Some plant growth promoting traits of Streptomyces species isolated from various crop rhizospheres with high root colonization ability of spinach (Spinacia oleracea L.). Appl. Ecol. Environ. Sci. 2021, 19, 3069–3081. [Google Scholar] [CrossRef]
- European Pharmacopoeia Commission. European Pharmacopoeia; Maisonneuve: Sainte-Ruffine, France, 1975; pp. 68–80. [Google Scholar]
- Gruszecki, R. The effect of growth stage on essential oil composition of ‘Hamburg’ parsley. Acta Hort. 2007, 826, 391–396. [Google Scholar] [CrossRef]
- Lamarti, A.; Badoc, A.; Bouriquet, R. A chemotaxonomic evaluation of Petroselinum crispum (Mill.) AW Hill (parsley) marketed in France. J. Essent. Oil Res. 1991, 3, 425–433. [Google Scholar] [CrossRef]
- Snoussi, M.; Dehmani, A.; Noumi, E.; Flamini, G.; Papetti, A. Chemical composition and antibiofilm activity of Petroselinum crispum and Ocimum basilicum essential oils against Vibrio spp. strains. Microb. Pathog. 2016, 90, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Daferera, D.; Akoumianakis, C.A.; Passam, H.C.; Polissiou, M.G. The effect of sowing date and growth stage on the essential oil composition of three types of parsley (Petroselinum crispum). J. Sci. Food Agric. 2004, 84, 1606–1610. [Google Scholar] [CrossRef]
- John, R.P.; Tyagi, R.D.; Prevost, D.; Brar, S.K.; Pouleur, S.; Surampalli, R.Y. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp. Spadzuki and Pythium arrhenomanes as a growth promoter of soybean. Crop. Prot. 2010, 29, 1452–1459. [Google Scholar] [CrossRef]
- Doni, F.; Anizan, I.; Che Radziah, C.M.Z.; Salman, A.H.; Rodzihan, M.H.; Wan Mohtar, W.Y. Enhancement of rice seed germination and vigour by Trichoderma spp. Res. J. App. Sci. Eng. Technol. 2014, 7, 4547–4552. [Google Scholar] [CrossRef]
- Pascale, A.; Vinale, F.; Manganiello, G.; Nigro, M.; Lanzuise, S.; Ruocco, M.; Marra, R.; Lombardi, N.; Woo, S.L.; Lorito, M. Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop. Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-growth promotion and biocontrol properties of three Streptomyces spp. isolates to control bacterial rice pathogens. Front. Microbiol. 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, A.F.; Clark, G.C.; Dupuy, H.P. Antimicrobial activity of some ricinoleic acid derivatives. J. Am. Oil Chem. Soc. 1960, 38, 321–324. [Google Scholar] [CrossRef]
- Abdellatief, S.A.; Galal, A.A.A.; Farouk, S.M.; Abdel-Daim, M.M. Ameliorative effect of parsley oil on cisplatin-induced hepato-cardiotoxicity: A biochemical, histopathological, and immunochemical study. Biomed. Pharmacother. 2017, 86, 482–491. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.M.; Metwally, N.H.; Ibrahim, I.A.; Abdel-Hady, H.; Abdel-Wahab, B.S.A. Antioxidant activity, total phenolic and flavonoid contents of Petroselinum crispum Mill. J. Appl. Life Sci. 2018, 19, 1–7. [Google Scholar] [CrossRef]
- Caboni, P.; Saba, M.; Oplos, C.; Aissani, N.; Maxia, A.; Menkissoglu-Spiroudi, U.; Casu, L.; Ntalli, N. Nematicidal activity of furanocoumarins from parsley against Meloidogyne spp. Pest. Magan. Sci. 2015, 71, 1099–1105. [Google Scholar] [CrossRef]
- de Menezes Epifanio, N.M.; Cavalcanti, L.R.I.; dos Santos, K.F.; Duarte, P.S.C.; Kachlicki, P.; Ozarowski, M.; Riger, C.J.; de Almeida Chaves, D.S. Chemical characterization and in vivo antioxidant activity of parsley (Petroselinum crispum) aqueous extract. Food Funct. 2020, 11, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An important class of phytochemicals. In Phytochemicals—Isolation, Characterization and Role in Human Health, 1st ed.; Venket Rao, A.V.A., Rao, L.G., Eds.; IntechOpen: London, UK, 2015. [Google Scholar]
- Murray, R.D.H.; Mendez, J.; Brown, S.A. The Natural Coumarins. Occurance, Chemistry and Biochemistry, 1st ed.; John Wiley & Sons Ltd.: New York, NY, USA, 1982. [Google Scholar]
- Hauffe, K.D.; Hahlbrock, K.; Scheel, D. Elicitor-stimulated furanocoumarin biosynthesis in cultured parsley cells: S-adenosyl-L-methionine: BergaptoI and S-adenosyl-L-methionine: Xanthotoxol O-methyltransferases. Z. Naturforsch C 1986, 41, 228–239. [Google Scholar] [CrossRef]
- Harborne, J.B. Flavonol O-glycosides. In The Handbook of Natural Flavonoids, 1st ed.; Harborne, J.B., Baxter, H., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 1999; Volume 1, pp. 297–548. [Google Scholar]
- Uitterhaegen, E.; Sampaio, K.A.; Delbeke, E.I.P.; De Greyt, W.; Cerny, M.; Evon, P.; Merah, O.; Talou, T.; Stevens, C.V. Characterization of French coriander oil as source of petroselinic acid. Molecules. 2016, 21, 1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placek, L.L. Review on petroselinic acid and its derivatives. J. Am. Oil Chem. Soc. 1963, 40, 319–329. [Google Scholar] [CrossRef]
- Yeganehpoor, F.; Zehtab-Salmasi, S.; Ghassemi-Golezani, K.; Shafagh-Kolvanagh, J.; Dastborhan, S. The effect of water stress, salicylic acid and bio-fertilizer on quality of leaf and seed essential oil, and oil components of coriander. Net. J. Agric. Sci. 2017, 5, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Donato, P.; Dugo, P.; Mondello, L. Chapter 8: Separation of Lipids. In Liquid Cromathography, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signaling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Narita, Y.; Inouye, K. Chlorogenic acids from coffee. In Coffee in Health and Disease Prevention, 1st ed.; Preedy, V.R., Ed.; Elsevier Science & Technology: Amsterdam, The Netherlands, 2014; Volume 1, pp. 189–197. [Google Scholar]
- Aissani, N.; Albouchi, F.; Sebai, H. Anticancer Effect in human glioblastoma and antioxidant activity of Petroselinum crispum L. methanol extract. Nutr. Cancer 2020, 29, 1–9. [Google Scholar] [CrossRef]
- Jain, A.; Singh, A.; Chaudhary, A.; Singh, S.; Singh, H.B. Modulation of nutritional and antioxidant potential of seeds and pericarp of pea pods treated with microbial consortium. Food Res. Int. 2014, 64, 275–282. [Google Scholar] [CrossRef]
- Vinci, G.; Cozzolino, V.; Mazzei, P.; Monda, H.; Spaccini, R.; Piccolo, A. An alternative to mineral phosphorus fertilizers: The combined effects of Trichoderma harzianum and compost on Zea mays, as revealed by 1H NMR and GC-MS metabolomics. PLoS ONE. 2018, 13, e0209664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gębarowska, E.; Pytlarz-Kozicka, M.; Nöfer, J.; Łyczko, J.; Adamski, M.; Szumny, A. The effect of Trichoderma spp. on the composition of volatile secondary metabolites and biometric parameters of coriander (Coriandrum sativum L.). J. Food Qual. 2019, 2019, 5687032. [Google Scholar] [CrossRef] [Green Version]
- Coppola, M.; Cascone, P.; Chiusano, M.L.; Colantuono, C.; Lorito, M.; Pennacchio, F.; Rao, R.; Woo, S.L.; Guerrieri, E.; Digilio, M.C. Trichoderma harzianum enhances tomato indirect defense against aphids. Insect Sci. 2017, 24, 1025–1033. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | Compound | RT (min) | Experimental Mass (Da) | Theoretical Mass (Da) | Molecular Formula |
|---|---|---|---|---|---|
| 1 | Isorhamnetin 3,7-di-O-beta-glucopyranoside | 3.848667 | 640.1663 | 640.1639 | C28H32O17 |
| 2 | Xanthotoxin/Bergapten * | 6.000033 | 216.0427 | 216.0422 | C12H8O4 |
| 3 | Capsanthone | 6.425001 | 582.4098 | 582.4072 | C40H54O3 |
| 4 | Petroselinic acid | 6.530999 | 282.257 | 282.2558 | C18H34O2 |
| 5 | Bergaptol/Xanthotoxol * | 7.286808 | 202.0272 | 202.0266 | C11H6O4 |
| 6 | Piperochromanoic acid | 7.955463 | 356.198 | 356.1988 | C22H28O4 |
| - | Quinic acid ** | 0.9503336 | 192.0642 | 192.0633 | C7H12O6 |
| Compound | Regulation | ||||
|---|---|---|---|---|---|
| S vs. C | T vs. C | S + T vs. C | S + T vs. S | S + T vs. T | |
| Xanthotoxin/Bergapten * | ↓ | ↑ | ↓ | ↓ | ↓ |
| Piperochromanoic acid | ↑ | ↑ | ↓ | ↓ | ↓ |
| Xanthotoxol/Bergaptol * | ↓ | ↓ | ↓ | ↑ | ↓ |
| Isorhamnetin 3,7-di-O-beta-glucopyranoside | ↓ | ↓ | ↓ | ↓ | ↓ |
| Petroselinic acid | ↑ | ↓ | ↓ | ↓ | ↑ |
| Capsanthone | ↓ | ↓ | ↓ | ↑ | ↑ |
| Quinic acid | ↑ | ↑ | ↑ | ↑ | ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staropoli, A.; Vassetti, A.; Salvatore, M.M.; Andolfi, A.; Prigigallo, M.I.; Bubici, G.; Scagliola, M.; Salerno, P.; Vinale, F. Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms. Horticulturae 2021, 7, 281. https://doi.org/10.3390/horticulturae7090281
Staropoli A, Vassetti A, Salvatore MM, Andolfi A, Prigigallo MI, Bubici G, Scagliola M, Salerno P, Vinale F. Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms. Horticulturae. 2021; 7(9):281. https://doi.org/10.3390/horticulturae7090281
Chicago/Turabian StyleStaropoli, Alessia, Anastasia Vassetti, Maria Michela Salvatore, Anna Andolfi, Maria Isabella Prigigallo, Giovanni Bubici, Marina Scagliola, Pasquale Salerno, and Francesco Vinale. 2021. "Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms" Horticulturae 7, no. 9: 281. https://doi.org/10.3390/horticulturae7090281
APA StyleStaropoli, A., Vassetti, A., Salvatore, M. M., Andolfi, A., Prigigallo, M. I., Bubici, G., Scagliola, M., Salerno, P., & Vinale, F. (2021). Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms. Horticulturae, 7(9), 281. https://doi.org/10.3390/horticulturae7090281