
Changes in Metal Distribution, Vegetative Growth, Reactive Oxygen and Nutrient Absorption of Tagetes patula under Soil Cadmium Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Plant Material
2.3. Experimental Design
2.4. Measurement of Morphological Traits
2.5. Measurement of Inflorescence and Flowering Dynamics
2.6. Measurement of Cd, Total N, Total P and Total K Content
2.7. Measurement of ROS and MDA
2.8. Statistics
3. Results
3.1. Effect of Cd Stress on Morphological Traits
3.2. Effect of Cd Stress on Flowering Traits
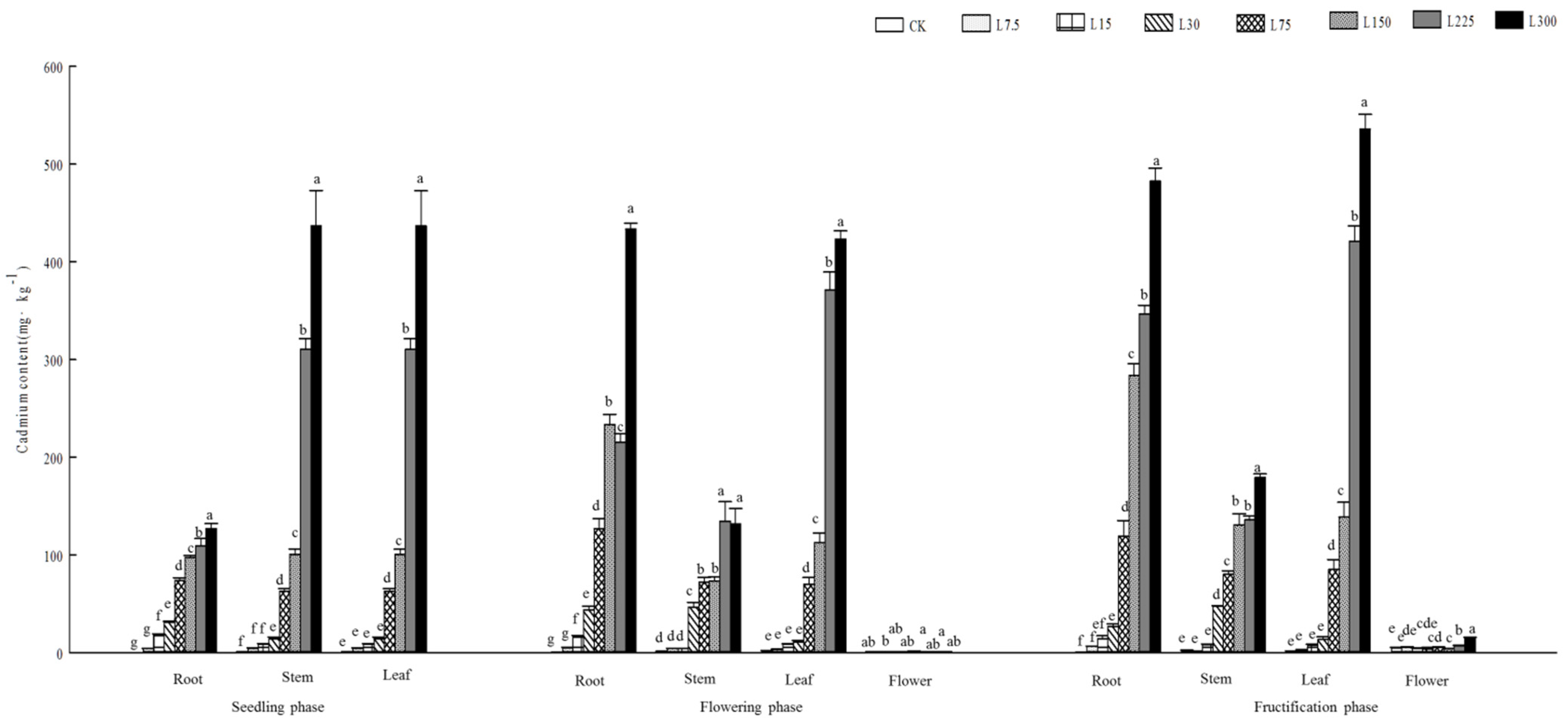
3.3. Cd Distribution with Plant
3.4. Effect of Cd Stress on Nutrient Distribution
3.5. Active Oxygen Injury Caused by Cd Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CK | control treatment |
| AFmax | the maximum accumulative flower number per plant |
| Dfirst | the day the first flower appeared |
| Dlast | the day the last flower appeared |
| RSD | relative standard deviation |
| ROS | reactive oxygen species |
| MDA | malondialdehyde |
| LSD | least significant difference |
| GLMs | generalized linear models |
References
- Eid, E.M.; Galal, T.M.; El-Bebany, A.F. Prediction models for monitoring heavy-metal accumulation by wheat (Triticum aestivum L.) plants grown in sewage sludge amended soil. Int. J. Phytoremediat. 2020, 22, 1000–1008. [Google Scholar] [CrossRef]
- NBS (National Bureau of Statistics). 2020. Available online: http://data.stats.gov.cn/easyquery.htm?cn¼C01 (accessed on 1 June 2021).
- Zou, M.; Zhou, S.; Zhou, Y.; Jia, Z.; Guo, T.; Wang, J. Cadmium pollution of soil-rice ecosystems in rice cultivation dominated regions in China: A review. Environ. Pollut. 2021, 280, 116965. [Google Scholar] [CrossRef]
- Wang, L.; Cui, X.; Cheng, H.; Chen, F.; Wang, J.; Zhao, X.; Lin, C.; Pu, X. A review of soil cadmium contamination in China including a health risk assessment. Environ. Sci. Pollut. Res. 2015, 22, 16441–16452. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, S.; Wan, X.; Yang, P. Research on the environmental property and potential ecological risk of the soil and sediment around the typical enterprises relating to heavy mental in the southwest. Environ. Monit. China 2014, 30, 13–19. [Google Scholar] [CrossRef]
- Pająk, M.; Błońska, E.; Szostak, M.; Gąsiorek, M.; Pietrzykowski, M.; Urban, O.; Derbis, P. Restoration of Vegetation in Relation to Soil Properties of Spoil Heap Heavily Contaminated with Heavy Metals. Water Air Soil Pollut. 2018, 229, 392. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef]
- Fu, S.; Wei, C.; Xiao, Y.; Li, L.; Wu, D. Heavy metals uptake and transport by native wild plants: Implications for phytoremediation and restoration. Environ. Earth Sci. 2019, 78, 103. [Google Scholar] [CrossRef]
- Li, F.; Ma, D. Species composition and floristic characteristics of hyperaccumulators in Sichuan province. J. Sichuan Norm. Univ. 2020, 43, 263–269. [Google Scholar] [CrossRef]
- Nakbanpote, W.; Meesungnoen, O.; Prasad, M.N.V. Potential of Ornamental Plants for Phytoremediation of Heavy Metals and Income Generation. In Bioremediation and Bioeconomy; Elsevier, Inc.: Philadelphia, PA, USA, 2016; pp. 179–217. [Google Scholar] [CrossRef]
- Ye, X.; Hu, H.; Li, H.; Xiong, Q.; Gao, H. Combined nitrogen fertilizer and wheat straw increases the cadmium phytoextraction efficiency of Tagetes patula. Ecotoxicol. Environ. Saf. 2019, 170, 210–217. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, W.; He, X. Evaluation of hyperaccumulation potentials to cadmium (Cd) in six ornamental species (compositae). Int. J. Phytoremediat. 2018, 20, 1464–1469. [Google Scholar] [CrossRef]
- Sun, R.; Sun, Q.; Wang, R.; Cao, L. Cadmium accumulation and main rhizosphere characteristics of seven French marigold (Tagetes patula L.) cultivars. Int. J. Phytoremediation 2018, 20, 1171–1178. [Google Scholar] [CrossRef]
- Tőzsér, D.; Magura, T.; Simon, E. Heavy metal uptake by plant parts of willow species: A meta-analysis. J. Hazard. Mater. 2017, 336, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Mondal, M.K.; Mishra, G.; Kumar, P. Adsorption of Cadmium (II) and Chromium (VI) from Aqueous Solution by Waste Marigold Flowers. J. Sustain. Dev. Energy Water Environ. Syst. 2015, 3, 405–415. [Google Scholar] [CrossRef]
- Liu, L.-Z.; Gong, Z.-Q.; Zhang, Y.-L.; Li, P.-J. Growth, Cadmium Accumulation and Physiology of Marigold (Tagetes erecta L.) as Affected by Arbuscular Mycorrhizal Fungi. Pedosphere 2011, 21, 319–327. [Google Scholar] [CrossRef]
- Tabrizi, L.; Lakzaei, M.; Motesharezadeh, B. The yield potential and growth responses of licorice (Glycyrrhiza glabra L.) to mycorrhization under Pb and Cd stress. Int. J. Phytoremediat. 2021, 23, 316–327. [Google Scholar] [CrossRef]
- Zhou, Y.-M.; Long, S.-S.; Li, B.-Y.; Huang, Y.-Y.; Li, Y.-J.; Yu, J.-Y.; Du, H.-H.; Khan, S.; Lei, M. Enrichment of cadmium in rice (Oryza sativa L.) grown under different exogenous pollution sources. Environ. Sci. Pollut. Res. 2020, 27, 44249–44256. [Google Scholar] [CrossRef]
- Omer, C.; Dursun, K. Flower yields of pot marigold (Calendula officinalis L.) plants as effected by flowering durations and number of harvests. J. Med. Plants Stud. 2018, 6, 159–161. [Google Scholar]
- Thakur, P.; Dilta, B.; Gupta, Y.; Kumar, P.; Mehta, D. Effect of Planting Dates, Mulching and Application of GA3 on growth and flower yield of Marigold (Tagetes Erecta L.) cv. ‘Pusa Narangi Gainda’. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 3028–3039. [Google Scholar] [CrossRef]
- Balijagić, J.; Crnobarac, J.; Jovancević, M.; Mujanovic, I. The effect of variety and plant den-sity on marigold flower yield (Calendula officinalis L.) in agro-ecological conditions of Bijelo Polje. J. Agric. For. 2018, 64, 113–120. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, P.; Hu, Y.; Liu, Y.; Feng, S.; Guo, D.; Dang, X. Immobilization of cadmium in soil and improved iron concentration and grain yields of maize (Zea mays L.) by chelated iron amendments. Environ. Sci. Pollut. Res. 2021, 28, 53161–53170. [Google Scholar] [CrossRef]
- Türker, A.; Acar, O. Investigation of cadmium and lead contents of boron minerals and boron waste by electrothermal atomic absorption spectrometer. J. Iran. Chem. Soc. 2021, 18, 1607–1612. [Google Scholar] [CrossRef]
- Chen, L.; Hu, X.; Yang, W.; Xu, Z.; Zhang, D.; Gao, S. The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in Populus cathayana. Ecotoxicol. Environ. Saf. 2015, 113, 460–468. [Google Scholar] [CrossRef]
- Fang, Z.; Wan, H.; Xu, Y.; Liu, Q.; Liang, J.; Shi, X.; Chen, F. Indole-acyl esters improve the effect of nitrogen and phosphorous fertilization by mitigating the phytotoxicity and concentrations of cadmium and lead in Jatropha curcas L. in contaminated soils. Arch. Biol. Sci. 2019, 71, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Ondrasek, G.; Rengel, Z.; Romic, D. Humic acids decrease uptake and distribution of trace metals, but not the growth of radish exposed to cadmium toxicity. Ecotoxicol. Environ. Saf. 2018, 151, 55–61. [Google Scholar] [CrossRef]
- Dharma-Wardana, M.W.C. Fertilizer usage and cadmium in soils, crops and food. Environ. Geochem. Heal. 2018, 40, 2739–2759. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Li, J.; Chen, F.; Liao, M.; Tang, Y.; Liang, N.; Xia, H.; Lai, Y.; Wang, X.; Chen, C.; et al. Effects of melatonin on the growth and cadmium characteristics of Cyphomandra betacea seedlings. Environ. Monit. Assess. 2018, 190, 119. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Mahmood, A.; Zia-Ur-Rehman, M.; Ibrahim, M.; Arshad, M.; Qayyum, M.F. Biochar application increased the growth and yield and reduced cadmium in drought stressed wheat grown in an aged contaminated soil. Ecotoxicol. Environ. Saf. 2018, 148, 825–833. [Google Scholar] [CrossRef]
- Zemanová, V.; Pavlík, M.; Pavlíková, D.; Hnilička, F.; Vondráčková, S. Responses to Cd Stress in Two Noccaea Species (Noccaea praecox and Noccaea caerulescens) Originating from Two Contaminated Sites in Mežica, Slovenia and Redlschlag, Austria. Arch. Environ. Contam. Toxicol. 2015, 70, 464–474. [Google Scholar] [CrossRef]
- Thongchai, A.; Meeinkuirt, W.; Taeprayoon, P.; Pichtel, J. Soil amendments for cadmium phytostabilization by five marigold cultivars. Environ. Sci. Pollut. Res. 2019, 26, 8737–8747. [Google Scholar] [CrossRef]
- Rasheed, R.; Ashraf, M.A.; Kamran, S.; Iqbal, M.; Hussain, I. Menadione sodium bisulphite mediated growth, secondary metabolism, nutrient uptake and oxidative defense in okra (Abelmoschus esculentus Moench) under cadmium stress. J. Hazard. Mater. 2018, 360, 604–614. [Google Scholar] [CrossRef]
- Liu, C.-H.; Chao, Y.-Y.; Kao, C.H. Effect of potassium deficiency on antioxidant status and cadmium toxicity in rice seedlings. Bot. Stud. 2013, 54, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.-Q.; Zhu, Y.-G.; Li, H.-Y.; Smith, S.E.; Smith, F. Effects of forms and rates of potassium fertilizers on cadmium uptake by two cultivars of spring wheat (Triticum aestivum L.). Environ. Int. 2004, 29, 973–978. [Google Scholar] [CrossRef]
- Ben Ghnaya, A.; Charles, G.; Hourmant, A.; Ben Hamida, J.; Branchard, M. Physiological behaviour of four rapeseed cultivar (Brassica napus L.) submitted to metal stress. Comptes Rendus Biol. 2009, 332, 363–370. [Google Scholar] [CrossRef]
- Wei, S.; Zhou, Q.; Koval, P.V. Flowering stage characteristics of cadmium hyperaccumulator Solanum nigrum L. and their significance to phytoremediation. Sci. Total Environ. 2006, 369, 441–446. [Google Scholar] [CrossRef]
- Moradi, R.; Pourghasemian, N.; Naghizadeh, M. Effect of beeswax waste biochar on growth, physiology and cadmium uptake in saffron. J. Clean. Prod. 2019, 229, 1251–1261. [Google Scholar] [CrossRef]
- Hladun, K.R.; Parker, D.R.; Trumble, J.T. Cadmium, Copper, and Lead Accumulation and Bioconcentration in the Vegetative and Reproductive Organs of Raphanus sativus: Implications for Plant Performance and Pollination. J. Chem. Ecol. 2015, 41, 386–395. [Google Scholar] [CrossRef]
- Marquez, J.; Pourret, O.; Faucon, M.-P.; Weber, S.; Hoàng, T.; Martinez, R. Effect of Cadmium, Copper and Lead on the Growth of Rice in the Coal Mining Region of Quang Ninh, Cam-Pha (Vietnam). Sustainability 2018, 10, 1758. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, C.C.; Ricketts, T.H.; Koh, I.; Smith, H.G.; Lonsdorf, E.V.; Olsson, O. Flowering resources distract pollinators from crops: Model predictions from landscape simulations. J. Appl. Ecol. 2019, 56, 618–628. [Google Scholar] [CrossRef]
- Prins, C.N.; Hantzis, L.J.; Quinn, C.F.; Pilon-Smits, E.A.H. Effects of selenium accumulation on reproductive functions in Brassica juncea and Stanleya pinnata. J. Exp. Bot. 2011, 62, 5633–5640. [Google Scholar] [CrossRef] [Green Version]
- Korshin, G.V.; Ferguson, J.F.; Lancaster, A.N. Influence of natural organic matter on the morphology of corroding lead surfaces and behavior of lead-containing particles. Water Res. 2005, 39, 811–818. [Google Scholar] [CrossRef]
- Xun, E.; Zhang, Y.; Zhao, J.; Guo, J. Translocation of heavy metals from soils into floral organs and rewards of Cucurbita pepo: Implica-tions for plant reproductive fitness. Ecotoxicol. Environ. Saf. 2017, 145, 235–243. [Google Scholar] [CrossRef]
- Meeinkuirt, W.; Kruatrachue, M.; Pichtel, J.; Phusantisampan, T.; Saengwilai, P. Influence of organic amendments on phytostabilization of Cd-contaminated soil by Eucalyptus camaldulensis. Science 2016, 42, 83. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.U.; Tang, Z.; Zheng, T.; Asghar, M.A.; Zeng, R.; Su, Y.; Ei, H.H.; Liang, Y.; Zhang, Y.; Ye, X.; et al. Cross-Talk between Cadmium and Selenium at Elevated Cadmium Stress Determines the Fate of Selenium Uptake in Rice. Biomolecules 2019, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.; Huang, S.; Ashraf, U.; Mo, Z.; Duan, M.; Pan, S.; Tang, X. Ultrasonic seed treatment improved cadmium (Cd) tolerance in Brassica napus L. Ecotoxicol. Environ. Saf. 2019, 185, 109659. [Google Scholar] [CrossRef]
- Xie, L.; Hao, P.; Cheng, Y.; Ahmed, I.M.; Cao, F. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 162, 71–76. [Google Scholar] [CrossRef]
- Bhatti, S.S.; Kumar, V.; Sambyal, V.; Singh, J.; Nagpal, A.K. Comparative analysis of tissue compartmentalized heavy metal uptake by common forage crop: A field experiment. Catena 2018, 160, 185–193. [Google Scholar] [CrossRef]
- Qin, Q.; Li, X.; Wu, H.; Zhang, Y.; Feng, Q.; Tai, P. Characterization of cadmium (108Cd) distribution and accumulation in Tagetes erecta L. seedlings: Effect of split-root and of remove-xylem/phloem. Chemosphere 2013, 93, 2284–2288. [Google Scholar] [CrossRef]
- Zaheer, M.M.; Yasin, N.A.; Ahmad, S.R.; Khan, W.U.; Ahmad, A.; Ali, A.; Rehman, S.U. Amelioration of cadmium stress in gladiolus (Gladiolus grandiflora L.) by application of potassium and silicon. J. Plant Nutr. 2017, 41, 461–476. [Google Scholar] [CrossRef]
- Sarma, H. Metal Hyperaccumulation in Plants: A Review Focusing on Phytoremediation Technology. J. Environ. Sci. Technol. 2011, 4, 118–138. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wei, H.; Zhou, C.; Gu, Y.; Li, R.; Chen, H.; Ma, W. Estimating cadmium concentration in the edible part of Capsicum annuum using hyperspectral models. Environ. Monit. Assess. 2017, 189, 548. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Matter (%) | Macronutrients (mg·kg−1) | Micronutrients (mg·kg−1) | EC (mS·cm−1) | pH | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | Cd | Zn | Fe | B | Mo | Mn | Cu | |||
| 2.38 ± 0.25 | 79.62 ± 1.49 | 161.48 ± 11.74 | 261.47 ± 18.91 | 212.6 ± 16.55 | 19.06 ± 2.51 | 1.84 ± 0.49 | 0.85 ± 0.12 | 0.81 ± 0.18 | 8.12 ± 1.22 | 0.33 ± 0.09 | 1.45 ± 0.33 | 2.32 ± 0.51 | 1.38 ± 0.26 | 2.51 ± 0.73 | 6.0 ± 0.41 |
| Cd Treatment | Fmax (Flower/Plant) | Dfirst (d) | Dlast (d) | FD (d) | Mathematic Model Parameter | Test Coefficient | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | x0 | F | p | AJR2 | n | |||||
| CK | 20.52 | 44.63 | 108.57 | 63.94 | 20.52 | 11.65 | 76.6 | 526.14 | <0.0001 | 0.93 | 80 |
| L7.5 | 18.76 | 46.22 | 106.18 | 59.96 | 18.76 | 11.13 | 76.2 | 229.52 | <0.0001 | 0.8526 | 80 |
| L15 | 16.02 | 44.8 | 100.36 | 55.56 | 16.02 | 11.01 | 72.58 | 164.61 | <0.0001 | 0.8055 | 80 |
| L30 | 18.48 | 45.82 | 109.04 | 63.22 | 18.48 | 12.38 | 77.43 | 385.27 | <0.0001 | 0.9068 | 80 |
| L70 | 17.16 | 46.12 | 101.7 | 55.58 | 17.16 | 10.42 | 73.91 | 500.89 | <0.0001 | 0.9268 | 80 |
| L150 | 17.15 | 45.11 | 102.07 | 56.97 | 17.15 | 10.89 | 73.59 | 215.64 | <0.0001 | 0.8446 | 80 |
| L225 | 15.09 | 46.8 | 108.56 | 61.77 | 15.09 | 14.77 | 77.68 | 35.09 | <0.0001 | 0.4632 | 80 |
| L300 | 3.99 | / | / | / | 3.99 | 8.45 | 64.2 | 3.08 | 0.0613 | 0.1183 | 32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.; Wang, Y.; Chen, H.; Deng, F.; Liu, Y.; Cao, G. Changes in Metal Distribution, Vegetative Growth, Reactive Oxygen and Nutrient Absorption of Tagetes patula under Soil Cadmium Stress. Horticulturae 2022, 8, 69. https://doi.org/10.3390/horticulturae8010069
Shi H, Wang Y, Chen H, Deng F, Liu Y, Cao G. Changes in Metal Distribution, Vegetative Growth, Reactive Oxygen and Nutrient Absorption of Tagetes patula under Soil Cadmium Stress. Horticulturae. 2022; 8(1):69. https://doi.org/10.3390/horticulturae8010069
Chicago/Turabian StyleShi, Hongzhou, Yangmei Wang, Hong Chen, Fuxiang Deng, Yongan Liu, and Guoxing Cao. 2022. "Changes in Metal Distribution, Vegetative Growth, Reactive Oxygen and Nutrient Absorption of Tagetes patula under Soil Cadmium Stress" Horticulturae 8, no. 1: 69. https://doi.org/10.3390/horticulturae8010069
APA StyleShi, H., Wang, Y., Chen, H., Deng, F., Liu, Y., & Cao, G. (2022). Changes in Metal Distribution, Vegetative Growth, Reactive Oxygen and Nutrient Absorption of Tagetes patula under Soil Cadmium Stress. Horticulturae, 8(1), 69. https://doi.org/10.3390/horticulturae8010069