
Characterisation of Breadfruit (Artocarpus altilis) Plants Growing on Lakoocha (A. lakoocha) Rootstocks


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Comparison of Breadfruit Trees on Different Rootstocks
2.3. Measurement of Leaf Chlorophyll Content
2.4. Non-Structural Carbohydrate Analysis
2.5. Statistical Analyses
3. Results
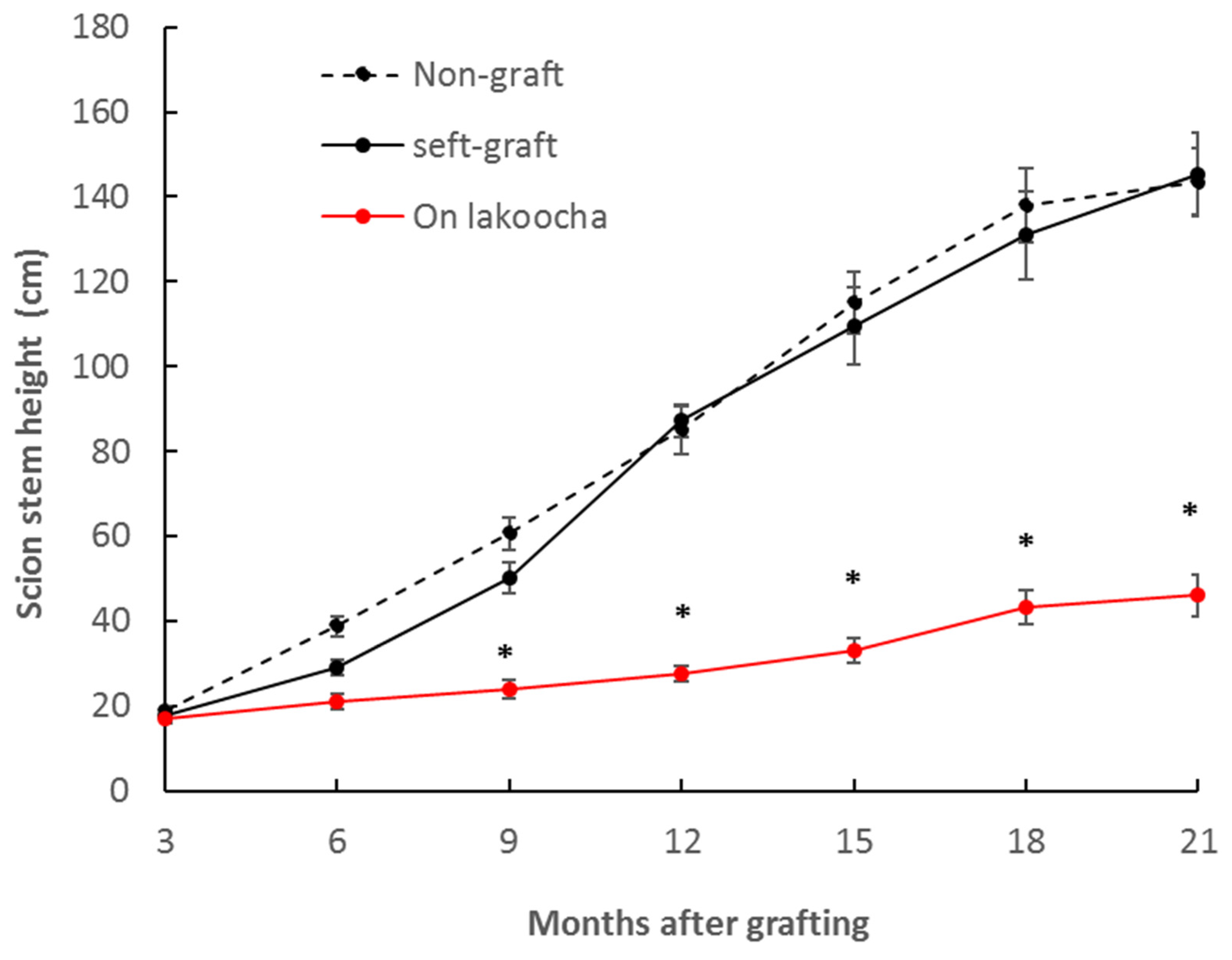
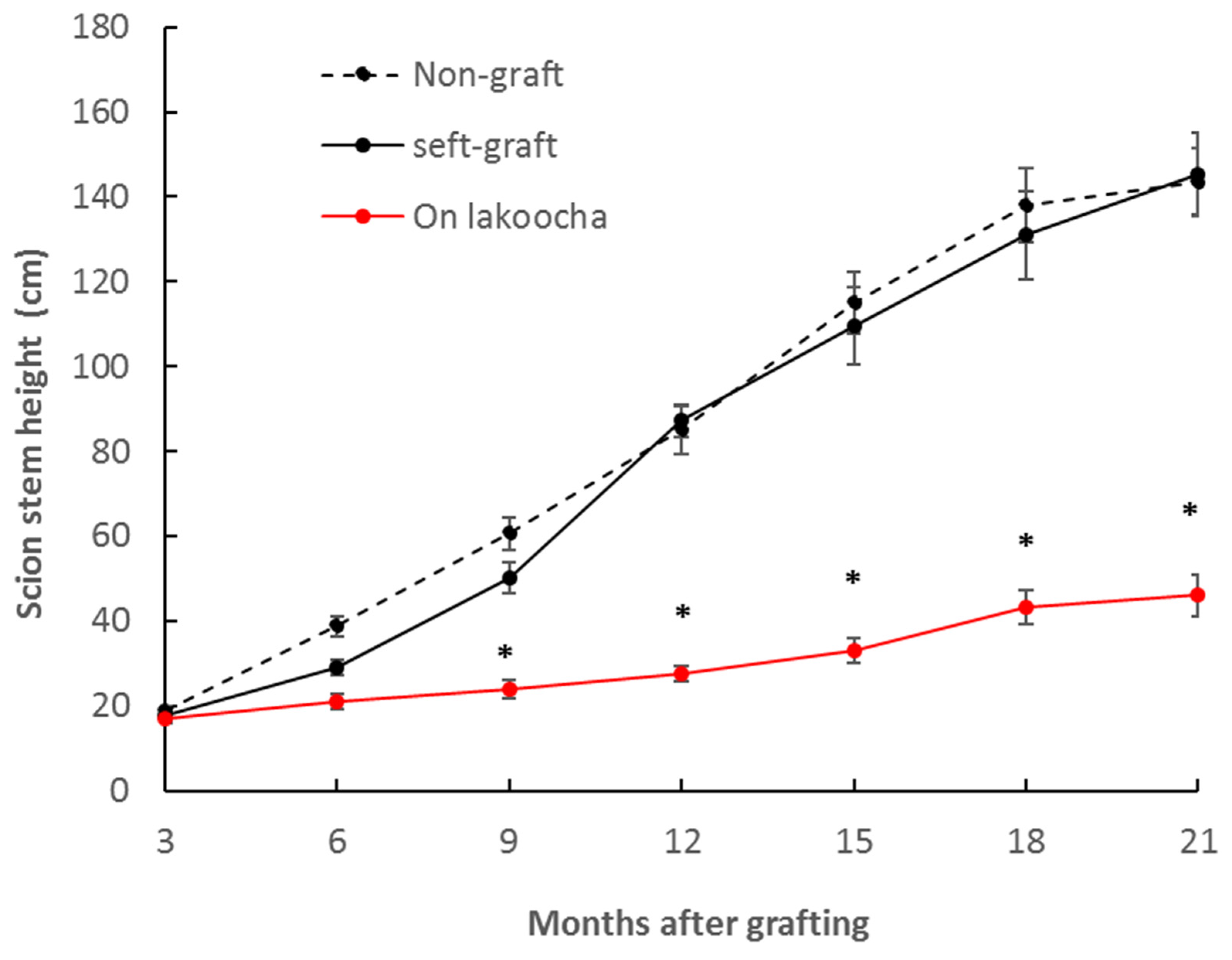
3.1. Growth Habit of Breadfruit Plants on Different Rootstocks
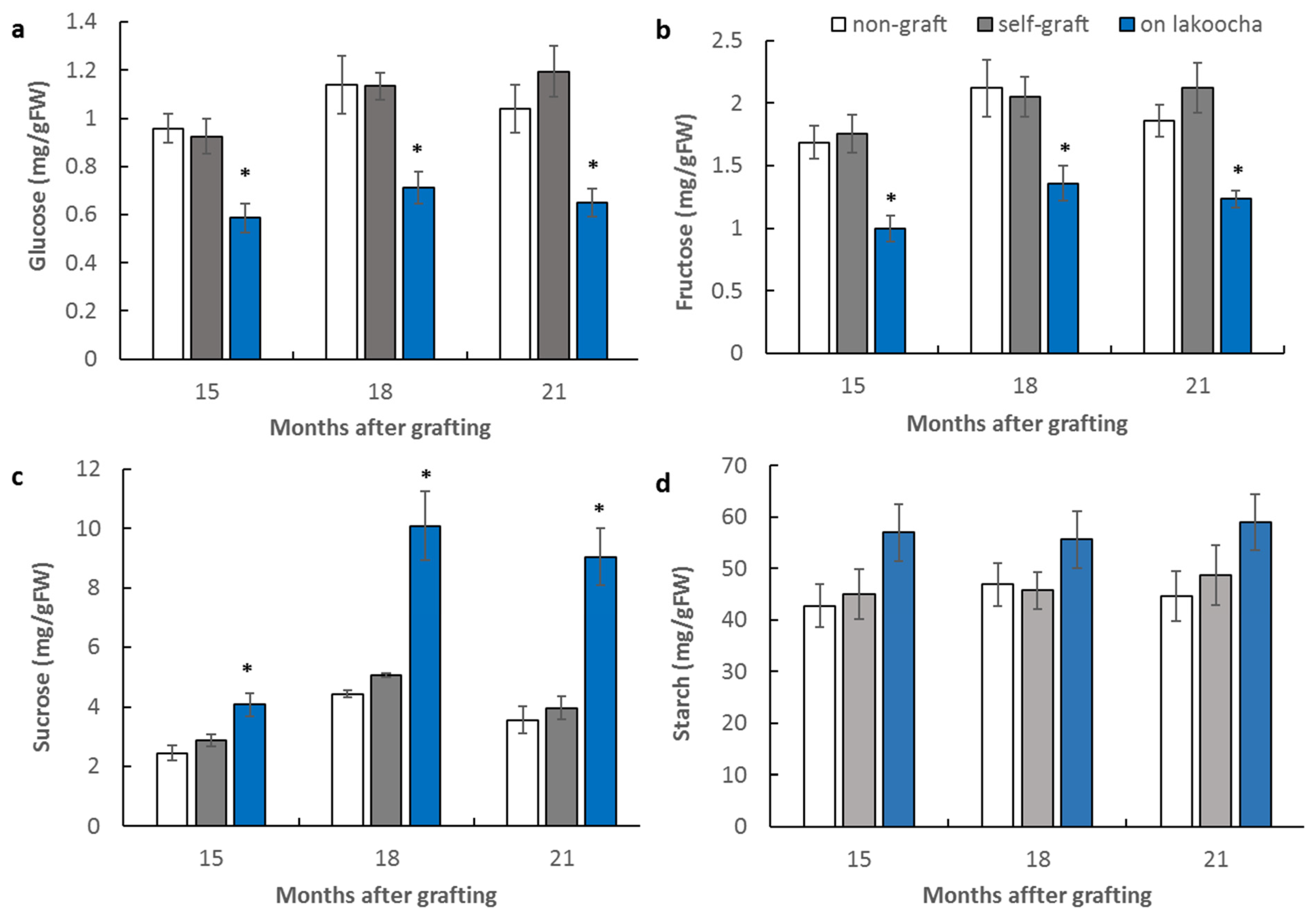
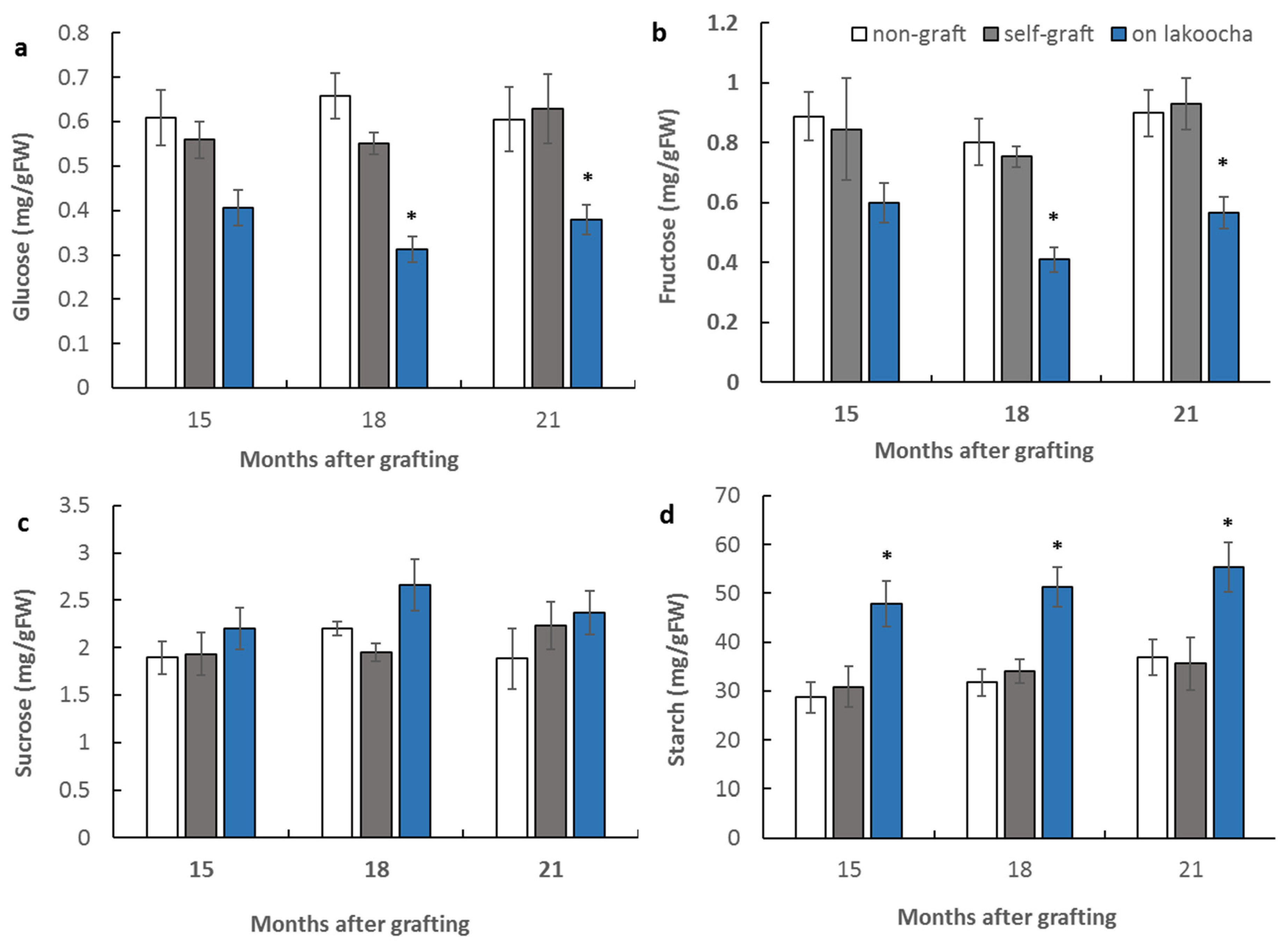
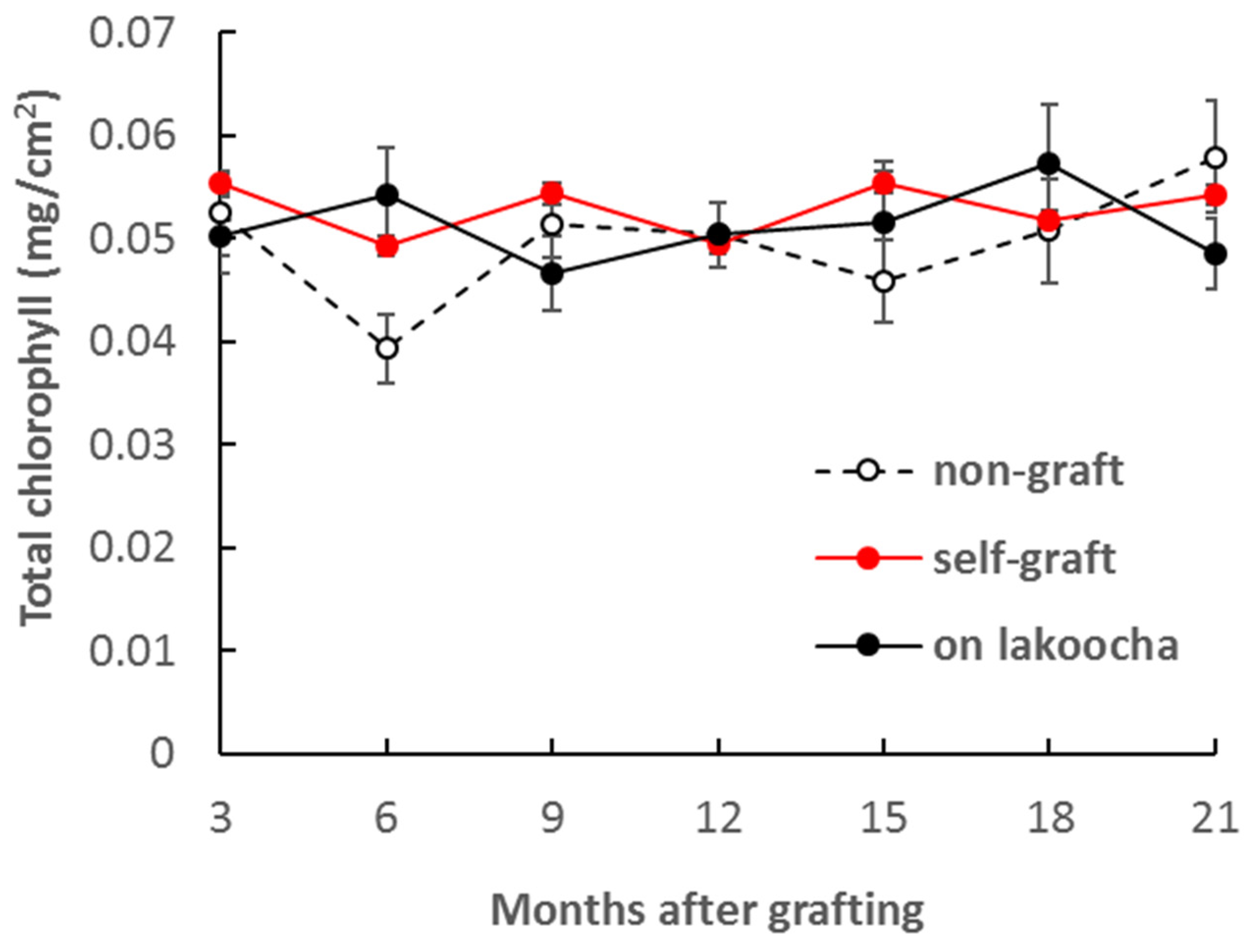
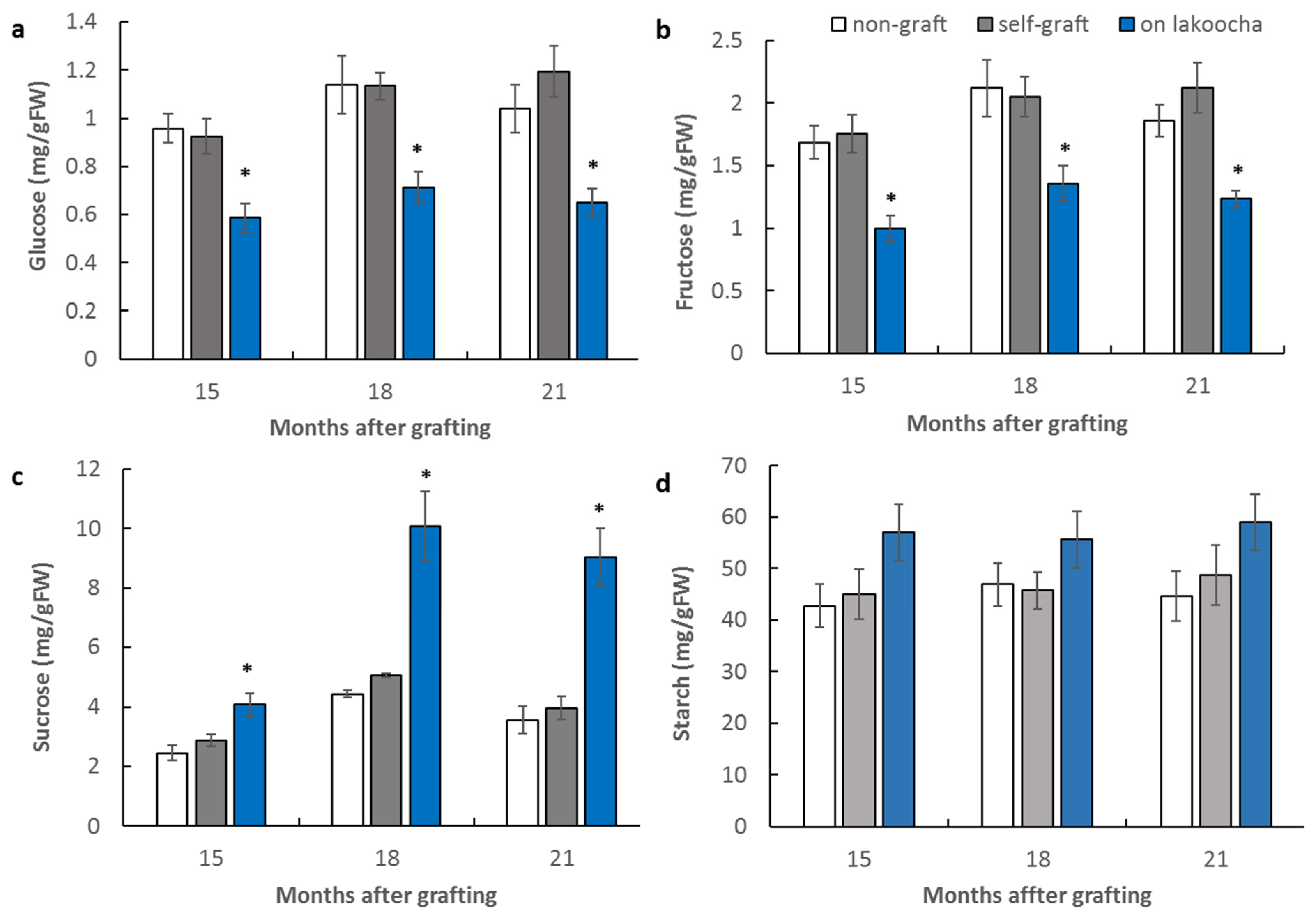
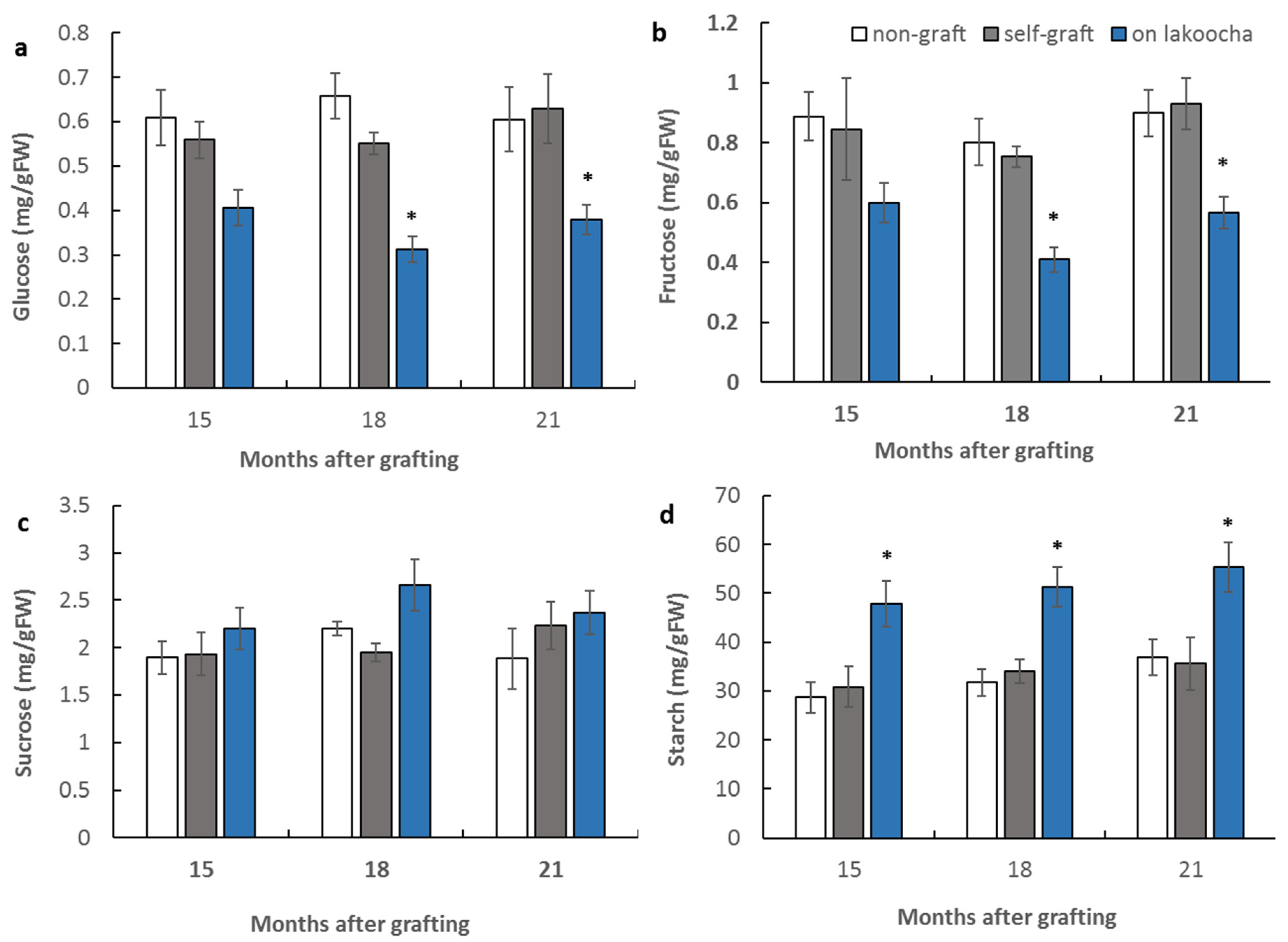
3.2. Effect of Rootstocks on Non-Structural Carbohydrate Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kehinde, A.Z.; Erland, L.A.E.; Liu, Y.; Ragone, D.; Jones, A.M.P.; Murch, S.J. South Pacific cultivars of breadfruit (Artocarpus altilis (Parkinson) Fosberg and A. mariannensis Trécul) and their hybrids (A. altilis × A. mariannensis) have unique dietary starch, protein and fiber. J. Food Compos. Anal. 2022, 105, 104228. [Google Scholar] [CrossRef]
- Roberts-Nkrumah, L.B.; Legall, G. Breadfruit (Artocarpus altilis, Moraceae) and Chataigne (A. camansi) for Food Security and Income Generation: The Case of Trinidad and Tobago. Econ. Bot. 2013, 67, 324–334. [Google Scholar] [CrossRef]
- Langston, B.J.; Lincoln, N.K. The Role of Breadfruit in Biocultural Restoration and Sustainability in Hawai‘i. Sustainability 2018, 10, 3965. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ragone, D.; Murch, S.J. Breadfruit (Artocarpus altilis): A source of high-quality protein for food security and novel food products. Amino Acids 2015, 47, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Ragone, D. Breadfruit, Artocarpus Altilis (Parkinson) Fosberg-Promoting the Conservation and Use of Underutilized and Neglected Crops, 10; Leibniz Institute of Plant Genetics and Crop Plant Research (IPK): Gatersleben, Germany, 1997; p. 77. [Google Scholar]
- Zhou, Y.; Taylor, M.B.; Underhill, S.J.R. Dwarfing of breadfruit (Artocarpus altilis) trees: Opportunities and challenges. Am. J. Exp. Agric. 2014, 4, 1743–1763. [Google Scholar] [CrossRef]
- Daley, O.; Gloster, M.; Roberts-Nkrumah, L.B. Assessment and characterization of damage by hurricane Tomas to major tree crops with special emphasis on breadfruit (artocarpus altilis) and breadnut (artocarpus camansi) in St. Lucia and St. Vincent and the Grenadines. Proc. Caribb. Food Crop Soc. 2012, 48, 124–131. [Google Scholar]
- Daley, O.O.; Roberts-Nkrumah, L.B.; Gloster, M.C.; Legall, G. Impacts of hurricanes on fruit tree crops in the Caribbean with emphasis on hurricane Tomas on breadfruit (Artocarpus altilis) and breadnut (Artocarpus camansi) in St Lucia and St Vincent and the Grenadines. Int. J. Environ. Sci. Nat. Res. 2020, 25, 556167. [Google Scholar] [CrossRef]
- Roberts-Nkrumah, L.B. Breadfruit and Breadnut Orchard Establishment and Management, a Manual for Commercial Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; p. 48. [Google Scholar]
- Jones, A.M.P.; Murch, S.J.; Wiseman, J.; Ragone, D. Morphological diversity in breadfruit (Artocarpus, Moraceae): Insights into domestication, conservation, and cultivar identification. Genet. Resour. Crop Evol. 2013, 60, 175–192. [Google Scholar] [CrossRef]
- Daley, O.O.; Roberts-Nkrumah, L.B.; Alleyne, A.T. Morphological diversity of breadfruit [Artocarpus altilis (Parkinson) Fosberg] in the Caribbean. Sci. Hortic. 2020, 266, 109278. [Google Scholar] [CrossRef]
- Roberts-Nkrumah, L.B. Breadnut and Breadfruit Propagation, a Manual for Commercial Propagation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; p. 18. [Google Scholar]
- Foster, T.M.; McAtee, P.A.; Waite, C.N.; Boldingh, H.L.; McGhie, T.K. Apple dwarfing rootstocks exhibit an imbalance in carbohydrate allocation and reduced cell growth and metabolism. Hortic. Res. 2017, 4, 17009. [Google Scholar] [CrossRef] [Green Version]
- Bruckner, C.H.; DeJong, T.M. Proposed pre-selection method for identification of dwarfing peach rootstocks based on rapid shoot xylem vessel analysis. Sci. Hortic. 2014, 165, 404–409. [Google Scholar] [CrossRef]
- Olmstead, M.A.; Lang, N.S.; Lang, G.A. Carbohydrate profiles in the graft union of young sweet cherry trees grown on dwarfing and vigorous rootstocks. Sci. Hortic. 2010, 124, 78–82. [Google Scholar] [CrossRef]
- Forner-Giner, M.A.; Rodriguez-Gamir, J.; Martinez-Alcantara, B.; Quinones, A.; Iglesias, D.J.; Primo-Millo, E.; Forner, J. Performance of Navel orange trees grafted onto two new dwarfing rootstocks (Forner-Alcaide 517 and Forner-Alcaide 418). Sci. Hortic. 2014, 179, 376–387. [Google Scholar] [CrossRef]
- Gora, J.S.; Kumar, R.; Sharma, B.D.; Ram, C.; Berwal, M.K.; Singh, D.; Bana, R.S.; Kumar, P. Performance evaluation of Fremont mandarin on different rootstocks under the hot arid environment of India. South Afr. J. Bot. 2022, 144, 124–133. [Google Scholar] [CrossRef]
- Baron, D.; Esteves Amaro, A.C.; Pina, A.; Ferreira, G. An overview of grafting re-establishment in woody fruit species. Sci. Hortic. 2019, 243, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.M.; Watson, A.E.; van Hooijdonk, B.M.; Schaffer, R.J. Key flowering genes including FT-like genes are upregulated in the vasculature of apple dwarfing rootstocks. Tree Genet. Genomes 2014, 10, 189–202. [Google Scholar] [CrossRef]
- Nandwani, D.; Kuniyuki, A.H. Grafting and improvement of breadfruit production in micronesia. In International Symposium on Harnessing the Potential of Horticulture in the Asian-Pacific Region; ISHS Acta Horticulturae: Coolum, QLD, Australia, 2005; Volume 694, pp. 307–310. [Google Scholar]
- Medagoda, I.; Chandrarathna, K.W.M.C.J. Grafting of breadfruit (artocarpus altilis) using breadnut (Artocarpus camansi) as root stock. Acta. Hortic. 2007, 757, 149–152. [Google Scholar] [CrossRef]
- Solomon, F.J.; Roberts-Nkrumah, L.B. An evaluation of factors influencing successful grafting of breadfruit on chataigne rootstock. Proc. Caribb. Food Crops Soc. 2008, 44, 304–312. [Google Scholar]
- Zhou, Y.; Underhill, S.J.R. Plasma membrane H -ATPase activity and graft success of breadfruit (Artocarpus altilis) onto interspecific rootstocks of marang (A. odoratissimus) and pedalai (A. sericicarpus). Plant Biol. 2018, 20, 978–985. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S.J.R. A dwarf phenotype identified in breadfruit (Artocarpus altilis) plants growing on marang (A. odoratissimus) rootstocks. Horticulturae 2019, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Underhill, S.J.R. Differential transcription pathways associated with rootstock-induced dwarfing in breadfruit (Artocarpus altilis) scions. BMC Plant Biol. 2021, 21, 261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Underhill, S.J.R. Expression of gibberellin metabolism genes and signalling components in dwarf phenotype of breadfruit (Artocarpus altilis) plants growing on marang (Artocarpus odoratissimus) rootstocks. Plants 2020, 9, 634. [Google Scholar] [CrossRef] [PubMed]
- Zerega, N.; Supardi, M.N.N.; Motley, T.J. Phylogeny and recircumscription of Artocarpeae (Moraceae) with a focus on Artocarpus. Syst. Bot. 2010, 35, 766–782. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhou, Z.; Gilbert, M.G. Moraceae. Flora China Efloras.Org 2003, 5, 21–73. [Google Scholar]
- Joshee, N.; Bastola, D.; Agrawal, V.; Yadav, A. Lakoocha: A multipurpose tree of warm climate. In Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002. [Google Scholar]
- Bishnoi, S.; Shinde, R.; Sarkar, P. Monkey Jack (Artocarpus lakoocha Roxb.): Hope for Sustaining Livelihood. In Fruits for Livelihood: Production Technology and Management Practices; Dutta, A.K., Mondal, B., Eds.; Agrobios (India): Jodhpur, India, 2017; pp. 199–213. [Google Scholar]
- Gupta, A.K.; Rather, M.A.; Kumar Jha, A.; Shashank, A.; Singhal, S.; Sharma, M.; Pathak, U.; Sharma, D.; Mastinu, A. Artocarpus lakoocha Roxb. and Artocarpus heterophyllus Lam. flowers: New sources of bioactive compounds. Plants 2020, 9, 1329. [Google Scholar] [CrossRef]
- Birnberg, P.R.; Brenner, M.L. A one-step enzymatic assay for sucrose with sucrose phosphorylase. Anal. Biochem. 1984, 142, 556–561. [Google Scholar] [CrossRef]
- Webster, A.D. Rootstock and interstock effects on deciduous fruit tree vigour, precocity, and yield productivity. N. Z. J. Crop Hortic. Sci. 1995, 23, 373–382. [Google Scholar] [CrossRef]
- Hayat, F.; Li, J.; Liu, W.; Li, C.; Song, W.; Iqbal, S.; Khan, U.; Umer Javed, H.; Ahsan Altaf, M.; Tu, P.; et al. Influence of Citrus Rootstocks on Scion Growth, Hormone Levels, and Metabolites Profile of ‘Shatangju’ Mandarin (Citrus reticulata Blanco). Horticulturae 2022, 8, 608. [Google Scholar] [CrossRef]
- Gao-Takai, M.; Katayama-Ikegami, A.; Nakano, S.; Matsuda, K.; Motosugi, H. Vegetative Growth and Fruit Quality of ‘Ruby Roman’ Grapevines Grafted on Two Species of Rootstock and Their Tetraploids. Hortic. J. 2017, 86, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Marini, R.P.; Fazio, G. Apple rootstocks: History, physiology, management, and breeding. In Horticultural Reviews; Warrington, I., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2018; Volume 45, pp. 197–312. [Google Scholar]
- Clearwater, M.; Seleznyova, A.; Thorp, T.; Blattmann, P.; Barnett, A.; Lowe, R.; Austin, P. Vigor-controlling rootstocks affect early shoot growth and leaf area development of kiwifruit. Tree Physiol. 2006, 26, 505–515. [Google Scholar] [CrossRef]
- Seleznyova, A.N.; Thorp, T.G.; White, M.; Tustin, S.; Costes, E. Application of architectural analysis and AMAPmod methodology to study dwarfing phenomenon: The branch structure of ‘Royal Gala’ apple grafted on dwarfing and non-dwarfing rootstock/interstock combinations. Ann. Bot. 2003, 91, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina, A.; Cookson, S.; Calatayud, A.; Trinchera, A.; Errea, P. Physiological and molecular mechanisms underlying graft compatibility. In Vegetable Grafting: Principles and Practices; Colla, G., Perez-Alfocea, F., Schwarz, D., Eds.; CAB International: Cambridge, UK, 2017; pp. 132–154. [Google Scholar]
- Gautier, A.T.; Chambaud, C.; Brocard, L.; Ollat, N.; Gambetta, G.A.; Delrot, S.; Cookson, S.J. Merging genotypes: Graft union formation and scion-rootstock interactions. J. Exp. Bot. 2019, 70, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, S.; Pina, A.; Fevereiro, P.; Kragler, F. A phenotypic search on graft compatibility in grapevine. Agronomy 2020, 10, 706. [Google Scholar] [CrossRef]
- Zarrouk, O.; Gogorcena, Y.; Moreno, M.A.; Pinochet, J. Graft compatibility between peach cultivars and prunus rootstocks. HortScience 2006, 41, 1389. [Google Scholar] [CrossRef]
- Reig, G.; Salazar, A.; Zarrouk, O.; Forcada, C.F.I.; Val, J.; Moreno, M.A. Long-term graft compatibility study of peach-almond hybrid and plum based rootstocks budded with European and Japanese plums. Sci. Hortic. 2019, 243, 392–400. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Pina, A.; Errea, P. A review of new advances in mechanism of graft compatibility–incompatibility. Sci. Hortic. 2005, 106, 1–11. [Google Scholar] [CrossRef]
- Yoon, J.; Cho, L.H.; Tun, W.; Jeon, J.S.; An, G. Sucrose signaling in higher plants. Plant Sci. Int. J. Exp. Plant Biol. 2021, 302, 110703. [Google Scholar] [CrossRef]
- Zhou, Y.; Chan, K.; Wang, T.; Hedley, C.; Offler, C.; Patrick, J. Intracellular sucrose communicates metabolic demand to sucrose transporters in developing pea cotyledons. J. Exp. Bot. 2009, 60, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Lochard, R.G.; Schneider, G.W. Stock and scion growth relationships and the dwarfing mechanism in apple. Hortic. Rev. 1981, 3, 315–375. [Google Scholar] [CrossRef]
- Zhou, Y.; Tian, X.; Yao, J.; Zhang, Z.; Wang, Y.; Zhang, X.; Li, W.; Wu, T.; Han, Z.; Xu, X.; et al. Morphological and photosynthetic responses differ among eight apple scion-rootstock combinations. Sci. Hortic. 2020, 261, 108981. [Google Scholar] [CrossRef]
- Vriet, C.; Smith, A.M.; Wang, T.L. Root starch reserves are necessary for vigorous re-growth following cutting back in Lotus japonicus. PLoS ONE 2014, 9, e87333. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Graft | Self-Graft | On Lakoocha | |
|---|---|---|---|
| Final scion height (cm) | 151.17 ± 8.60 b | 156.67 ± 10.14 b | 47.97 ± 5.05 a |
| Main stem diameter (cm) | 2.91 ± 0.80 b | 3.10 ± 0.78 b | 1.46 ± 0.37 a |
| Node number on main stems | 28.22 ± 4.53 a | 27.04 ± 4.61 a | 22.48 ± 2.56 a |
| Number of branches per plant | 2.20 ± 0.42 b | 2.42 ± 0.27 b | 0.61 ± 0.45 a |
| Main stem elongation rate (cm/month) | 7.32 ± 2.19 b | 6.96 ± 0.75 b | 2.21 ± 0.62 a |
| Second internode length in Main stems (cm) | 5.62 ± 1.20 b | 5.91 ± 1.25 b | 2.25 ± 0.23 a |
| Leaf numbers per plant | 7.23 ± 0.53 b | 8.40 ± 0.60 b | 4.00 ± 0.35 a |
| Leaf chlorophyll a (mg/g) | 2.22 ± 0.55 a | 2.49 ± 0.61 a | 2.12 ± 0.42 a |
| Leaf chlorophyll b (mg/g) | 0.83 ± 0.10 a | 0.87 ± 0.12 a | 1.32 ± 0.34 a |
| leaf chlorophyll a + b (mg/g) | 2.76 ± 0.81 a | 3.36 ± 0.61 a | 3.44 ± 0.63 a |
| Leaf chlorophyll a/b | 3.04 ± 0.56 a | 3.01 ± 0.88 a | 1.97 ± 0.47 a |
| Leaf length | 82.55 ± 1.16 b | 78.50 ± 3.18 b | 55.89 ± 5.90 a |
| Leaf width | 58.50 ± 2.43 b | 60.25 ± 2.56 b | 39.25 ± 3.67 a |
| Stem diameter at 5 cm above graft union | 3.25 ± 0.25 a | 1.71 ± 0.10 b | |
| Stem diameter at 5 cm below graft union | 3.31 ± 0.34 a | 1.60 ± 0.16 b | |
| Stem diameter difference above and below graft union | 0.18 ± 0.06 a | 0.17 ± 0.09 a | |
| Ratio of stem diameter above and below graft union | 0.99 ± 0.04 a | 1.09 ± 0.08 a | |
| Final survival (%) | 59.72 ± 6.15 a | 40.05 ± 4.85 b | |
| 1-year graft compatibility (%) | 81.25 ± 8.84 a | 68.75 ± 8.84 a | |
| Final graft compatibility (%) | 75.00 ± 0.00 a | 62.50 ± 0.00 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Underhill, S.J.R. Characterisation of Breadfruit (Artocarpus altilis) Plants Growing on Lakoocha (A. lakoocha) Rootstocks. Horticulturae 2022, 8, 916. https://doi.org/10.3390/horticulturae8100916
Zhou Y, Underhill SJR. Characterisation of Breadfruit (Artocarpus altilis) Plants Growing on Lakoocha (A. lakoocha) Rootstocks. Horticulturae. 2022; 8(10):916. https://doi.org/10.3390/horticulturae8100916
Chicago/Turabian StyleZhou, Yuchan, and Steven J. R. Underhill. 2022. "Characterisation of Breadfruit (Artocarpus altilis) Plants Growing on Lakoocha (A. lakoocha) Rootstocks" Horticulturae 8, no. 10: 916. https://doi.org/10.3390/horticulturae8100916
APA StyleZhou, Y., & Underhill, S. J. R. (2022). Characterisation of Breadfruit (Artocarpus altilis) Plants Growing on Lakoocha (A. lakoocha) Rootstocks. Horticulturae, 8(10), 916. https://doi.org/10.3390/horticulturae8100916