
Methods and Factors Influencing In Vitro Propagation Efficiency of Ornamental Tuberose (Polianthes Species): A Systematic Review of Recent Developments and Future Prospects

 ,
,  ,
,
Abstract
:1. Introduction
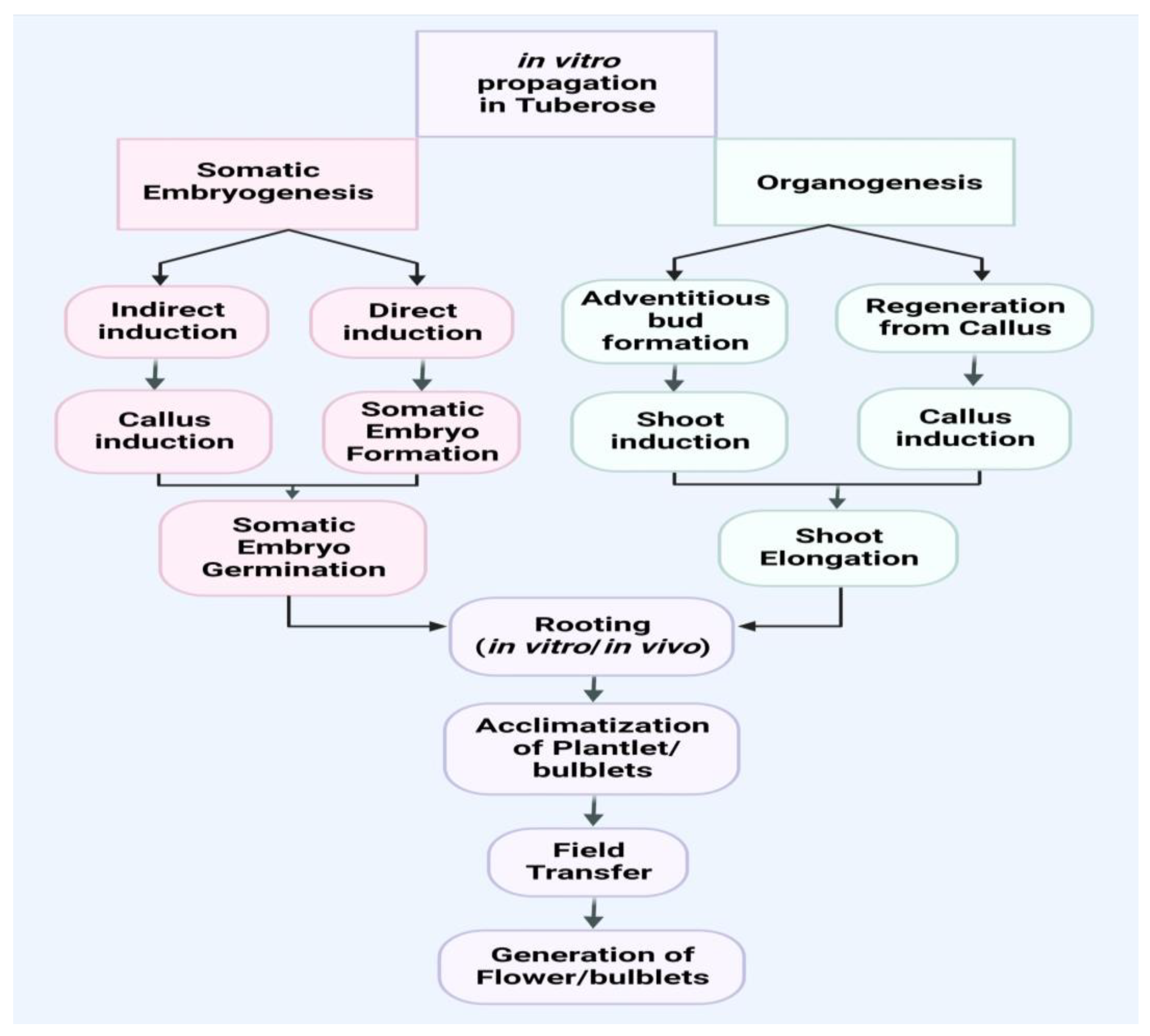
2. In Vitro Propagation
2.1. Somatic Embryogenesis in Cell Suspension
2.2. Organogenesis
2.2.1. Direct Organogenesis
2.2.2. Indirect Organogenesis
3. Stages in the Micro-Propagation Process
3.1. Initiation of Aseptic Cultures and Factors Affecting In Vitro Establishment
3.1.1. Genotype
3.1.2. Selection and Type of Explants
3.1.3. Sanitation Conditions of Mother Plant and Explant Sterilization
3.2. Shoot Multiplication Stage
3.2.1. Factors Affecting the Shoot Multiplication Stage
Genotypes/Cultivars/Species
Media
Size and Position of Explants
Growth Regulators
Carbon Source
Gelling Agents
Inorganic Salts, Organic Compounds, Antibiotics, and Vitamins
Incubation Conditions and Environmental Factors
4. In Vitro Rooting of Shoots and Ex Vitro Rooting
4.1. Factors Affecting In Vitro Rooting of Shoots
4.1.1. Species/Cultivars/Genotype/Varieties
4.1.2. Type of Explants
4.1.3. Media
4.1.4. Inorganic Salts/Vitamins/Activated Charcoal
4.1.5. Carbohydrate/Carbon Source
4.1.6. Gelling Agent
4.1.7. Growth Regulators
4.1.8. Ex Vitro Rooting
5. Acclimatization and Field Transfer
6. In vitro Bulb/Rhizome Formation
7. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lin, T.S.; Shen, T.M. Study on karyotype of Polianthes tuberosa L. J. Agric. For. NCYU 2004, 1, 1–12. [Google Scholar]
- Karihaloo, J.L. Cytological and hybridization studies in three cultivars of tuberose (Polianthes tuberosa L.). Cytologia 2019, 84, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.P.; Uma, S. Studies on ratoon crop in tuberose cv. Single and Double. Indian Perfum. 1995, 39, 158–160. [Google Scholar]
- Espejo, S.A. Las Monocotiledoneas Mexicanas: Una Synopsis Floristica [Enlinea]; Serna, A.E.S., López-Ferrari, A.R., Eds.; UAM, Consejo Nacional de la Flora de México, A.C.: Iztapalapa, México, 1993; p. 112. [Google Scholar]
- Datta, S.K. Breeding of ornamentals: Tuberose (Polianthes tuberosa L.). Curr. Sci. 2017, 113, 1255–1263. [Google Scholar] [CrossRef]
- Misra, R.L.; Mahesh, K.S. Bulbous ornamental breeding. Adv. Hortic.-Ornam. Plants 1995, 12, 475–494. [Google Scholar]
- Ahmad, M.S.; Ahmad, T.; Zaidi, N.; Nasir, I.A. High frequency in vitro propagation of Polianthes tuberosa. Pak. J. Sci. Ind. Res. 2006, 49, 344–348. [Google Scholar]
- Khan, N.H.; Zaidi, N.; Jabeen, S.; Javaid, I. Micropropagation potential of Polianthes tuberosa L. bulb scales and leaves. Pak. J. Sci. Ind. Res. 2000, 43, 118–122. [Google Scholar]
- Rajasekharan, V.; Haripriya, K.; Arumugam, S.; Shakila, A. In vitro propagation of tuberose (Polianthes tuberosa L.). In Proceedings of the Centennial Conference on Spices and Aromatic Plants: Challenges and Opportunities in the New Century, Calicut, India, 20–23 September 2000; pp. 86–88. [Google Scholar]
- Nazneen, S.; Jabeen, M.; Ilahi, I. Micropropagation of Polianthus tuberosa (tuberose) through callus formation. Pak. J. Bot. 2003, 35, 17–25. [Google Scholar]
- Mishra, A.; Pandey, R.K.; Gupta, R.K. Micropropagation of tuberose (Polianthus tuberosa L.) cv Calcattia Double. Progress. Hortic. 2005, 37, 226–236. [Google Scholar]
- Jyothi, R.; Singh, A.K.; Singh, K.P. In vitro propagation studies in tuberose (Polianthes tuberosa L.). J. Ornam. Hortic. 2008, 11, 196–201. [Google Scholar]
- Raghuvanshi, S.; Tripathi, M.K.; Vidhya-Sankar, M.; Singh, O.P. Establishment of low-cost effective protocol for massive in vitro propagation in Polianthes tuberosa L. Plant Cell Biotechnol. Mol. Biol. 2013, 14, 49–59. [Google Scholar]
- Taksande, P.N.; Patil, S.R.; Rathod, A.D.; Karad, G.W.; Sayyad, R.A.; Jayade, V.S. Direct shoot organogenesis from stem disc explants of tuberose (Polianthes tuberose L.). J. Soils Crops 2018, 28, 157–164. [Google Scholar]
- Nalousi, A.M.; Hatamzadeh, A.; Azadi, P.; Mohsenpour, M.; Lahiji, H.S. A procedure for indirect shoot organogenesis of Polianthes tuberosa L. and analysis of genetic stability using ISSR markers in regenerated plants. Sci. Hortic. 2019, 244, 315–321. [Google Scholar] [CrossRef]
- Cheesman, L.; Finnie, J.F.; Van Staden, J. Eucomiszambesiaca baker: Factors affecting in vitro bulblet induction. South Afr. J. Bot. 2010, 76, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Ascough, G.D.; Erwin, J.E.; Van Staden, J. Reduced temperature, elevated sucrose, continuous light and gibberellic acid promote corm formation in Watsonia vanderspuyiae. PCTOC 2008, 95, 275–283. [Google Scholar] [CrossRef]
- Aitken-Christie, J.; Kozai, T.; Smith, M.A.L. Automation and Environmental Control in Plant Tissue Culture; Springer Science + Business Media: Dordrecht, The Netherlands, 1995; p. 500. [Google Scholar]
- Krishnamurthy, K.B.; Mythili, J.B.; Srinivas, M. Micro-propagation studies in ‘single’ vs. ‘double’ types of tuberoses (Polianthes tuberosa L.). J. Appl. Hortic. 2001, 3, 82–84. [Google Scholar] [CrossRef]
- Bindhani, B.K.; Dalai, A.K.; Behera, B. In vitro multiple shoot induction in Polianthes tuberosa L. using shoot bud explants. Plant Sci. J. 2004, 26, 24–27. [Google Scholar]
- Ali, M.R.; Akand, M.H.; Hoque, M.E.; Homayra, H.; Mehraj, H.; Jamal Uddin, A.F.M. In vitro regeneration and rapid multiplication of tuberose. Int. J. Bus. Soc. Sci. Res. 2015, 3, 35–38. [Google Scholar]
- Khanchana, K.; Kannan, M.; Hemaprabha, K.; Ganga, M. Standardization of protocol for sterilization and in vitro regeneration in tuberose (Polianthes tuberose L.). Int. J. Chem. Stud. 2019, 7, 236–241. [Google Scholar]
- Copetta, A.; Marchioni, I.; Mascarello, C.; Pistelli, L.; Cambournac, L.; Dimita, R.; Ruffoni, B. Polianthes tuberosa as edible flower: In vitro propagation and nutritional properties. Int. J. Food Eng. 2020, 6, 57–62. [Google Scholar] [CrossRef]
- Kahrizi, D.; Barzegar, B.; Azadi, P. Study on somatic embryogenesis in Polianthes tuberosa. J. Biotechnol. 2008, 136, S150. [Google Scholar] [CrossRef]
- Bindhani, B.K.; Dalal, A.K.; Behara, B. Role of auxins for callus induction and chromosomal variation in Polianthes tuberosa L. ‘Single’. Indian J. Genet. Plant Breed. 2004, 64, 173–174. [Google Scholar]
- Kadam, G.B.; Singh, K.P.; Jyoti, R. Role of sterilants in establishment of aseptic culture using different explants in tuberose (Polianthes tuberosa L.). Progress. Hortic. 2011, 43, 105–109. [Google Scholar]
- Hutchinson, M.J.; Onamu, R.; Obukosia, S. Effect of thidiazurone, benzylaminopurine and naphthalene acetic acid on in vitro propagation of tuberose (Polianthes tuberosa L.) from shoot tip explants. J. Agric. Sci. Technol. 2004, 6, 48–59. [Google Scholar]
- Sangavai, C.; Chellapandi, P. In vitro propagation of tuberose plant (Polianthes tuberosa L.). Electron. J. Biol. 2008, 4, 98–101. [Google Scholar]
- Samanta, A.; Maity, T.R.; Jana, D.; Saha, B.; Datta, S. Standardization of in vitro propagation of Polianthes tuberosa L. (calcutta double). J. Plant Dev. Sci. 2015, 7, 889–891. [Google Scholar]
- Kumari, N.; Pal, A. Multiple shoot regenera tiple shoot regenera tiple shoot regeneration of tuberose (Polianthes tuberosa L.) cv Prajwal by using bulb as explant. Bioscan 2016, 11, 1407–1410. [Google Scholar]
- Narayanaswamy, S.; Prabhudesai, V.R. Somatic psedoembryogeny in tissue cultures of tuberose (Polianthes tuberose L.). Indian J. Exp. Biol. 1979, 17, 873–875. [Google Scholar]
- Upadhyay, G.K.; Bindhani, B.K.; Behera, B. In vitro micropropagation of two varieties of Polyanthes tuberosa L. Plant Sci. J. 2001, 23, 25–28. [Google Scholar]
- Bindhani, B.K.; Dalai, A.K.; Behera, B. Photomorphogenetic effect on callus initiation in tuberose (Polianthes tuberosa L.). Plant Sci. J. 2002, 24, 43–45. [Google Scholar]
- Krishnan, A.G. In vitro Multiplication and Genetic Improvement of Tuberose (Polianthes tuberosa L.). Ph.D. Thesis, Kerala Agricultural University, Kerala, India, 2003. [Google Scholar]
- Surendranath, R.; Ganga, M.; Jawaharlal, M. In vitro propagation of tuberose. Ecol. Environ. 2016, 34, 2556–2560. [Google Scholar]
- Singh, K.B.M.; Madhavan, J.; Sadhukhan, R.; Chandra, S.; Rao, U.; Mandal, P.K. Production of nematode free plantlets in Polianthes tuberosa using in vitro culture techniques. Hortic. Environ. Biotechnol. 2020, 61, 929–937. [Google Scholar] [CrossRef]
- Beyrami, Z.E.; Azadi, P.; Safari, A.; Shafii, M.R.; Sadeqi, S. Study on Somoclonal Variation in Polianthes Tuberosa In vitro Culture; The National Ornamental Plant Research Station: Mahallat, Iran, 2008; p. 102. Available online: http://agris.fao.org/agris-search/search.do?recordID=IR2010000115 (accessed on 12 April 2022).
- Ali, M.R.; Mehraj, H.; Jamal Uddin, A.F.M. Kinetin (KIN) and Indole-3-acetic acid (IAA) on in vitro shoot and root initiation of tuberose. Int. J. Sustain. Agric. Res. 2014, 10, 1–4. [Google Scholar]
- Naz, S.; Aslam, F.; Ilyas, S.; Shahzadi, K.; Tariq, A. In vitro propagation of tuberose (Polianthes tuberose L.). J. Med. Plant Res. 2012, 6, 4107–4112. [Google Scholar]
- Lindsey, K. Plant Tissue Culture Manual, 1st ed.; Kluwer Academic Publishing: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Datta, S.K.; Misra, P.; Mandal, A.K.; Chakrabarty, D. Direct shoot organogenesis from different explants of chrysanthemum, marigold, and tuberose. Isr. J. Plant Sci. 2002, 50, 287–291. [Google Scholar] [CrossRef]
- Pohare, M.; Rathod, H.P.; Shahakar, S.B.; Kelatkar, S.K.; Suryawanshi, P. Effects of UV radiations on morphological characters in in vitro regenerated Polianthes tuberosa. Res. J. Agric. Sci. 2012, 3, 1307–1308. [Google Scholar]
- Jala, A.; Kachonpadungkitti, Y. Tuberose (Polianthes tuberose L.) shoots multiplying and callus induction by benzyladenine, naphthaline acetic acid and oryzaline. Thammasat Int. J. Sci. Technol. 2014, 19, 15–20. [Google Scholar]
- Gajbhiye, S.S.; Tripathi, M.K.; Vidya, M.; Singh, M.S.; Baghel, B.S.; Tiwari, S. Direct shoot organogenesis from cultured stem disc explants of tuberose (Polianthes tuberosa L.). J. Agric. Sci. Technol. 2011, 7, 695–709. [Google Scholar]
- Estrada-Basaldua, J.; Pedraza-Santos, M.; De La Cruz Torres, E.; Martínez-Palacios, A.; Saenz-Romero, C.; Morales-García, J. Effect of 60Co gamma rays in tuberose (Polianthes tuberosa L.). Rev. Mex. Cienc. Agric. 2011, 2, 445–458. [Google Scholar]
- Peña, A.Y.T.; López, L.L.P.; Couoh, E.V.; Ramírez, A.R.; López, D.R. Axenic establishment and in vitro formation of adventitious shoots in nardo (Polianthes tuberose L.). In Proceedings of the Biotechnology Summit, Mérida, Mexico, 12–21 March 2012; pp. 170–173. [Google Scholar]
- Surendranath, R.; Ganga, M.; Ranjitha, G. Can in vitro contaminated culture be revived—A case study with contaminated cultures of tuberose (Polianthes tuberosa). Curr. Biot. 2015, 9, 285–288. [Google Scholar]
- Panigrahi, J.; Rana, V.D.; Patel, I. Multiple shoot regeneration of Polianthes tuberosa cultivars Phulerajni and Calcutta double. Paripex-Indian J. Sci. Res. 2013, 3, 12–14. [Google Scholar]
- Otsuji, K.; Honda, Y.; Sugumira, Y.; Takei, A. Production of polysaccharides in liquid cultures of Polianthes tuberosa cells. Biotechnol. Lett. 1994, 16, 943–948. [Google Scholar] [CrossRef]
- Honda, Y.; Inaoka, H.; Takei, A.; Sugimura, Y.; Otsuji, K. Extracellular polysaccharides produced by tuberose callus. Phytochemistry 1996, 41, 1517–1521. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- White, P.R. The Cultivation of Animal and Plant Cells, 2nd ed.; Ronald Press: New York, NY, USA, 1963. [Google Scholar]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Dehdezi, A.A.; Mousavi, S.D.; Azadi, P. Evaluation of different growth regulators on proliferation of (Polianthes tuberosa). Bull. Environ. Pharmacol. Life Sci. 2014, 3, 172–174. [Google Scholar]
- Hernández-Mendoza, F.; Carrillo-Castañeda, G.; Pedraza-Santos, M.E.; de la Cruz-Torres, E.; del Carmen Mendoza-Castillo, M. Regeneration in vitro of shoots Polianthes tuberosa L. through vegetative buds of the inflorescence and from tissue corm. Nova Sci. 2014, 7, 32–47. [Google Scholar] [CrossRef]
- Hernández-Mendoza, F.; Carrillo-Castañeda, G.; García-Gaytán, V.; Pedraza-Santos, M.E.; de la Cruz-Torres, E.; del Carmen Mendoza-Castillo, M. In vitro plant regeneration of Polianthes tuberosa L. from leaf and flower buds tissue. Trop. Subtrop. Agroecosyst. 2021, 24, 55. [Google Scholar]
- Wang, P.J.; Hu, C.Y. Regeneration of virus free plants through in vitro culture. In: Fiechter, A. (ed.). Adv. Biochem. Eng. 1980, 18, 61–99. [Google Scholar]
- Panigrahi, J.; Saiyad, M.S.L. In vitro propagation of Polianthes tuberosa L. cultivars (Calcutta Single). Int. J. Plant Anim. Env. Sci. 2013, 3, 76–79. [Google Scholar]
- Kadam, G.B.; Singh, K.P.; Singh, A.K.; Jyothi, R. In vitro regeneration of tuberose through petals and immature flower buds. Indian J. Hortic. 2010, 67, 76–80. [Google Scholar]
- Panigrahi, J.; Chaudhury, A.A. In vitro propagation of hybrid cultivars (Hyderabad Single & Local Double Navsari) of ornamental plant Polianthes tuberosa L. GRA—Glob. Res. Anal. 2013, 2, 3–4. [Google Scholar]
- Kozai, T.; Kubota, C.; Jeong, B.R. Environmental control for the large-scale production of plants through in vitro techniques. Plant Cell Tissue Organ Cult. 1997, 51, 49–56. [Google Scholar] [CrossRef]
- Pohare, M.; Batule, B.; Bhor, S.; Shahakar, S.B.; Kelatkar, S.K.; Varandani, S. Effect of gamma radiations on the morphological characters in in vitro regenerated Polianthes tuberosa. Indian J. Hort. 2013, 3, 95–97. [Google Scholar]
- Bose, T.K.; Jana, B.K.; Moulik, S. A note on micropropagation of tuberose from stem scale section. Indian J. Hortic. 1987, 42, 100–101. [Google Scholar]
- Misra, P.; Saema, S. Plant tissue culture for in vitro mutagenesis, large-scale propagation, and genetic transformation. In Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Anis, M., Ahmad, N., Eds.; Springer: Singapore, 2016; pp. 309–342. [Google Scholar]
- Stefaniak, B. Somatic embryogenesis and plant regeneration of Gladiolus (Gladiolus hort.). Plant Cell Rep. 1994, 13, 386–389. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Cultivar(s) and Species | Explant Used (Type, Size, Origin) | Sterilization | Basal Medium | PGR (Type and Concentration, mg/Lor molar) | Other Medium Additives | Other Culture Conditions | Effect on Tissue Culture, Development, etc. (Regeneration Type) | Productivity, Somaclonal Variation and Abnormalities | Acclimatization (% Survival) | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Single and double varieties of tuberose (Polianthes tuberosa L.) | Bulbs | 70% ethanol for 30 s followed by HgCl2 0.1% for 6 min | MS medium | Diff. conc. of Kinetin and BAP (1, 2, 3, 4 mg L−1), 3.0 mg L−1 Kin (best SI and SEL), 2.0 mg L−1 IBA or 3.0 mg L−1 NAA (best RI). | 0.8% agar, 3% sucrose, and deproteinized coconut water (15%). | Incubated at-temp. 25 ± 20 °C, light intensity of 2000–3000 lux, with 16 h photoperiod, and at 75% RH. | Shoot induction and root induction. | - | - | [9] |
| - | Bulb pieces and in vitro formed shoot buds | 70% ethanol for 2 min and 1% HgCl2 for 3–4 min. | MS | 0.5 mg L−1 BAP + 0.5 mg L−1 2, 4-D (bulb to CI), 1 mg L−1 BAP + 1 mg L−1 2, 4-D (shoot bud to CI), 3 mg L−1 BAP (callus to SI), 0.5 mg L−1 NAA (shoots to RI). | 4% sucrose, 0.9% agar. | pH 5.6, autoclaved at 15 psi for 15 min, cultured at 25 ± 1 °C on 16 h photoperiod. | Callus induction, shooting and rooting, transfer to soil. | - | 100% survival rate observed. | [10] |
| Calcuttia Doubles tuberose (Polianthes tuberosa L.) | Radial cutting of bulblets | 0.1% HgCl2 for 6 min. followed by NaOCI 1% for 8 min. | Modified MS | 4.0 mg L−1 BAP and 0.2 mg L−1 IBA (SM), ½ basal MS medium + 0.1 mg L−1 NAA (best RI), 1.0 mg L−1 IBA (maximum RL and number of roots). | pH 5.7; ± 0.1, sucrose-3%, agar-0.7%, 0.6% bactoagar (for rooting), autoclaved at 121 °C (0.1 M Pa). | Temp. 25 + 1 °C under 16 h photoperiod (3000 lux) and 8 h dark period. | - | - | Highest survival (86.6%) was achieved after transplantation when rooted plantlet leaves were treated with 1.0% glycerol for 15 min before planting in sterilized potting mixture containing sand + soil + FYM (1:1:1). | [11] |
| cv. Calcuttia Double (Polianthes tuberose.) | Radial cutting of bulblets | 0.1% HgCl2 for 6 min followed by NaOCl 1% for 8 min. | MS | 4.0 mg L−1 BAP + 0.2 mg L−1 IBA (SI), ½ MS + 0.1 mg L−1 NAA (best RI), 1 mg L−1 IBA (max. RL and root number). | 3% sucrose, 0.7 % agar (shoots), 0.6% bactoagar (rooting). | pH 5.7 ± 0.1, autoclaved at 121 °C, cultured at-25 ± 1 °C. | Effect of surface sterilants, effect of growth regulators on shoots and roots form ation, effect of Glycerol and ABA on survival of plantlets. | - | Plantlets were potted in sterilized sand-FYM-loam soil substrate (1:1:1) after treatment with diff. conc. of glycerol and ABA for 15 min and covered with polythene bags, maximum survival (86%) was observed in 1% glycerol. | [11] |
| Single and double cultivars, namely Prajwal and Vaibhav (Polianthes tuberosa L.), | Bulb scales (1 cm) and bud (0.5 cm) | Fungicides, namely carbendazim (0.1%), mancozeb (0.1%), and bactericide 8-HQC (200 mg L−1) in different combinations for 2–4 h; 0.1% HgCl2 and 1.0 % NaOCl for 6–8 min. | MS | Diff. conc. of BAP (4–6 mg L−1), IAA (0.2–0.5 mg L−1) and NAA (0.2–0.5 mg L−1) in combinations or alone, 4 mg/LBAP (SM mg L−1), ½ MS + 1.0 mg/L IBA (RI). | 30 g/L sucrose. | Shoot induction, root induction and acclimatization. | - | - | Acclimatized in glass jars or plastic pots containing mixture of Peat + Solarite (1:1 v/v). | [12] |
| cvs. Nagpur local 1, Nagpur local 2, Amravati local 1 (Polianthes tuberosa L.) | Stem disc | 2–3 drops of Tween-20 followed by 90% ethanol for 30 s, followed by 0.1% HgCl2 with 1% Bavistin for 10 min. | White’s media | 28 different compositions of BAP, Kinetin, BAP + NAA, Kinetin + NAA (SI), 12 diff. conc. of IAA and NAA (rooting), 4.0 mg L−1 BAP + 0.5 mg L−1 NAA (best SI), 2.0 mg L−1 IAA (best RI). | 2% sucrose, 1% agar. | pH 5.8, cultured at 25 ± 2 °C and 16 h light. | Direct organogenesis and hardening of plantlets. | - | Maximum survival rate was in Nagpur local 1 (53.84%), followed by Nagpur local 2 (40.90%) and Amravati local 1 (36.36%). | [14] |
| cv. Pearl Double (Polianthes tuberosa L.) | Various explants (crown explants, leaves, petals, flowering stems, and bulb scales) | 70% (v/v) ethanol for 30 s followed by 4% (w/v) NaOCl for 30 min. | MS | 1.95 μM 2,4,5-T (callus), 2.26 μM TDZ + 250 mg/L proline (CI to SI), 2.22 μM BAP (SEL), 9.89 μM IBA (SI to RI). | 3% sucrose, 7 g/L agar. | pH 5.8 autoclaved at 121 °C and 1.5 kg cm−1 pressure for 20 min, incubated at 24 ± 2 °C in the light intensity of 80 μ mol m-2 s-1 and 60–65% RH. | Callus induction, shoot regeneration from callus, shoot elongation and rooting, acclimatization and transplantation, flow cytometry analysis, evaluation of genetic stability. | Higher similarity of ploidy level between regenerated and mother plants was observed. | Plantlets that were well rooted successfully hardened and adapted in the greenhouse condition with a survival rate of 100%. | [15] |
| Single (Shringar) and double (Suvasini) types of tuberoses (Polianthes tuberosa L.) | Terminal and axillary stem scale section | Bavistin (1000 ppm) + Kavach (1000 ppm) + Cetrimide (500 ppm) overnight followed by HgCl2 (0.1%) for 15 min (effective to remove contamination). | MS and ½ MS | 2 mg/L BAP and 0.1 mg/L IAA (regeneration), 1/2 MS + 0.2 mg L-1 IAA + 0.25 mg/L IBA (RI), IBA 0.5 mg L−1 (RL). | 3% sucrose, 0.25% Phytagel. | - | Decontamination of cultures, multiple shoot induction, in vitro rooting. | - | Both the cultivars could be hardened with 100% survival using the closed sachet technique. The hardened plants established with 100% success when transplanted to pots. | [19] |
| ‘Single’ and ‘Double’ (Polianthes tuberosa L.) | Shoot buds | 0 1% HgCl2 for 15 min. | MS | Varied concentrations of different phyto-hormones, such as 2,4-D, BAP and IBA in various combination, 4 mg L−1 2,4-D + 0.5 mg/L BAP (highest % of shoot initiation), 4.0 mg L−1 2,4-D + 0.5 mg L−1 BAP (best SM), 0.5 mg L−1 IBA + 3% sucrose (best RI). | pH 5.7, agar-8 g/L, autoclaved at 121 °C with 1 05-kg/sq cm−1 pressure for 15 min., cultured at 25 ± 2 °C at 55% RH., 16/8 h photoperiod. | Shoot and root induction, shoot multiplication, field transfer of plants. | - | - | Survival rates of regenerated plants were highest in ‘Single’ (75%) and in ‘Double’ (65%) and least in ‘Swarna Rekha’ (10%). | [20] |
| - | Buds | 70% (v/v) ethanol for few seconds followed by immersing in 0.1% HgCl2 solution for 4–5 min followed by several washings. | MS | 0.5 mg L−1 IAA + 1.0 mg L−1 Kin (SI, RI). | - | Autoclaved at 121 °C for 20 min at 15 psi pressure, cultured at Temp. 25 ± 1 °C with the16 h photoperiod and 3000 lux light intensity. | - | Number of leaves/plantlet and length of leaves, number of roots/plantlet and length of roots, acclimatization and establishment of plantlets in soil. | Survival rates in small hole of plastic tray at growth chamber was 55.55% while in open atmosphere was 72.00%. | [21] |
| Four varieties viz., ArkaNirantara, Shringar, Calcutta Double, and Vaibhav (Polianthes tuberosa L.) | Axillary bud and bulblets | 70% ethanol for 30 s followed by 0.2% HgCl2 for 10 min, 5% NaOCl (10 min) and 0.5% AgNO3 (15 min). | MS | 2.0 mg L−1 BAP +0.5 mg/L NAA (SI and SEL), + 1 mg L−1 (IBA + NAA) (RI). | Pot mixture (1:1:1) (hardening). | Temp. 25 ± 2 °C, humidity 70–80%, light intensity: 2500–3000 Lux Photoperiod: 16/8 h light/dark. | Standardization of protocol for in vitro regeneration, in vitro shoot regeneration and multiplication, in vitro root induction, hardening of plants. | - | H1 pot mix [vermicompost: sand: cocopeat @ 1:1:1] recorded the highest survival of 97.17%. | [22] |
| Tuberose (Polianthes tuberosa L.) | Bulbs | 70% (v/v) ethanol solution for 30 min and then transferred in a 2.5 % (v/v) sodium hypochlorite (NaClO) solution for 20 min and finally rinsed twice in autoclaved water. | MS | 1.5 mg L−1 BA, 0.5 mg L−1 IAA. | 0.7% agar concentration with 3% sucrose. | pH 5.8 and kept at 23 ± 1 °C with a 16/8 h light/dark cycle and 209 ± 5 μmol m−2 s−1 light irradiances at tube level (white fluorescent Philips Master TL-D 36 W/840 lamp). | Multiplication of plantlets. | - | Acclimatized plantlets of four clones in two greenhouse sites (CREA and CREAM). Percentages of survival were 99.3 %, 99.4 %, 100.0%, and 98.9%, respectively. | [23] |
| Single and Double cultivars, namely Prajwal and Vaibhav (Polianthes tuberosa L.), | Bulb scale, buds, petal segment, and immature flower bud | HgCl2 (0.1%) and NaOCl (1.0%), for 6–8 min. | MS | Fungicides, namely carbendazim (0.1%), mancozeb (0.1%), and bactericide 8-HQC (200 mg L−1) in different combinations for 2–4 h for bulb scale and bud explants, pretreatment given to flower-based explants with same treatment, but with different time durations viz, 1–2 h. | 3% sucrose | pH 5.8 autoclaved at 121 °C for 21 min. | Role of sterilant in establishment of aseptic culture using different explants. | 0.1 per cent of carbendazim, mancozeb and 200 mg L−1 of 8-HQC for 4 h gave the maximum culture survival, in case of bulb scale and bud, petal segment and immature flower bud explants with same pre-treatment combination for 2 h, gave the maximum culture survival. | - | [26] |
| Pre-cooled rhizome (2 °C for 6 weeks) | Detergents solution for 5 min followed by surfactant treatment for 10 min. Then, the alcohols were washed for few min followed by 0.1% HgCl2 for 2–3 min. | MS | Diff. conc. Of IAA (0.5, 1, 2, 2,5, 3, 3.5 mg L−1) and/or 0.5 mg L−1 BAP in combination, 1.5 mg L−1 BAP (higher explants regeneration and shoot differentiation), 3.0 mg L−1 IAA + 0.5 mg L−1 BAP (Rhizome to CI), 1.5 mg L−1 BAP + 0.5 mg L−1 IAA (CI to shoot diff.). | 3% sucrose, 0.8% Difco agar. | pH 5.8, autoclaved at 120 °C for 20 min, 16 h light at 360 nmol·m-2·s-1 3500 Ingelec fluorescent tube, 27 °C day/21 °C night. | Callus induction, shoot regeneration, phytochemical analysis. | - | - | [28] | |
| Variety Calcutta double (Polianthes tuberosa L.) | Rhizomes | 1% Bavistin for 30 min followed by 2–3 drops of Tween-20 solutions for 10 min, 0.1% HgCl2 solution for 5 min. | MS | Various conc. of IAA (0.25, 0.50 and 0.75 mg L−1) and BAP (1, 2, 3, 4 and 5 mg L−1) in combination used for shoot proliferation, 0.5 mg L−1 IAA + 3 mg L−1 BAP (SM). | 3% sucrose, 0.8% agar. | pH 5.6, autoclaved at 120 °C, 15 psi for 15 min, cultured at 25 ± 2 °C under 16 h light and 8 h dark photoperiod, light intensity of 1600 Lux. | In vitro propagation. | - | - | [29] |
| Single type cv. Prajwal (Polianthes tuberosa L.) | Bulb | Few drops of Tween-20 or Teepol (0.05%) followed by Bavistin (0.4%) for 15 min followed by HgCl2 (0.1%) for 5–7 min. | MS | BAP (0.5, 1.0, 1.5, 2.0 mg L−1) and NAA (0.5, 1.0, 1.5, 2.0 mg L−1), 2.0 mg L−1 BAP + 0.5 mg L−1 NAA (higher explants regeneration), 0.5 mg L−1 BAP + 2.0 mg L−1 NAA (highest SI and SEL, number of leaves and number of roots in regenerated shoots). | 0.8% (w/v) agar, pH 5.7 autoclaved at 121 °C for 15 min. | Effect of different conc. of BAP and constant, NAA concentration on shoot regeneration of tuberose, effect of different, rooting and acclimatization concentrations of NAA and constant. | - | - | After four weeks of acclimatization, the plants grew vigorously and 80% survived in a pot containing garden soil and sand, after a few weeks hardend plants were transferred to the field with 100% survival rate. | [30] |
| Single and Double (Polianthes tuberosa L.) | Bulbs and shoot buds | 0 1% HgCl2 for 15 min. | MS | Varied conc. Of different PGRs, such as 2, 4-D, BA and IBA (4 mg L−1 2,4 D + 0.5 mg L−1 BA, 0.5 mg L−1 IBA). | 8 g/L agar. | pH 5.7, autoclaved at 121 °C with 105 Kg/Sq cm pressure for 15 min., cultured at-25 ± 1° C at 55% RH and 16/8 photoperiod. | In vitro propagation. | - | - | [32] |
| Single (Polianthes tuberosa L.) | Bulb | 0 1% HgCl2 for 10 min. | MS | Varied concentrations of two auxins 2,4-D 0.5 mg L-1 + NAA 0.75 mg L-1 (SI), NAA (0.25, 0.5) mg L-1 (RI). | 3% bacto agar. | pH 5 7, cultured at 25 ± 1 °C and 55% relative humidity, light-5000-lux. | Photo-morphogenetic effect on callus initiation. | - | - | [33] |
| Unknown (Polianthes tuberosa L.) | Scale stem sections from bulbs were found to be the ideal explant for the enhanced release of axillary buds. Inflorescence segments containing immature flower buds were ideal for somatic organogenesis, whether direct or callus mediated | Bavistin 0.1% for 30 min, followed by ethyl alcohol 50% for 3 min and mercuric chloride 0.10% for 30 min for scale stem sections. For inflorescence segments treatment with 0.10% mercuric chloride for 10 min alone was enough. | MS | BAP 6.0 mg L−1 + KIN 4.0 mg L−1 (bud growth and multiple axillary bud prod.), ½ MS (SEL), 4.0 mg L-1 IBA + 0.2% activated charcoal (SI to RI), 0.2 mg L−1 NAA + 2.0 mg L−1 BAP + 1.0 − 3.0 mg/L KIN (ommature inflorescence to direct organogenesis), 15.0 to 20.0 mg L−1 NAA + 10.0 mg L−1 adenine sulphate (Max. callus index). | - | Shoot induction, multiple shooting, root induction, effect of Gy radiation. | - | Gamma rays at 15 Gy and 20 Gy as well as EMS at 1.0 and 2.0 per cent, were most effective. | - | [34] |
| Prajwal and Suvasini (Polianthes tuberosa L.) | Different plant parts viz. axillary buds, nodal segments of rachis, stem scale, immature flower buds, and petal segments | Bavistin (0.3%) and 8-HQC (0.4%) along with Teepol (2–3 mL) for 3 h followed by 70% ethanol for 1 min and by 0.5% HgCl2 for 2 min and NaOCl (0.25%) for 10 min. | MS | 4.0 mg L−1 BAP + 0.5 mg L−1 NAA (axillary bud proliferation), 4.0 mg L−1 BAP + 0.5 mg L−1 NAA + 0.5 mg L−1 TDZ (SE), 3.0 mg L−1 IBA + 1 mg L−1 NAA (RI). | Hardening in pot mix + cocopeat at 90% RH, 28 ± 2 °C. | Shoot regeneration, multiple shoots, rooting. | - | - | [35] | |
| Prajwal, Phule Rajani, Calcutta single, Calcutta Double, Hyderabad Double, Shringar, Bidhanujwal and Pearl Double (Polianthes tuberosa L.) | Shoot tip culture | Tubers were treated with 1% Bavistin containing 0.1 Tween® 20 overnight followed by 0.1% HgCl2 for 1–2 min. | MS | 4 mg L−1 BAP + 0.1 mg L−1 NAA (tubers to SM), 1.0 mg L−1 (2, 4D + NAA) + 0.5 mg L−1 BAP (CI), ½ MS + 0.5 mg L−1 IBA (shoot to RI). | 30 g/L sucrose, 8 g/L agar. | pH 5.8, temp. 25 ± 2 °C and photoperiod: 16 h light and 8 h dark. | Nematode infection assay to detect nematodes in tuberose roots, callus induction, regeneration, rooting, acclimatization. | - | - | [36] |
| Unknown | Buds | 70% (v/v) ethanol for fews econds followed by immersing in 0.1% HgCl2 solution for 4–5 min followed by several washings. | MS | 1.0 mg L−1 Kin (SI), 1.0 mg L−1 IAA (RI). | - | - | Multiple shoots from callus, multiple shoots from floral axis buds, multiple shoot formation from bulb explants. | Effect of KIN on in vitro shoots initiation in tuberose. Effect of IAA on in vitro root initiation in tuberose. | - | [38] |
| Tuberose (Polianthes tuberosa L.) | Outer and inner scales of bulb | Detergents for 5 min followed by 96% ethyl alcohol followed 0.1% HgCl2 for 2–3 min followed by 20% NaOCl for 15 min. | MS | Different conc. of BAP, NAA and TDZ,3.0 mg L−1 BAP,18 μM TDZ (SI), 2.5 mg L−1, BAP + 0.5 mg L−1 NAA + 0.1 mg L−1 Kin (SM), 1 mg L−1 NAA (RI), 3.0 mg L−1 BAP or 2.5 mg L−1 BAP + 0.5 mg L−1 NAA (BF). | 3% sucrose, Phytagel (1.5 gm/L). | pH 5.5–5.8, autoclaved at 121 °C for 20 min., cultures were maintained at 22 ± 2 °C under 16 h light and 8 h dark photoperiod with a light intensity of 2000 Lux. | Shoot initiation and multiplication, in vitro rooting of shoots, bulblet formation, acclimatization. | After four weeks of acclimatization, the plants grew vigorously and 81% survived in sand. | - | [39] |
| cv. Rajat Rekha (Polianthes tuberosa L.) | Leaves | 70% ethanol for 30 s followed by 0.1% HgCl2 for 2 min. | MS | 1 mg L−1 BA + 15 mg L−1 GA3 or 5 mg L−1 AgNO3 (IC), 0.05 mg L-1−1 BA + 0.1 mg L−1 ABA + 5 mg L−1 GA3 + 0.5 mg L−1 IAA (SM). | 3% sucrose, 0.8% agar. | pH 5.6, autoclaved at 121 °C for 15 min, cultured at- 25 + 1 °C for 16 h photoperiod. | Regeneration of plants. | - | - | [41] |
| Unknown (Polianthes tuberose L.) | Suckers | 5% sodium NaOCl for 10 min followed by 0.1% HgCl2 for 10 min. | - | 0.5 mg L−1 BAP (SI), 0.5 mg L−1 BAP + 0.5 mg L−1 Kin + 0.8 mg L−1 IAA and 2.0 mg L−1 2,4-D + 0.5 mg L−1 BAP + 0.5 mg L−1 Kin + 0.4 mg L−1 IAA (SM), 1.0 mg L−1 2,4-D + 0.5 mg/L IBA (RI). | 30 g/L sucrose, 2.5 g/L Phytagel, pH 5.8. | Shoots were exposed to UV light in laminar air flow for different time interval (5, 10, 15, 20, and 25 min). | The marked differences in the morphological characters were observed with a UV radiation dose of 25 minutes. | Effect of UV radiations on the morphological characters, culture initiation, shoot multiplication, rooting and hardening. | - | [42] |
| Unknown | Axillary buds | 70% alcohol for 20 min followed by 20 % (v/v) clorox for 15 min followed by 5% (v/v) clorox for 10 min. | MS | Various combination of BA (0, 0.1, 0.5, 1.0 and 2.0 mg/L and NAA (0, 0.1, 0.5, 1.0 and 2.0 mg L−1), 0.5 and 1.0 mg L−1 BA (callus), 0.5 mg L−1 BA and 2.0 mg L−1 NAA (C to SM and RI). | 3% sucrose, Oryzalin with high concentrations (more than 1.0 and 2.0 mg/L) and long periods (4, 5, and 6 days) (callus turned brown). | Incubated at 25 ± 2 °C under 16 h photoperiod with illumination provided at 60 μmol m−2 s−1. | Callus induction, multiple shoots induction, effect of oryzalin. | - | - | [43] |
| Four genotypes of tuberose viz: Phule Rajni, Shringar, Prajwal, and Mexican Single (Polianthes tuberosa L.) | Stem disc | 90% (v/v) ethanol for 30 s followed by a treatment of i.e., HgCl2 or Ca(OCl)2 (alone) in different concentrations as well as in combinations of HgCl2 or Ca(OCl)2 with Bavistin at initial vacuum of 100 psi. | MS, WH (White, 1963), and B5 (Gamborg et al., 1968) | WH.3Td.5N (WH) + 0.3 mg L−1 TDZ + 0.5 mg L−1 NAA (SI & SM), WH3N.5Td (WH) + 3.0 mg L−1 NAA + 0.5 mg L−1 TDZ (SEL), WH + 2.0 mg L−1 IBA (RI). | 20 g/L sucrose, pH 5.8 ± 0.1, agar powder at 7.5 g/L. | - | Effects of different surface sterilizing and antifungal agents on recovery of aseptic culture (Ca(OCl)2, HgCl2, Bavistin + Ca(OCl)2, Bavistin+ HgCl2], plant regeneration, multiple shoots, rooted shootlets, regnerants transferred in environmental growth chamber, regnerants transferred in net house, regnerants transferred in field. | - | Regenerated plants were found phenotypically normal and true to the type. | [44] |
| Unknown (Polianthes tuberosa L.) | Lateral bulbs from irradiated tubers | Immersed 3 h in Tecto 60® (Thiabendazole 60% a. i.) (1 g/L), Switch® (Cyprodinil Fludioxonil 37.5 and 25% a. i.) (1 g/L) and Cercobin M® (70% Methyl Thiophanate a. i.) (2 g/L). | MS | 1.0 mg L−1 BA, 0.5 mg L−1 NAA. | 1 g/L Switch® Fungicide. | Photoperiod of 16 h/8 h light/dark and a photosynthetically active radiation of 45 μE m−2 s−1. | Gy irradiation to tubers, in vitro culture of lateral bulbs, tuber’s culture in greenhouses. | 5 Gy showed a shoots’ growth equal than the control’s treatment (100%) and this variable decreased when radiation doses increased (also showed higher explants contamination) | - | [45] |
| Unknown (Polianthes tuberosa L.) | Bulbs of spikenard cultured in Chocholá, Yucatán, México, was selected | NaOCl (3% a.i) found best. | Potato-dextrose agar (PDA) (planting), MS (effect of auxin/cytokinin combination in organogenesis) | Diff. combinations of BA (0,7, 13, 22 µM) and NAA (0, 2.5, 4.5), 7.0 µM BA alone (Best SI). | 3% sucrose, 0.25% Gelrite, pH-5.8, autoclaved at 121 °C for 15 min, incubated at 27 ± 2 °C and 40 µmoles- m−2 s−1 of light. | In vitro formation of adventitious shoots. | - | - | - | [46] |
| Prajwal and Suvasini (Polianthes tuberosa L.) | Axillary buds scooped out from bulbs | Cefotaxime (900 ppm) and 70% ethanol has been found effective. | MS media | 4 mg L−1 BAP + 1 mg L−1 NAA. | - | - | Can in vitro contaminated culture be revived? | - | - | [47] |
| Prajwal and Shringar (Polianthes tuberosa L.) | Bulb | - | MS | Prajwal: 2.0 mg L−1 each (BAP + Kin) (SI), 0.5 mg L−1 IBA + 2.0 mg L−1 IAA (RI), Shringar: 0.2 mg L−1 (BAP + Kin) (SI), 0.5 mg L−1 IBA + 2.5 mg L−1 IAA (RI). | 0.8% agar. | pH 5.8, autoclaved at 120 °C for 20 min and cultured at-16 h light/8 h dark cycle at 2000-lux at 25 ± 2 °C) and 60% RH. | Effect of BAP and Kn, effect of IBA and IAA. | - | - | [48] |
| Unknown (Polianthes tuberosa L.) | Base of the bulb scales, leaf, petals, and stems | - | MS | 1.0 mg L−1 NAA + 0.5 mg L−1 NAA + 4.0 mg L−1 L BAP (SI). | 2% sucrose. | pH 5.7, autoclaved at 1.05 kg/cm2, 121 °C for 20 min, cultures incubated at 25 ± 1 °C, 1200 Lux, photoperiod-16/8 h. | Shot induction, multiple shoots, length of shoot, number of leaves. | - | - | [54] |
| Unknown (Polianthes tuberosa L.) | Vegetative buds of the inflorescence and from tissue corm | - | MS | Diff. conc. BAP, NAA, 2,4-D, IAA, and Kinetin. 4.5 mg L−1 BAP + 0.1 mg L−1 NAA (SI). | 50 ml/L of coconut water, 20 g/L sucrose, 6.4 g/Lagar, pH-5.7. | - | Regeneration in vitro of shoots. | - | - | [55] |
| Polianthes tuberosa L. | Leaf and button explants | - | MS | 4.5 mg L−1 BAP and 0.05 mg L−1 NAA(CI), 4.5 mg L−1 BAP and 0.1 mg L−1 NAA (SI, RI). | 20 g/L sucrose, 6.4 g/L agar. | pH 5.7 | - | - | [56] | |
| Calcutta Single (Polianthes tuberosa L.) | Bulbs | HgCl2 (0.1%) for 2 min and rinsed 3–4 times in distilled water. | MS | 0.2 mg L−1 each (BAP + Kin) (SI), 0.5 mg L−1 (IAA + BAP) (RI). | - | Cultured at 16 h light/8 h dark cycle at 2500–3000 lux at 25 ± 4 °C) and 45–50% RH. | Effect of BAP and Kinetin on shoot development proliferation BAP with IAA for root development. | - | Plantlets kept in garden soil and leaf mold (3:1) for 30 days at high humidity (99%) in hardening room, after that 70% humidity maintained for 30 days followed by 2 months kept in a greenhouse before transplanting in the field. | [58] |
| Hyderabad Single and Local Double Navsari (Polianthes tuberosa L.) | Bulbs | - | MS | Hyderabad Single: 0.5 mg L−1 (BAP + Kin) (SI), 0.5 mg L−1 BAP + 0.2 mg L−1 IAA (RI), Local Double Navsari: 2.0 mg L−1 (BAP + Kin) (SI), 0.5 mg L−1 BAP + 2.5 mg L−1 IAA (RI). | 0.8% agar. | pH 5.8, autoclaved at 120 °C for 20 min., cultured at-16 h light/8 h dark cycle at 2000-lux at 25 ± 2 °C) and 60% RH. | Effect of BAP and Kn on shoot proliferation BAP with IAA on in vitro rooting. | - | - | [60] |
| Cvs Phule Rajni and CalcuttaDouble (Polianthes tuberosa L.) | Bud | - | MS | Phule Rajni: 1.0 mg/L each (BAP + Kin) (SI), 0.5 mg L−1 each (NAA + IAA) (RI), Calcutta Double: 2.5 mg L−1 (BAP + Kin) (SI), 0.5 mg L−1 NAA + 3.5 mg L−1 IAA (RI). | 0.8% agar. | pH 5.8, autoclaved at 120 °C for 15 min, (16 h light/8 h dark cycle at 2000-lux luminance at 25 ± 2 °C) and 60% RH. | Effect of BAP and Kn on shoot induction, effect of NAA and IAA on root induction. | - | - | [61] |
| Local cultivar (Polianthes tuberosa L.) | Suckers | 5% sodium NaOCl for 10 min. followed by 0.1% HgCl2 for 10 min. | Shoots were exposed to gamma radiations doses of 10, 20, 30, 40, and 50 Gy | 0.5 mg L−1 BAP (SI), 0.5 mg L−1 BAP + 0.5 mg L−1 Kin + 0.8 mg L−1 IAA & 2.0 mg L−1 2,4-D + 0.5 mg L−1 BAP + 0.5 mg L−1 Kin + 0.4 mg L−1 IAA (SM), 1.0 mg/L 2,4-D + 0.5 mg L−1 IBA (RI). | - | Cultured at 26 ± 1 °C under 16 h photoperiod with 70 μ mol m−2 s−1 photon flux. and 80% humidity. | Effect of Gamma radiations on the morphological characters, culture initiation, shoot multiplication, and rooting. | Marked differences in the morphological characters were observed with a gamma radiation dose of 30 Gy. | - | [62] |
| Unknown | Bulb, stem scale section | 70% ethanol for a few seconds and finally chlorine water for 15 min. | MS medium | 0.1 mg L−1 2,4-D (CI), NAA2 mg L−1 (SI and RI). | 20 g/L sugar, 6 g/L agar. | 22–24 °C under light intensity 2000 lux. | Shoot induction and root induction. | - | Plants well acclimatized. | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, M.; Sirohi, U.; Malik, S.; Kumar, S.; Ahirwar, G.K.; Chaudhary, V.; Yadav, M.K.; Singh, J.; Kumar, A.; Pal, V.; et al. Methods and Factors Influencing In Vitro Propagation Efficiency of Ornamental Tuberose (Polianthes Species): A Systematic Review of Recent Developments and Future Prospects. Horticulturae 2022, 8, 998. https://doi.org/10.3390/horticulturae8110998
Kumar M, Sirohi U, Malik S, Kumar S, Ahirwar GK, Chaudhary V, Yadav MK, Singh J, Kumar A, Pal V, et al. Methods and Factors Influencing In Vitro Propagation Efficiency of Ornamental Tuberose (Polianthes Species): A Systematic Review of Recent Developments and Future Prospects. Horticulturae. 2022; 8(11):998. https://doi.org/10.3390/horticulturae8110998
Chicago/Turabian StyleKumar, Mukesh, Ujjwal Sirohi, Sunil Malik, Satendra Kumar, Gaurav Kumar Ahirwar, Veena Chaudhary, Manoj Kumar Yadav, Jitender Singh, Arvind Kumar, Virendra Pal, and et al. 2022. "Methods and Factors Influencing In Vitro Propagation Efficiency of Ornamental Tuberose (Polianthes Species): A Systematic Review of Recent Developments and Future Prospects" Horticulturae 8, no. 11: 998. https://doi.org/10.3390/horticulturae8110998