Abstract
Radish (Raphanus sativus L.) is an important root vegetable crop that is easily infected by various pathogens that result in decreased yield and quality. Nucleotide-binding site and leucine-rich repeat (NBS-LRR) genes play vital roles in resisting pathogen infection in plants. However, the genome-wide characterization and functional roles of NBS-LRR genes remain largely unexplored in radish. Here, a total of 187 RsNBS-LRR genes were identified at the whole-genome level in radish, among which 80 RsNBS-LRR genes were unevenly distributed on nine radish chromosomes. Interestingly, 15 clusters containing 36 RsNBS-LRR genes occurred in eight chromosomes. RNA-Seq data showed that several RsNBS-LRR genes exhibited significant differential expression profiles in different radish tissues. Moreover, a range of cis-acting regulatory elements associated with ABA, MeJA, or SA were identified in the promoter region of some RsNBS-LRR genes. RT-qPCR analysis showed that the expression of a few RsNBS-LRR genes (e.g., RsNBS021 and RsNBS163) was significantly induced under Peronospora parasitica infection and/or ABA treatment, indicating that they might play critical roles in ABA-dependent defense resistance processes. These results could enhance our understanding of the evolutionary relationship of RsNBS-LRR genes and facilitate the genetic manipulation of disease resistance in radish breeding programs.
1. Introduction
Plants are often attacked by various pathogens during the development process, including bacteria, fungi, oomycetes and viruses, resulting in decreased yield and quality. In the evolutionary process, plants have evolved complex defense systems to resist pathogens, such as race-specific resistance and race non-specific resistance [1]. The largest class of R genes contains genes harboring the nucleotide-binding site (NBS) and leucine-rich repeat (LRR) domain [2]. Thus far, more than 300 R genes have been cloned from all plant species, of which more than 60% encode NBS and LRR domains [2,3]. According to the structure of the N-terminal domain, NBS-encoding resistance genes can be divided into two groups, including TNL (TIR-NBS-LRR, a Drosophila Toll/mammalian interleukin-1 receptor domain) and CNL (CC-NBS-LRR, a coiled-coil motif) [4,5]. In addition, members of the CNL group containing LZ (leucine zipper) or RPW8 (Resistance to Powdery Mildew 8) domains are named as non-TIR-NBS-LRR. Comparative studies of five Brassicaceae genomes suggested that both the TNL and CNL genes respond simultaneously to pathogen pressures [6], and the gain/loss of TNL genes is more frequent and drastic in different evolutionary nodes. Previous studies reported that the TIR domain has intrinsic NADase activity, which plays a crucial function in pathogen recognition and immune signal transduction [7,8,9]. The CC structure is a functional domain for many proteins such as structural proteins, kinesins and transcription factors, and it plays an important role in protein interaction [10]. RPW8 contains a putative N-terminal transmembrane domain and a CC motif and exhibits broad-spectrum mildew resistance in Arabidopsis thaliana [11,12]. The LRR domain, harboring a conserved sequence of xxLxLxx, is highly irregular and variable, and is responsible for protein–protein interactions and pathogen identification [13].
Phytohormones, including salicylic acid (SA), and jasmonic acid (JA), play important roles in signal transduction networks of pathogen responses and disease resistance in plants [14]. Accumulation of SA and JA in plant can induce the expression of pathogenesis-related (PR) genes and consequently increase plant disease resistance [15]. Moreover, some other hormones such as gibberellins, abscisic acid (ABA) and ethylene can enhance plant immunity by modulating the SA- or JA-dependent signaling pathways [16]. The cross-talk among different phytohormones has formed complex and efficient signaling pathways for regulating plant defense against pathogen infection [14,15,16]. In addition, several vital transcription factor genes such as WRKYs, ERFs and MYBs are reported to be involved in the regulatory process of disease resistance in plants [17]. These findings have provided useful information to clarify the molecular mechanism underlying NBS-mediated disease resistance in plants.
Currently, the availability of many plant genome databases makes it feasible to identify, locate and characterize R genes at the genome-wide level. To date, the NBS-LRR-encoding gene family has been identified in many plant species, including A. thaliana [5,18], Medicago truncatula [4], Brassica rapa [19], Sorghum bicolor [20], Vitis vinifera [21], Solanum tuberosum [22] and Setaria italica [23]. The NBS-LRR-encoding gene family is highly replicate, diversely evolved and clustered [24], and mainly participates in promoting plant disease resistance. More than 2000 NBS-LRR-encoding genes have been identified in bread wheat [25], whereas less than 100 NBS-LRR-encoding genes have been identified in some species such as cabbage [19], cucumber [26], and S. italic [23]. The great diversity in the number of NBS-LRR-encoding genes indicates the complex evolutionary patterns in different plant genomes. Thus far, several main mechanisms underlying the formation of the novel R gene have been proposed, such as gene duplication, gene recombination, the differences in the intergenic spacer region and the composition of transposable elements [24,26].
Radish (Raphanus sativus L., 2n = 2x = 18) is an important root vegetable crop belonging to the Brassicaceae family [27]. Many radish cultivars and accessions are highly susceptible to downy mildew, root rot and turnip mosaic virus. Peronospora parasitica is a destructive and widespread pathogen that can easily infect radish plants during the seedling and adult-plant stages under humid and warm conditions, which results in low yield and quality and even unmarketable products [28]. The traditional breeding method is ineffective and time-consuming in practice. Adoption of an effective molecular method to identify disease-resistance genes has becoming a practical and effective strategy to improve resistance and develop multi-resistance cultivars [29]. Currently, the available radish genome sequences [30,31,32,33,34] provide a valuable platform for comprehensive identification of the NBS-LRR-encoding gene family at the whole genome level. Although several resistance gene analogues (RGAs) from the radish transcriptome have been identified [35], the genome-wide identification and characterization of NBS-LRR-encoding genes remains largely unexplored in radish. In this study, we aimed to characterize the genetic structure, chromosome localization and genetic evolutionary relationship of the NBS-LRR-encoding gene family in radish and to investigate their expression profiles in different radish tissues and under biotic stress as well as ABA treatments. These findings could provide valuable information for efficient utilization of disease-resistance genes in radish and could facilitate improvement of multiple disease resistance in radish.
2. Materials and Methods
2.1. Plant Materials and Treatments
Two advanced inbred lines of radish, ‘NAU-DHP’ (resistant to downy mildew, DM) and ‘NAU-YZH’ (susceptible to DM), were used in this study. The seeds were grown in sterilized soil at 25 °C/18 °C (day/night) with a photoperiod of 14 h light and 10 h dark. In total, ten one-month-old radish seedlings were infected with P. parasitica isolate, or treated with 100 µM ABA, respectively. The untreated seedlings grown in normal condition were used as controls. Then, the young leaf tissues from three independent individuals were collected at 6, 12, 24, 48, 72 and 96 h after treatments, which were immediately frozen in liquid nitrogen and stored at −80 °C for further use.
2.2. Identification of NBS-LRR-Encoding Genes in Radish Genome
The genomic sequences downloaded from the Radish Genome DataBase (http://radish.kazusa.or.jp/, accessed on 20 December 2021) [30] were used for identification of the NBS-LRR-encoding genes. A set of predicted NBS-LRR proteins was identified from the radish genome using the Hidden Markov Model (HMM) (HMMER3) [36] corresponding to the NBS (NB-ARC; Pfam: PF00931) family. Proteins with an e-value lower than 0.01 were chosen as candidate genes, which were further filtered based on manual curation and functional annotation against the Arabidopsis database (TAIR, http://www.arabidopsis.org/index.jsp, accessed on 20 December 2021). TIR-only (TIR, Pfam: PF01582) and RPW8-only (RPW8, Pfam: PF05659) sequences harboring no NBS domain were also identified.
2.3. Analysis of Gene Structure and Conserved Domains/Motifs
The ProtParam tool of ExPASy (https://web.expasy.org/protparam/, accessed on 20 December 2021) was utilized to analyze the characteristics of RsNBS proteins including molecular weight, theoretical pI (isoelectric point) and instability index. The Gene Structure Display Server (GSDS: http://gsds.cbi.pku.edu.cn/, accessed on 20 December 2021, GSDSS 2.0) [37] was used for gene structure prediction. Conserved domains were identified using Pfam (http://pfam.xfam.org/, accessed on 20 December 2021, version 29.0) [38] and SMART (http://smart.embl-heidelberg.de/, accessed on 20 December 2021, version 7) [39]. Potential motifs of the candidate NBS-LRR protein sequences were identified using MEME (Multiple Expectation Maximization for Motif Elicitation) program (http://meme-suite.org/, accessed on 20 December 2021, version 4.11.2) with a motif width of 50-100 amino acids (aa) and a maximum motif number of 15. The CC motif was identified using the MARCOIL program with a threshold above 90% [40].
2.4. Sequence Alignment and Phylogenetic Analysis
The NBS-encoding resistance proteins were further subdivided into CNL and TNL groups, and the divergence between groups was examined based on phylogeny. The predicted proteins were trimmed at ~10 aa N-terminal to the first Gly before the P-loop motif and ~30 aa beyond the MHDL motif [7], and multiple alignment for these sequences was performed by CLUSTALW with default options. The neighbor-joining (NJ) method was performed to generate a phylogenetic tree by MEGA6.0 with a bootstrap of 1000 replicates.
2.5. Anchoring NBS-LRR-Encoding Genes to the Radish Chromosomes
The chromosomal location analysis of RsNBS-LRR genes was carried out using the Radish Genome Database (http://radish-genome.org/, accessed on 20 December 2021) [32]. Gene sequences with coverage ≥80% and identity ≥92% were considered as identical gene sequences. MapChart Software [41] was used for localizing the RsNBS genes on the chromosomes according to the corresponding location parameters. The Multiple Collinearity Scan toolkit (MCScanX) [42] was adopted to identify RsNBS duplication events with default parameters. The relationships of orthologous RsNBS genes among R.sativus, A. thaliana, B. rapa, C. sativus and O. sativa were plotted using TBtools software [43].
2.6. Promoter Analysis and Transcriptome-Based Expression Profiling of RsNBS-LRR Genes
The 2 kb upstream sequences of the transcription start site (TSS) for the chromosome-located RsNBS-LRR genes were isolated and loaded into the New PLACE (https://www.dna.affrc.go.jp/PLACE/?action=newplace, accessed on 20 December 2021) and PlantPAN (http://plantpan.itps.ncku.edu.tw/promoter.php, accessed on 20 December 2021) database to identify potential cis-acting regulatory elements (CREs) targeted by transcription factors.
The expression profile of the chromosomal-located RsNBS-LRR genes in three tissues (cambium, cortex and parenchyma) and three growth stages (5, 7 and 9 weeks post-seed planting) was investigated using a previous taproot transcriptome [44]. The sequences of RsNBS-LRR genes were converted into identifiable genes ID name using local Blast software with the identity of at least 98%. The FPKM (fragments per kilobase of transcript per million mapped fragments) method was employed to present the expression level of each RsNBS gene. Heat maps were generated using TBtools software [43].
2.7. RNA Isolation and Quantitative Real-Time PCR
Total RNA was extracted from leaves using Trizol reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). The first-strand cDNA was synthesized using a Superscript III First-Strand Synthesis System (Invitrogen). The RT-qPCR reaction and cycling profile were carried out on a LightCycler® 480 System (Roche, Mannheim, Germany). The RsActin gene was selected as the reference gene [29]. Three biological and technical replicates were performed. The data were statistically analyzed with Duncan’s multiple range test at the p < 0.05 level of significance using SPSS 21 software (SPSS Inc., Chicago, IL, USA). The RT-qPCR primer sequences are shown in Table S1.
3. Results
3.1. Characterization of RsNBS-LRR Genes in Radish
Based on an HMM profile search against Radish Genome protein sequences with the NBS domain (Pfam: PF00931), a total of 288 putative NBS-LRR proteins were obtained. To ensure the reliability of the sequences, low-quality sequences without start and/or stop codons were removed. Ultimately, a total of 187 NBS-LRR-encoding genes were identified, which were named as RsNBS001-RsNBS187 (Table S2). Among them, 24 and 79 were classified as CNLs (CC-NBS-LRR) and TNLs (TIR-NBS-LRR) by conserved domains, respectively (Table 1). Additionally, the remaining 84 proteins were classified into four distinct groups, namely CN (24), TN (28), NL (21) and N (11) (Table 1).

Table 1.
The number of identified NBS-LRR genes in radish.
The average number of exons in RsNBS-LRR genes was 4.84. Moreover, the average number of exons in TNL-class genes (5.55 per gene) was higher than that in CNL ones (3.33 per gene). Thirty percent of CNL-class genes were encoded by a single exon, and the rest of the genes had from 2 to 11 exons (Table S2 and Figure S1). The number of exons in most TNL-class genes varied from 2 to 19, except for RsNBS163. The MW (molecular weight) of the RsNBS-LRR proteins was 34.01~311.5 KDa, and the PI ranged from 4.97 to 9.57 (Figure S3A; Table S3). The theoretical PIs of 132 (70.6%) RsNBS proteins were less than 7. The MWs of 171 (91.4%) RsNBS proteins fell into the range from 50 to 200 KDa. The instability index of 160 (85.6%) RsNBS proteins was above 40.0 with an average of 44.69 (Figure S3B; Table S3), indicating that most of the RsNBS-LRR proteins are unstable.
3.2. Sequence Alignment and Gene Structure Analysis
In all, these RsNBS-LRR protein sequences were divided into four groups according to phylogenetic tree order (Figure S1). The LOGO of the amino acid sequence of 15 conserved motifs for the RsNBS-LRR proteins was predicted (Figure S2). All protein sequences contained a highly conserved NBS domain containing several conserved motifs, such as motifs 1, 3, 6, and 7. Among them, motif 3 was a P-loop, motif 7 represented Kinase-2, motif 1 contained three motifs (RNBS-B, RNBS-C and GLPL), and motif 6 was an MHDL motif. Nevertheless, C-terminals with continuous motifs 4, 5 and 10 were considered as a repeated conserved LRR domain (Figure S1 and S2). Moreover, motifs 9 (CC) and 14 (RPW8) were presented at the N-terminal of most CNL-class genes, respectively. The conserved RPW8 domain was identified in some CNL-class proteins (e.g., RsNBS055, RsNBS057, RsNBS074 and RsNBS076), while P-loop, Kinase-2, RNBS-B, RNBS-C, GLPL and MHDL were found in the TNL-class proteins. In addition, eight major motifs (P-loop, RNBS-A, Kinase 2, RNBS-B, RNBS-C, GLPL, RNBS-D and MHDL) were identified among the RsNBS-LRR proteins (Table 2). Protein sequences in the same clade have similar protein structures based on their motif composition (Figure S1C), suggesting that they may have similar biological functions.
Table 2.
Consensus sequences of major NBS domains in radish CNL and TNL proteins.
Both the TIR-only and RPW8-only proteins are important parts of the NLR repeat family. In this study, a total of 54 TIR-only and 134 TIR-X proteins were identified (Table S4), while 107 out of 134 TIR-X proteins were TIR-NBS-encoding proteins (Table 1). Moreover, three RPW8-only proteins and eight RPW8-NBS-encoding proteins were found (Table S4). Phylogenetic analysis showed that all eight RPW8-NBS-encoding proteins were classified as CNL-A. Notably, three RPW8-NBS-encoding genes (RsNBS055, RsNBS057 and RsNBS074) had high similarity with ADR1-like1 in Arabidopsis, while RsNBS062 and RsNBS068 had high similarity with ADR1-like2 and ADR1, respectively (Table S3).
3.3. Phylogenetic Analysis of RsNBS-LRR Proteins
To better understand the phylogenetic relationship of the NBS-encoding genes between radish and A. thaliana, a phylogenetic tree of NBS-encoding resistance genes between radish and A. thaliana (136) was constructed (Figure S4). The radish NBS proteins were divided into two subgroups, clade CNL (60) and clade TNL (127). Phylogenetic trees for CNL-class and TNL-class NBS-encoding genes were generated using A. thaliana and radish proteins, respectively (Figure 1A,B). The TNL-class resistance genes were divided into nine subgroups (Figure 1A), accounting for 67.9% of the total RsNBS genes, namely TNL-A (9), TNL-B (16), TNL-C (2), TNL-D (12), TNL-E (7), TNL-F (7), TNL-G (37), TNL-H (23) and TN (14). Among them, the largest RsNBS gene group was TNL-G (39), and the subgroup TNL-C had the fewest numbers (2). Moreover, 60 CNL-class RsNBS genes were classified into four subgroups (Figure 1B) including CNL-A (10), CNL-B (32), CNL-C (5) and CNL-D (13). CNL-B was the largest subgroup consisted of 32 RsNBS-LRR genes.
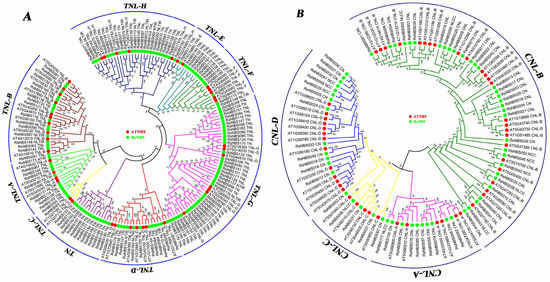
Figure 1.
Phylogenetic relationship of radish and A. thaliana TNL (A) and CNL (B) NBS proteins. Subgroups are marked in different colors.
3.4. Chromosomal Location and Evolutionary Analysis of RsNBS-LRR Genes
In total, 80 of 187 (42.78%) NBS-LRR-encoding genes (24 CNL and 56 TNL) were dispersedly localized to nine radish chromosomes (Figure 2 and Table S5). The R9 contained the largest number of genes (17, 21.25%), followed by 12 (15.00%) genes located in R2 and R4. Interestingly, 15 gene clusters were unevenly distributed on eight radish chromosomes (Figure 2). In detail, R2 contained four gene clusters, and each cluster contained two genes. Some members in one gene cluster came from the same subfamily. RsNBS140 and RsNBS157 in R2 belonged to the TN subfamily, while RsNBS071, RsNBS099 and RsNBS124 in R9 belonged to the TNL-H subfamily. However, some other genes in one gene cluster belonged to different subfamilies. In R7, RsNBS107 and RsNBS155 belonged to the TNL-D subfamily, while RsNBS096 and RsNBS092 belonged to the TN and TNL-H subfamily, respectively.
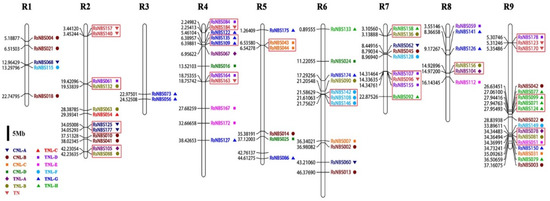
Figure 2.
Distribution of the RsNBS-LRR genes on radish chromosomes (R1–R9).
Additionally, ten pairs of whole genome duplication (WGD) genes (e.g., RsNBS004-RsNBS014 and RsNBS157-RsNBS096) were identified in all chromosomes except for R3 (Figure 3 and Table S6). Five RsNBS-LRR genes located at R9 experienced a WGD event (Figure 3 and Table S6).
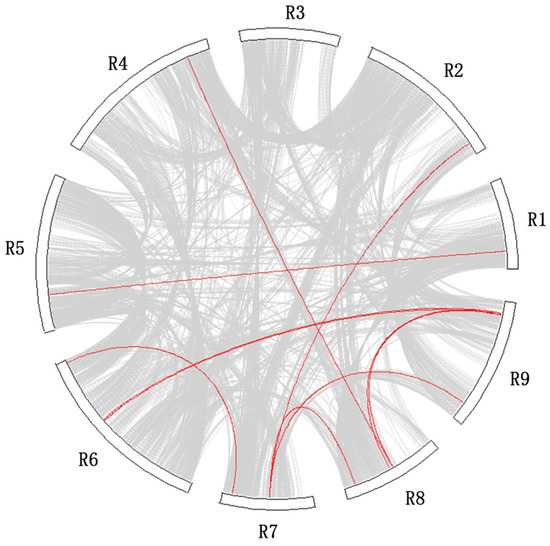
Figure 3.
The interchromosomal relationships of RsNBS-LRR genes. Red and grey lines indicate duplicated NBS-LRR-encoding gene pairs and synteny blocks, respectively.
To further explore the evolutionary origin of the radish NBS-LRR-encoding gene family, three dicotyledonous plants (A. thaliana, B. rapa and Cucumis sativus) and a monocotyledonous plant, Oryza sativa, were used to construct syntenic maps (Figure 4). A great number of RsNBS-LRR genes have a syntenic relationship with the reference plants, such as 54 genes in B. rapa, 43 in A. thaliana, 8 in C. sativus and 3 in O. sativa. RsNBS060 has homology to four reference plants. Syntenic analysis revealed that 21 RsNBS-LRR genes were associated with at least three syntenic gene pairs in B. rapa, and the highest number of associations was 15. These results showed that a large proportion of RsNBS-LRR genes are closely related to their homologous genes in Arabidopsis and B. rapa.
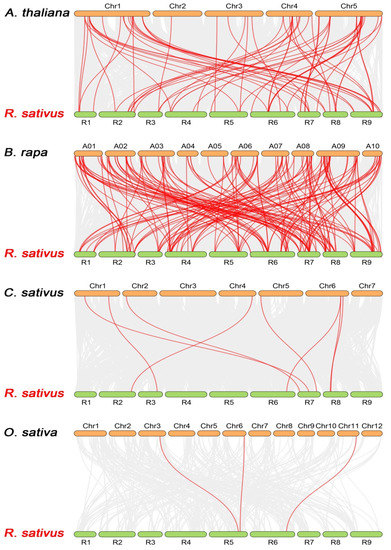
Figure 4.
Synteny analysis of NBS-LRR-encoding genes between radish and four representative plant species. Grey lines in the background indicate collinear blocks within radish and other plant genomes, while red lines highlight the syntenic NBS-LRR-encoding gene pairs.
3.5. Potential CREs Identification from Promoter Region of RsNBS-LRR Genes
To investigate potential regulatory networks of the 80 chromosome-located RsNBS-LRR genes, the CREs were analyzed from 2 kb upstream sequences of the TSS using both the New PLACE and PlantPAN database. Considering that the ABA, MeJA, and SA were vital signal molecules involved in regulatory pathways of plant responses to pathogen infections, we focused on the potential CREs involved in the responsiveness of these three phytohormones. The ABREs (e.g., CACGTG, TACGGTC or AACCCGG) were found in the promoter region of 65 RsNBS-LRR genes, including 22 CNLs and 43 TNLs (Table S7). Moreover, the CGTCA/TGACG-motif and TCA-element (TCAGAAGAGG or CCATCTTTTT) were found in the promoter region of 62 RsNBS-LRR genes (19 CNLs and 43 TNLs) and 39 RsNBS-LRR genes (14 CNLs and 25 TNLs), respectively (Table S7). The results indicated that these RsNBS-LRR genes harboring the ABREs, CGTCA/TGACG-motif or TCA-element might be crucial participators in ABA, MeJA, or SA-dependent defense resistance processes, respectively.
3.6. Differential Expression Profiles of RsNBS-LRR Genes in Radish Tissues
The FPKM values from three tissues (cambium, cortex and parenchyma) and three growth stages (5, 7, and 9 weeks post-seed planting) were used to estimate the expression levels of 80 chromosome-located RsNBS-LRR genes (Figure 5). A proportion of RsNBS-LRR genes (e.g., RsNBS007, RsNBS016, RsNBS099 and RsNBS124) was expressed in almost each tissue and developmental stage (Figure 5). However, a few RsNBS-LRR genes such as RsNBS024, RsNBS025, RsNBS078 and RsNBS079 exhibited low expression levels in all tissue and growth stages. Several RsNBS-LRR genes exhibited tissue-specific expression in radish. For example, RsNBS022 was only expressed in parenchyma at the 9-week-old stage, while RsNBS018 and RsNBS177 were specially highly expressed in cambium at the 5-, 7- and 9-week-old stage (Figure 5), indicating that they may play crucial roles in tissue-specific disease resistance in radish.
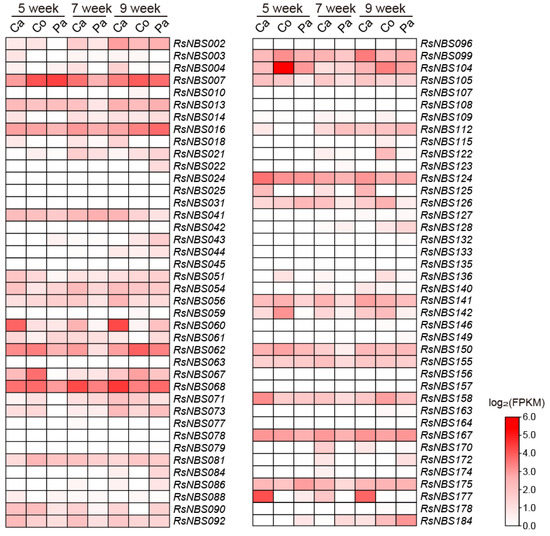
Figure 5.
Transcriptome-based expression profiling of RsNBS-LRR genes from different tissues and growth stages (5, 7, and 9 weeks post-seed planting). Ca, cambium; Co, cortex; Pa, parenchyma.
3.7. Expression Profiling of RsNBS-LRR Genes under Biotic and Abiotic Stresses
RT-qPCR analysis showed that some RsNBS-LRR genes exhibited differential expression patterns under P. parasitica infection or ABA treatments (Figure 6). In general, the RsNBS-LRR genes had a relatively higher expression level in the ‘NAU-DHP’ genotype than in the ‘NAU-YZH’ genotype (Table S8). After the plants were infected with P. parasitica isolate, the expressions of the RsNBS044 and RsNBS163 genes were upregulated to 28.18- and 27.09-fold higher than the control at 12 h in the ‘NAU-DHP’ genotype, respectively (Figure 6A). The expression level of the RsNBS021 and RsNBS062 genes peaked at 12 h at 12.64- and 13.94-fold higher than the control, respectively (Figure 6A). In the ‘NAU-YZH’ genotype, the RsNBS021 and RsNBS044 genes had the highest expression levels, at 1.83-fold higher than the control at 12 h and 1.90-fold higher at 24 h, respectively (Figure 6B).
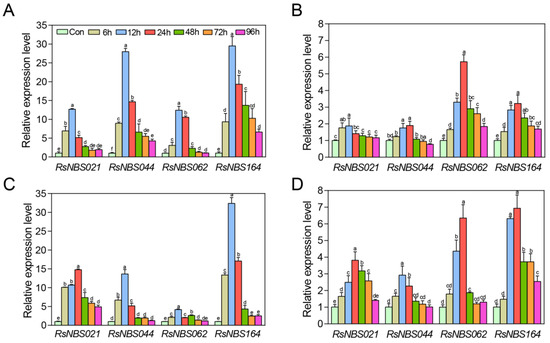
Figure 6.
Expression profiles of RsNBS-LRR genes under Peronospora parasitica infection or ABA treatments. (A), (B): The radish genotype ‘NAU-DHP’ (A) and ‘NAU-YZH’ (B) plants were infected with P. parasitica. (C), (D): The radish genotype ‘NAU-DHP’ (C) and ‘NAU-YZH’ (D) plants were treated with ABA. Con, control. Data are means ± SD. Different letters above the bars indicate significant differences (p < 0.05) according to Duncan’s test.
The expression level of these four RsNBS-LRR genes was significantly induced under the ABA treatment. RsNBS163 gene exhibited maximum expression level at 12 and 24 h in ‘NAU-DHP’ (C) and ‘NAU-YZH’, respectively (Figure 6C,D). The RsNBS021 gene peaked at 12 h at 14.68-fold and 3.65-fold higher than the control in ‘NAU-DHP’ and ‘NAU-YZH’, respectively. The results indicated that several RsNBS-LRR genes might be involved in disease resistance processes in an ABA-dependent manner.
4. Discussion
Radish plants are inevitably affected by many kinds of pathogens, especially under unfavorable environmental conditions. Identification of disease-resistance genes is a practical and useful approach to provide knowledge about multi-disease resistance in radish. In this study, a total of 187 RsNBS-LRR genes were identified, which were divided into two subfamilies (TNL and CNL) and 13 subgroups according to the different domains. The ratio of TNL to CNL genes in radish is approximately 2:1, which is similar to that in B. rapa and Arabidopsis [5,17]. The RsNBS-LRRs belonging to the TNL subfamily was larger than that in the CNL subfamily, indicating that TNL resistance genes had an earlier origin than CNL genes [3,7]. Moreover, the distribution of the RsNBS-LRR genes was dispersedly clustered on nine radish chromosomes, which is in accordance with that in some other plants species, including M. truncatula, B. rapa and C. sativus [5,19,26]. The irregular distribution of NBS-LRR-encoding genes may be partially attributed to the long-term evolution of plant adaptation to living environmental conditions. Previous studies revealed that the resistance gene cluster was generated by gene duplication and recombination in the evolution process, and a larger number of members in a gene cluster represents more gene duplications [19]. In the current study, a gene cluster containing four NBS-LRR-encoding genes is located in R7, indicating that R7 may have undergone large-scale replication of the NBS-LRR-encoding genes. Frequent gene duplication can increase the number of homologous genes and gene cluster members. Homologous genes promote intragenic and intergenic recombination, which can produce new resistance genes to cope with emerging pathogens [45]. RsNBS107 is associated with 15 syntenic gene pairs in B. rapa, suggesting that it might had a crucial role in RsNBS-LRR gene evolution.
Resistance genes containing an RPW8 domain belong to the powdery mildew resistance gene family, and they play critical roles in resistant to powdery mildew pathogens in Arabidopsis [12]. In the current study, a total of eight RPW8-NBS-encoding genes belonging to the CNL-A subfamily were identified (Table S4). Increasing evidence demonstrates that RPW8-NBS proteins could act as ‘helpers’, which are required in pathogen response and disease resistance in Arabidopsis [46,47]. Five RPW8-NBS-encoding genes (RsNBS055, RsNBS057, RsNBS062, RsNBS068, and RsNBS074) had high similarity with the AtADR1 genes in this study. Considering that the ADR1 protein can induce cell death and trigger effector-triggered immunity (ETI) in Arabidopsis [48], it is reasonable to indicate that these RPW8-NBS-encoding genes might play a vital role in the defense response process in radish.
Recently, a number of RsNBS genes have been identified from two other radish genomes [33,49]. A total of 101 and 111 RsNBS-LRR genes identified in this study were highly homologous with the RsNBS genes from the Xin-li-mei and WK10039 radish genome (Table S2; identity > 92% and p value < 1 × 10−20), respectively. The results indicated that a large proportion of RsNBS genes was highly conserved among different radish genotypes and may play crucial roles in disease resistance in radish. In this study, the expression of a few RsNBS genes was significantly induced in resistant genotype ‘NAU-DHP’ after P. parasitica inoculation (Figure 6), indicating their potential application for improving radish resistance to DM pathogen invasion. In a previous study, RNA-seq and RT-qPCR analysis showed that six NBS-encoding genes (RsN25.2, RsTNL03, RsNL07, RsTN18, RsTNL09, and RsTN17) might positively regulate radish resistance to Fusarium oxysporum [49,50]. The RsNBS080, RsNBS088 and RsNBS109 genes identified in this study were highly homologous with RsNL07, RsTNL09 and RsTN17 (Table S2; identity > 92% and p value < 1 × 10−20), respectively. The results indicated that these three RsNBS-LRR genes may play crucial roles in radish resistance to F. oxysporum. Moreover, the previously reported RsCNL02, RsCNL06, RsRN05 and RsTN19 genes [49] had high identities to the P. parasitica-induced RsNBS021, RsNBS044, RsNBS062 and RsNBS163, indicating that they might also be involved in the P. parasitica resistance process. Further characterization of these RsNBS genes using a segregated population would facilitate investigating the association between the candidate RsNBS genes and the phenotype, and contribute to clarify the molecular mechanism underlying RsNBS-mediated disease resistance in radish.
ABA is a crucial phytohormone, and it acts as an important signaling molecule in plant defense pathways [15]. In this study, several RsNBS-LRR genes (e.g., RsNBS021 and RsNBS163) exhibited significantly increased expression levels under ABA treatment, and the ABREs (e.g., CACGTG, TACGGTC or AACCCGG) were found in their promoter region. In Arabidopsis, several R genes including RPP1, RPP2, RPP5, RPP7, RPP8 and RPP13 were validated as resistance genes, which could recognize the DM pathogen and induce a hypersensitivity response (HR) [17]. Considering that the exogenous ABA treatment can trigger a series of defense responses to disease, it is reasonable to infer that these ABA-induced RsNBS-LRR genes might play a vital role in the ABA-mediated disease resistance process in radish.
5. Conclusions
In summary, this study represents a systematic report on genome-wide identification and characterization of the NBS-LRR-encoding gene family in radish. In all, 80 of these identified 187 RsNBS-LRR genes were unevenly distributed on nine radish chromosomes. A total of 15 gene clusters containing 36 RsNBS-LRR genes occurred in eight chromosomes. Several CREs that are responsive to ABA, MeJA, or SA were identified in the promoter region of a number of RsNBS-LRR genes. Besides exhibiting differential tissue-specific expression profiles, the expression level of a few RsNBS-LRR genes (e.g., RsNBS021 and RsNBS163) was significantly induced under P. parasitica infection and/or ABA treatment, indicating that they may be involved in phytohormone-triggered defense resistance processes. The outcomes of this study provide insights into the evolutional characterization of RsNBS-LRR genes and contribute to genetic improvement of multiple-disease resistance in a radish breeding program.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8121164/s1, Table S1: The primer sequences used for RT-qPCR analysis in this study; Table S2: Classification of RsNBS-LRR genes based on the NBS domain structure; Table S3: The detailed information of 187 RsNBS-LRR genes in radish; Table S4: The detailed information of TIR-only and RPW8-only proteins in radish; Table S5: The detailed information of chromosomal localization of RsNBS-LRR genes; Table S6: The interchromosomal relationships of RsNBS-LRR genes; Table S7: Characterization of potential CREs associated with ABA, MeJA and SA from promoter region of the chromosome-located RsNBS-LRR genes; Table S8: The expression data of RsNBS-LRR genes under Peronospora parasitica infection or ABA treatments; Figure S1: The phylogenetic analysis of RsNBS proteins and gene structures. (A) Phylogenetic tree of RsNBS proteins; (B) intron–exon structure distribution of RsNBS genes; (C) conserved motif distribution of RsNBS proteins. (a) The motif of clade CNL-A, CNL-B, CNL-C, CNL-D; (b) the motif of clade TNL-E, TNL-F, TNL-H; (c) the motif of clade TNL-D, TNL-G; (d) the motif of clade TNL-A, TNL-B, TNL-C,TN; Figure S2: The LOGO of 15 amino acid motifs in four parts; Figure S3: The physical and chemical properties of RsNBS proteins. (A) The distribution of MWs and PIs; (B) The distribution of aliphatic index, instability index and Grand Average of hydropathicity (GRAVY); Figure S4: Phylogenetic tree of RsNBS and AtNBS proteins.
Author Contributions
Conceptualization, L.X. and L.L.; writing—original draft preparation, L.X. and W.Z.; methodology, W.Z. and M.T.; data curation and software, M.T. and X.Z.; writing—review and editing, X.Z., J.W. and Y.W.; project administration, Y.W. and L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was in part supported by grants from the Jiangsu Agricultural Science and Technology Innovation Fund (CX(21)2020), National Key Technology Research and Development Program of China (2018YFD1000800), Fundamental Research Funds for the Central Universities (KYCYXT2022005; KYYZ202108), and Jiangsu Agricultural Industry Technology System (JATS [2022]463).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Y.-M.; Chen, M.; Sun, L.; Wang, Y.; Yin, J.; Liu, J.; Sun, X.-Q.; Huang, Y.-Y. Genome-wide identification and evolutionary analysis of NBS-LRR genes from Dioscorea Rotundata. Front. Genet. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Hamblin, M.-T.; Prochnik, S.; Jannink, J.-L. Identification and distribution of the NBS-LRR gene family in the Cassava genome. BMC Genom. 2015, 16, 360. [Google Scholar] [CrossRef] [PubMed]
- Ameline-Torregrosa, C.; Wang, B.-B.; O’Bleness, M.-S.; Deshpande, S.; Zhu, H.; Roe, B.; Young, N.-D.; Cannon, S.-B. Identification and characterization of nucleotide-binding site-leucine-rich repeat genes in the model plant Medicago truncatula. Plant Physiol. 2008, 146, 5–21. [Google Scholar] [CrossRef]
- Meyers, B.-C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.-W. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 2003, 15, 1683. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Shao, Z.-Q.; Wang, Q.; Hang, Y.-Y.; Xue, J.-Y.; Wang, B.; Chen, J.-Q. Uncovering the dynamic evolution of nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in Brassicaceae. J. Integr. Plant Biol. 2016, 58, 165–177. [Google Scholar] [CrossRef]
- Meyers, B.-C.; Morgante, M.; Michelmore, R.-W. TIR-X and TIR-NBS proteins: Two new families related to disease resistance TIR-NBS-LRR proteins encoded in Arabidopsis and other plant genomes. Plant J. 2002, 32, 77–92. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, Z.; Zhang, Y.-M.; Li, Q.; Jiang, X.-M.; Jiang, Z.; Tang, J.-H.; Chen, D.; Wang, Q.; Chen, J.-Q.; et al. An angiosperm NLR atlas reveals that NLR gene reduction is associated with ecological specialization and signal transduction component deletion. Mol. Plant 2021, 14, 2015–2031. [Google Scholar] [CrossRef]
- Goldner, R.; Yaron, A. TIR axons apart: Unpredicted NADase controls axonal degeneration. Neuron 2017, 93, 1239–1241. [Google Scholar] [CrossRef]
- Liu, J.-L.; Liu, X.-L.; Dai, L.-Y.; Wang, G.-L. Recent progress in elucidating the structure, function and evolution of disease resistance genes in plants. J Genet Genom. 2007, 34, 765–776. [Google Scholar] [CrossRef]
- Wang, W.; Wen, Y.; Berkey, R.; Xiao, S. Specific targeting of the Arabidopsis resistance protein RPW8.2 to the interfacial membrane encasing the fungal haustorium renders broad-spectrum resistance to powdery mildew. Plant Cell 2009, 21, 2898–2913. [Google Scholar] [CrossRef]
- Xiao, S.; Calis, O.; Patrick, E.; Zhang, G.; Charoenwattana, P.; Muskett, P.; Parker, J.-E.; Turner, J.-G. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant J. 2005, 42, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.-B.; Bogdanove, A.-J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X. Salicylic acid: Biosynthesis, perception, and contributions to plant immunity. Curr. Opin. Plant Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, A.M.; Argueso, C.T. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef]
- Wang, D.; Jiang, C.; Liu, W.; Wang, Y.J. The WRKY53 transcription factor enhances stilbene synthesis and disease resistance by interacting with MYB14 and MYB15 in Chinese wild grape. J. Exp. Bot. 2020, 71, 3211–3226. [Google Scholar] [CrossRef]
- Van de Weyer, A.-L.; Freddy, M.; Oliver, J.-F.; Marc, T.-N.; Volkan, C.; Kamil, W.; Jonathan, D.-G.-J.; Jeffery, L.-D.; Detlef, W.; Felix, B. A species-wide inventory of NLR genes and alleles in Arabidopsis thaliana. Cell 2019, 178, 1260–1272. [Google Scholar]
- Mun, J.-H.; Yu, H.-J.; Park, S.; Park, B.-S. Genome-wide identification of NBS-encoding resistance genes in Brassica rapa. Mol Genet. Genomics 2009, 282, 617–631. [Google Scholar] [CrossRef]
- Cheng, X.; Jiang, H.-Y.; Zhao, Y.; Qian, Y.-X.; Zhu, S.-W.; Cheng, B.-J. A genomic analysis of disease-resistance genes encoding nucleotide binding sites in Sorghum bicolor. Genet. Mol. Biol. 2010, 33, 292–297. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, X.; Yue, J.-X.; Tian, D.; Chen, J.-Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genom. 2008, 280, 187–198. [Google Scholar] [CrossRef]
- Prakash, C.; Trognitz, F.C.; Venhuizen, P.; Haeseler, A.; Trognitz, B. A compendium of genome-wide sequence reads from NBS (nucleotide binding site) domains of resistance genes in the common potato. Sci. Rep. 2020, 10, 11392. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Weng, Q.-Y.; Song, J.-H.; Ma, H.-L.; Yuan, J.-C.; Dong, Z.-P.; Liu, Y.-H. Bioinformatics analysis of NBS-LRR encoding resistance genes in Setaria italic. Biochem. Genet. 2016, 54, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.-X.; Liu, J.-G.; Wu, C.-F.; Deng, Y.-S.; Cai, C.-W.; Zhang, X.; Cai, Y.-F. Genome-wide comparative analysis of NBS-encoding genes in four Gossypium species. BMC Genom. 2017, 18, 292. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Si, W.; Zhao, L.; Yang, S.; Zhang, X. Dynamic evolution of NBS–LRR genes in bread wheat and its progenitors. Mol. Genet. Genom. 2015, 290, 727–738. [Google Scholar] [CrossRef]
- Zhang, W.; Yuan, Q.; Wu, Y.; Zhang, J.; Nie, J. Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2022, 23, 5048. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, F.; Tang, M.-J.; Wang, Y.; Dong, J.-H.; Ying, J.-L.; Chen, Y.-L.; Hu, B.; Li, C.; Liu, L.-W. Melatonin confers cadmium tolerance by modulating critical heavy metal chelators and transporters in radish plant. J. Pineal Res. 2020, 69, e12659. [Google Scholar] [CrossRef]
- Xu, L.; Jiang, Q.-W.; Wu, J.; Wang, Y.; Gong, Y.-Q.; Wang, X.-L.; Cecilia, L.; Liu, L.-W. Identification and molecular mapping of the RsDmR Locus conferring resistance to downy mildew at seedling stage in radish. J. Integr. Agric. 2014, 13, 2362–2369. [Google Scholar] [CrossRef]
- Wan, H.-J.; Yuan, W.; Bo, K.-L.; Shen, J.; Pang, X.; Chen, J.-F. Genome-wide analysis of NBS-encoding disease resistance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in Cucurbitaceae crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef]
- Kitashiba, H.; Li, F.; Hirakawa, H.; Kawanabe, T.; Zou, Z.-W.; Hasegawa, Y.; Tonosaki, K.; Shirasawa, S.; Fukushima, A.; Yokoi, S.; et al. Draft sequences of the radish (Raphanus sativus L.) genome. DNA Res. 2014, 21, 481–490. [Google Scholar] [CrossRef]
- Mitsui, Y.; Shimomura, M.; Komatsu, K.; Namiki, N.; Shibata-Hatta, M.; Imai, M.; Katayose, Y.; Mukai, Y.; Kanamori, H.; Kurita, K.; et al. The radish genome and comprehensive gene expression profile of tuberous root formation and development. Sci Rep. 2015, 5, 10835. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-M.; Kim, N.; Ahn, B.-O.; Oh, M.; Chung, W.-H.; Chung, H.; Jeong, S.; Lim, K.-B.; Hwang, Y.-J.; Kim, G.-B.; et al. Elucidating the triplicated ancestral genome structure of radish based on chromosome-level comparison with the Brassica genomes. Theor. Appl. Genet. 2016, 129, 1357–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, T.; Wang, J.; Wang, P.; Qiu, Y.; Zhao, W.; Pang, S.; Li, X.; Wang, H.; Song, J.; et al. Pan-genome of Raphanus highlights genetic variation and introgression among domesticated, wild, and weedy radishes. Mol. Plant 2021, 14, 2032–2055. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.; Jang, H.; Baek, S.; Kim, M.-J.; Yim, B.; ·Huh, S.; Kwon, S.-H.; Yu, H.-J.; Mun, J.-H. An improved Raphanus sativus cv. WK10039 genome localizes centromeres, uncovers variation of DNA methylation and resolves arrangement of the ancestral Brassica genome blocks in radish chromosomes. Theor. Appl. Genet. 2022, 135, 1731–1750. [Google Scholar] [CrossRef]
- Wang, Y.; Song, Z.-H.; Zhang, W.; Xu, L.; Su, X.-J.; Muleke, E.-M.; Liu, L.-W. Identification and characterization of expressed TIR- and non-TIR-NBS-LRR resistance gene analogous sequences from radish (Raphanus sativus L.) de novo transcriptome. Sci. Hortic. 2017, 216, 284–292. [Google Scholar] [CrossRef]
- Finn, R.-D.; Clements, J.; Eddy, S.-R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.-P.; Guo, A.-Y.; Zhang, H.; Luo, J.-C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Finn, R.-D.; Coggill, P.; Eberhardt, R.-Y.; Eddy, S.-R.; Mistry, J.; Mitchell, A.-L.; Potter, S.-C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2015, 44, D279–D285. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef]
- Delorenzi, M.; Speed, T. An HMM model for coiled-coil domains and a comparison with PSSM-based predictions. Bioinformatics 2002, 18, 617–625. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.-D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Chen, H.; Zhang, Y.; Thomas, H.-R.; Frank, M.-H.; He, Y.-H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Hoang, N.-V.; Choe, G.; Zheng, Y.; Aliaga Fandino, A.-C.; Sung, I.; Hur, J.; Kamran, M.; Park, C.; Kim, H.; Ahn, H.; et al. Identification of conserved gene-regulatory networks that integrate environmental sensing and growth in the root cambium. Curr. Biol. 2020, 30, 2887–2900. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.-R.; Baker, B.-J. The evolution of resistance genes in multi-protein plant resistance systems. Curr. Opin. Genet. Dev. 2007, 17, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Castel, B.; Ngou, P.-M.; Cevik, V.; Redkar, A.; Kim, D.-S.; Yang, Y.; Ding, P.; Jones, J.-D.-G. Diverse NLR immune receptors activate defence via the RPW8-NLR NRG1. New Phytol. 2019, 222, 966–980. [Google Scholar] [CrossRef]
- Monteiro, F.; Nishimura, M.T. Structural, functional, and genomic diversity of plant NLR proteins: An evolved resource for rational engineering of plant immunity. Annu. Rev. Phytopathol. 2018, 56, 243–267. [Google Scholar] [CrossRef]
- Collier, S.-M.; Hamel, L.-P.; Moffett, P. Cell death mediated by the N-terminal domains of a unique and highly conserved class of NB-LRR protein. Mol. Plant-Microbe Interact. 2011, 24, 918–931. [Google Scholar] [CrossRef]
- Ma, Y.; Chhapekar, S.-S.; Lu, L.; Oh, S.; Singh, S.; Kim, C.-S.; Kim, S.; Choi, G.-J.; Lim, Y.-P.; Choi, S.-R. Genome-wide identification and characterization of NBS-encoding genes in Raphanus sativus L. and their roles related to Fusarium oxysporum resistance. BMC Plant Biol. 2021, 21, 47. [Google Scholar] [CrossRef]
- Ma, Y.; Chhapekar, S.-S.; Lu, L.; Yu, X.; Kim, S.; Lee, S.-M.; Gan, T.-H.; Choi, G.-J.; Lim, Y.-P.; Choi, S.-R. QTL mapping for Fusarium wilt resistance based on the whole-genome resequencing and their association with functional genes in Raphanus sativus. Theor. Appl. Genet. 2021, 134, 3925–3940. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).