Abstract
NF-Y transcription factor consists of three subsets, A, B, and C, which play various roles during biological processes in plants, particularly in growth and development, stress response, and microorganism interactions. The current study was exhaustively conducted on the NF-Y gene family in Prunus mume (Japanese apricot). We identified 27 PmNF-Y genes that were distributed on eight chromosomes of the Japanese apricot genome, and which were divided into three subgroups according to their phylogenetic relationship. Protein replication analysis showed that the Ka/Ks ratio of one pair of tandem repeats and two pairs of segmental repeats was less than 1. Cis-acting elements upstream of PmNF-Y genes were divided into four distinct categories: light response, growth metabolism, stress, and hormones, among which the numbers of hormone and light-responsive cis-acting elements were higher. RNA-seq analysis revealed that most of the genes were expressed with slight variation in various tissues. Interestingly, some genes were expressed differently during dormancy. During dormancy with exogenous hormones and low temperature, the qRT-PCR results showed that these differentially expressed genes had specific expression responses under GA4, ABA, MeJA, and low-temperature treatments. Therefore, these findings could provide a novel theoretical foundation for future research into the function and molecular regulation mechanism of the NF-Y gene family in the Prunus species.
1. Introduction
Nuclear factor Y (NF-Y), also known as CCAAT-box binding factor (CBF) or heme activator protein (HAP), is a heterotrimeric transcription factor complex in plants that has three distinct subsets: NF-YA (CBF-B, HAP2), NF-YB (CBF-A, HAP3), and NF-YC (CBF-C, HAP5). This transcription factor is highly conserved in evolution, and almost universal in eukaryotes [1]. The NF-YA protein contains a core conserved region consisting of 53 conservative amino acids. The core region consists of two conserved α-helices and a conserved gap sequence connecting the two α-helices. As with the NF-YB and NF-YC proteins, the primary role of their conserved domain is to bind to the CCAAT box and interact with the other two subsets [2,3,4].
Previous studies on NF-Y have proven that it can control flowering time by modulating the blossoming gene, FLOWERING LOCUS T (FT), and the NF-YA, YB, and YC subgroups, all of which play a crucial role in the regulation of the flowering period in plant species [5,6]. The overexpression of Arabidopsis NF-YA subsets (YA1, YA2, YA3, YA4, YA7, and YA10) can inhibit plant flowering time, suggesting that NF-YA subsets negatively regulate plant flowering time [7]. Both NF-YA/B/C and NF-YB-C-CO (CONSTANS) trimers can target the FT-bound promoter region [8]. The irregular time-dependent differential expression rate of CO and NF-Y transcription factors and the protein availability are believed to regulate FT by the NF-YA/B/C and NF-YB/C-CO trimers that are involved in flowering processes. CO protein accumulates continually during the day and peaks at dusk/night [5,8]. Therefore, it can cooperate with NF-YA1/4 to produce the trimeric NF-YB/C-CO complex and induce FT transcription. The protein level of CO decreases gradually during the night, and the related NF-YB/C is substituted by nf-ya1/4 to form the NF-YA/B/C complex and inhibit the expression of FT [9]. Likewise, the overexpression of AtNF-YB1 inhibits the transcription of FT, delaying flowering [10]. NF-Y factor and the DELLA protein, a key component of the gibberellin pathway, also interact to control flowering in plants [11,12]. Arabidopsis AtNF-YA2/B2/C9 has been reported to interact with DELLA and form polymer complexes. GA subsequently causes DELLA protein degradation and NF-Y complex release, allowing it to recruit REF6 and target the NFYBE (NF-Y binding element) cis-acting element binding to the SOC1 promoter region, causing H3K27me3 locus demethylation and SOC1 expression, as well as plant flowering [6].
Most fruit trees are woody perennial plants with flowers, which are important reproductive organs that affect fruit formation or production. However, the fruit production cycle is relatively stable/consistent, which could be affected further by the regulation of the flowering period. Hence, determining the flowering and production time is of economic benefit. In the previous decade, the efficient cultivation of fruit has generated substantial economic benefits and promoted industrialization, with flowering management playing an important role. As a result, further study into the distinct process of flowering regulation in woody fruit trees associated with bud dormancy is required. Prunus mume Sieb. et Zucc. is a deciduous woody fruit tree native to China which is economically significant due to its early flowering time and dual-purpose flowers and fruits. Flowering times vary greatly among its varieties, which have a high genetic diversity. In terms of flowering time, late and early flowering cultivars can be about 30 days. Temperature appears to have a significant influence on flowering time and period. As a result, the flowering period of the same variety in the same region might vary for about 40 days depending on the weather of the year [13], suggesting that Prunus mume blooming time is affected by genetic variation and environmental factors such as temperature. In view of all these, both genetic diversity and temperature are regarded as valuable resources for studying the flowering regulation mechanism of woody plant species. Therefore, a comprehensive genome-wide analysis and different hormone and temperature treatments were undertaken for an in-depth understanding of the NF-Y gene family’s function and its effect in Prunus mume. The results of this current work will enhance our knowledge of the mechanism modulating flowering time in Prunus mume and lay the basis for further research on other Prunus species.
2. Materials and Methods
2.1. Plant Materials and Treatments
The plant materials were collected from the National Field Genebank for Prunus mume of Nanjing Agricultural University. The materials were from a 5-year-old early flowering cultivar, “Taoxingmei”, with strong growth. The sampling period was after the leaves of “Taoxingmei” had fallen. Annual branches of 30–40 cm with full and substantial buds were collected. The branches were divided into four groups and replicated three times with five annual branches. The base of the branch was cut evenly and submerged at a height of 3 cm into 200 μmol/L of GA4 solution, 200 μmol/L of ABA solution, 200 μmol/L of MeJA solution, and distilled water, respectively. After that, they were immediately put in a light incubator for cultivation. The conditions of GA4, ABA, and MeJA treatment were: day and night temperature 21 ± 3 °C, light/dark hours 12/12 h, light intensity 55 mmol m−2 s−1, and relative air humidity 70%, respectively. The conditions of the 4 ℃ treatment were: day and night temperature 4 ± 3 °C, light/dark hours 12/12 h, light intensity 55 mmol m−2 s−1, and relative air humidity 70%, respectively. After every 3 days, 2 mm of the base of the branches was cut to expose the fresh injury to the media. After 0, 1, 3, and 5 days of treatment, the buds were collected from each treatment and stored at −80 °C for future use.
2.2. Identification and Characterization of NF-Y Family in Prunus Mume
First, we downloaded the protein sequence data of Prunus mume from the NCBI database and obtained the HMM model (http://pfam.xfam.org/, accessed on 6 January 2021) database of NF-Y(PF00808) from PFAM. Subsequently, we used the HMM model of NF-Y as the query sequence to obtain relevant sequence data from Prunus mume protein sequence data (E value < 10−5). Furthermore, the acquired sequence confirmed that it contained the fully conserved NF-Y domain on InterPro (http://www.ebi.ac.uk/interpro/, accessed on 6 January 2021) and SMART [14], and the chromosome location information of NF-Y was obtained simultaneously. The ExPaSy (http://web.expasy.org/protparam, accessed on 6 January 2021) online platform was utilized to analyze the physical and chemical properties of NF-Y family member proteins, whereas the BUSCA website (http://busca.biocomp.unibo.it/, accessed on 6 January 2021) was used to predict the subcellular localization of PmNF-Y family members.
2.3. Chromosomal Location and Synteny Analysis
For the chromosome localization analysis of PmNF-Y genes, we used the TBtools program [14]. Meanwhile, the tandem and segmented replication events of PmNF-Y family members were analyzed using MCScanX software (http://chibba.pgml.uga.edu/mcscan2/, accessed on 6 January 2021). Furthermore, we used the KaKs Calculator 2.0 [15] to estimate the Ka and Ks values of tandem and segmented replication gene pairs in order to determine their selection in the evolutionary process.
2.4. Analysis of Structure and Conserved Motifs of NF-Y Gene in Prunus Mume
We used the GSDS tool [16] to analyze and draw the map of the gene structure of PmNF-Y gene family members. Using the MEME tool (http://meme-suite.org/tools/meme, accessed on 8 January 2021), the structure of the conserved domains of PmNF-Y proteins was analyzed. We set the maximum number of pattern discoveries to 10, the other parameters were the default values, and we viewed the results via TBtools software [14].
2.5. Analysis of Cis-Acting Elements in PmNF-Y Gene Upstream Promoter Region
The PlantCARE tool was used to identify the cis-acting components of the 2000 bp domain upstream of PmNF-transcription Y’s initiation point in the Prunus mume genome [17]. These cis-acting elements were subsequently analyzed and classified.
2.6. Phylogenetic Analysis and Prediction of the Protein–Protein Interaction Network of the PmNF-Y Genes Family
A phylogenetic tree was built from the sequences of members of the PmNF-Y gene family of Prunus persica, Prunus dulcis, Prunus armeniaca, Prunus mume, and Arabidopsis thaliana. Protein sequences were aligned utilizing the MAFFT software with the default settings [18]. We constructed phylogenetic trees using the maximum likelihood (ML) method. The ML analysis was conducted using IQ-TREE [19], with ModelFinder software selecting the optimal model [20], and 1000 bootstrap replicates were performed. Members of the PmNF-Y gene family’s protein–protein interactions were explored using the online database STRING (https://string-db.org/, accessed on 10 January 2021).
2.7. RNA-Seq and Quantitative qRT-PCR Analysis of PmNF-Y Gene
RNA-Seq analysis obtained clean reads by removing low-quality reads, reads containing adapters, and more than 10% anonymous nucleotides (N) from raw sequence data (raw reads). Subsequent analyses were conducted based on clean data with high quality and the clean reads were compared to the Prunus mume gene sequence reference data sets using Hisat2. The expression profiles of NF-Y genes in five different tissues—root, stem, leaf, flower bud, and fruit (presented in Table S1)—and the expression in the four different dormancy stages—paradormancy, endodormancy, ecodormancy, and dormancy release (presented in Table S2)—were mapped using the already-existing transcriptome data from the NCBI Sequencing GEO database (accession number PRJNA172987 and PRJNA615074) [21,22]. Among them, the transcriptome data of different dormancy stages had three replicates in each stage, while the transcriptome data of different tissues had only one replicate in each tissue. The RSEM tool was used to calculate gene and transcript expression levels using the FPKM index (fragments per kilobase of transcript per million mapped reads) [23].
The total RNA of flower buds under GA4, ABA, MeJA, and low-temperature treatments was extracted using an RNA extraction kit (FOREGENE, China), the concentrations were determined, and the quality was assured using agarose gel electrophoresis. We utilized the PrimeScript™ RT reagent Kit instructions in conjunction with the gDNA Eraser to synthesize cDNA. The ABI 7300 real-time PCR system (Applied Biosystems, Foster, CA, USA) and the SYBR green real-time PCR Master Mix (Dongbao, Osaka, Japan) were used for the qRT-PCR; the primers are listed in Table S3. The collected flower bud materials were made up of three independent biological replicates. Three technical replicates were performed for each biological replicate [24].
3. Results
3.1. Identification and Characterization of PmNF-Y Gene Family in Japanese Apricot
A total of 27 PmNF-Y genes were identified in the genome of Prunus mume, and family members of the PmNF-Y genes were named PmNF-YA01 to PmNF-YA05, PmNF-YB01 to PmNF-YB13, and PmNF-YC01 to PmNF-YC09 based on their sequence positions on eight chromosomes. The specific physicochemical properties of PmNF-Y genes are shown in Table 1. The proteins encoded by these 27 PmNF-Y genes range from 121 to 351 amino acids in length, and the maximum and minimum molecular weights are, respectively, 13301.29, and 38,208.65 Da. PmNF-YC03 has the smallest isoelectric point, with 4.35, while PmNF-YC01 has the largest, with 9.64. The different isoelectric points of these PmNF-Y proteins indicate that their acid–base characteristics are distinct. All protein GRAVY values are less than 0, suggesting that they are all hydrophilic proteins. Most PmNF-Y genes were located in the nucleus, whereas a few genes were sited in chloroplasts and the cytoplasm.

Table 1.
Identification and physicochemical properties of the NF-Y gene family in the Prunus mume genome.
3.2. Chromosomal Mapping and Synteny Analysis of PmNF-Y Genes
Figure 1 illustrates the chromosomal mapping of the 27 PmNF-Y genes. Among them, seven (PmNF-YC01, PmNF-YB01, PmNF-YB02, PmNF-YB03, PmNF-YB04, PmNF-YB05, and PmNF-YC02) were on Chr01, three (PmNF-YC03, PmNF-YC04, and PmNF-YB06) were on Chr02, seven (PmNF-YB07, PmNF-YA01, PmNF-YC05, PmNF-YB08, PmNF-YB09, PmNF-YA02, and PmNF-YB10) were on Chr03, three (PmNF-YA03, PmNF-YC06, and PmNF-YB11) were on Chr04, two (PmNF-YC07 and PmNF-YB12) were on Chr05, one (PmNF-YB13) was on Chr06, two (PmNF-YC08 and PmNF-YC09) were on Chr07, and two (PmNF-YA04 and PmNF-YA05) were on Chr08. However, Chr01 and Chr03 contain the greatest number of PmNF-Y genes with 25.92%, while Chr06 contains the lowest with 3.70%.
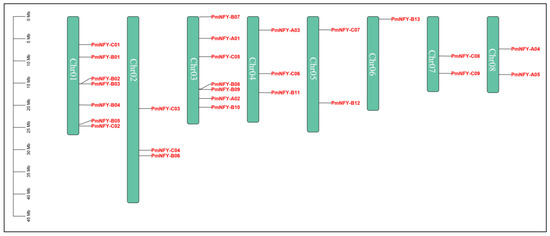
Figure 1.
Distribution of PmNF-Y genes among 8 chromosomes. The scale on the left represents the length of the chromosome. The chromosome numbers are on each chromosome.
3.3. Gene Structural, Protein Conserved Domain, and Conserved Motif Analysis of PmNF-Y Genes
The phylogenetic tree was generated according to protein sequences of PmNF-Y genes to analyze their genetic evolution relationship. Meanwhile, we analyzed the conserved domains of the protein sequences of PmNF-Y genes (Figure 2A) and drew gene structure maps (Figure 2B). It was found that the PmNF-YB04 genes had the highest number of introns (six introns). However, twelve genes did not contain an intron (PmNF-YB02, PmNF-YB05, PmNF-YB06, PmNF-YB09, PmNF-YB10, PmNF-YB011, PmNF-YC02, PmNF-YC03, PmNF-YC04, PmNF-YC05, PmNF-YC08, and PmNF-YC09), three had one intron (PmNF-YB03, PmNF-YB07, PmNF-YB13), two had two introns (PmNF-YB01 and PmNF-YB08), seven had four introns (PmNF-YA01, PmNF-YA02, PmNF-YA03, PmNF-YA04, PmNF-YA05, PmNF-YC06, and PmNF-YC07), and one had five introns (PmNF-YB12).
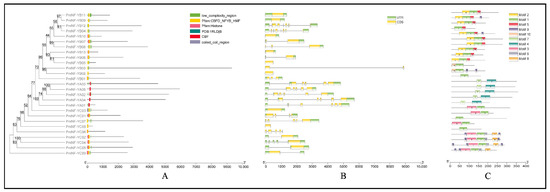
Figure 2.
The protein conserved domains, gene structures, and conserved motifs of PmNF-Y genes based on phylogenetic relationships. (A) Distribution of protein conserved domains. (B) The PmNF-Y gene structures. (C) The PmNF-Y gene conserved motifs.
The analysis of conserved domains revealed that all PmNF-YA genes have the CBF domain, while PmNF-YB and PmNF-YC genes contain the CBFD NFYB HMF domains. Moreover, the analysis results of the conserved motifs of the PmNF-Y gene family via MEME revealed that all PmNF-YA proteins had motifs 4 and 7 (Figure 2C). In addition, all PmNF-YB proteins possessed motif 1, motif 2, and motif 5, except for PmNF-YB01, PmNF-YB02, PmNF-YB03, and PmNF-YB04 (Figure 2C). The majority of PmNF-YC proteins possessed motif 1 and motif 3, except for PmNF-YC06 and PmNF-YC07. Furthermore, PmNF-YC05 contained the greatest number of motifs (7), whereas PmNF-YB02, PmNF-YC06, and PmNF-YC07 contained the lowest number of motifs (1). Therefore, the types and numbers of different gene motifs varied significantly.
The synteny analysis of the PmNF-Y genes revealed that one pair of tandemly repeated sequences (PmNF-YB02 and PmNF-YB03) occurred among the 27 PmNF-Y genes (Figure 3). Interestingly, we identified two pairs of segmental repeat genes (PmNF-YB10 and PmNF-YB11, PmNF-YB06 and PmNF-YB13).
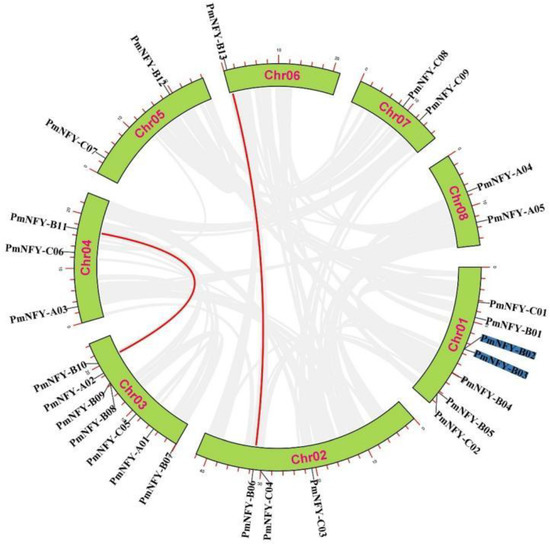
Figure 3.
Synteny relationships of Prunus mume NF-Y genes. PmNF-Y gene duplication relationships. Genes that are tandemly repeated are labeled in blue, and genes that are segmentally duplicated are linked with red lines.
We assessed the Ka/Ks ratio of these tandem repeat and segmented repeat genes to see if they had been subjected to natural selection during evolution. PmNF-Y gene sequences had Ka/Ks values less than 1 (Table 2), signifying that they had undergone purifying selection during their evolutionary process.
Table 2.
The Ka/Ks values of repeated PmNF-Y gene sequences.
3.4. Cis-Acting Elements Analysis in PmNF-Y Gene Promoter Region
The promoters of 2000 bp regions in the upstream region of PmNF-Y were studied to better understand the transcriptional process/mechanism of the PmNF-Y gene. However, the PmNF-Y promoter region contains a large number of cis-acting elements that may be divided into four categories: light-response-related elements type, hormone-response-related elements type, stress-response-related type, and plant growth and development elements type (Figure 4B). The number of light-response-related elements is the greatest, with 344, followed by hormone-related response elements, with a total of 246. Among these, there are 19 salicylic acid response elements, 96 abscisic acid response elements, 20 gibberellin response elements, 92 methyl jasmonate response elements, and 19 auxin response elements. The stress-related response elements came in third, with anaerobic induction, drought-inducibility, low-temperature responsiveness, defense and stress responsiveness, and anoxic specificity among them. Plant growth and development-related components were the weakest, including meristem expression, differentiation of the palisade mesophyll cells, zein metabolism regulation, endosperm expression, seed-specific regulation, circadian control, cell-cycle regulation, and flavonoid biosynthetic regulation (Figure 4A). Among all the members of the PmNF-Y gene family, the PmNF-YC06 genes contained 63 cis-acting elements, and almost all of the gene promoter regions contained more than 15 cis-acting elements (Figure 4B).
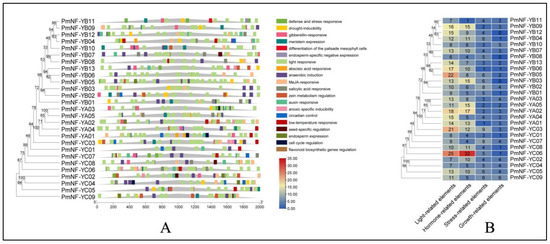
Figure 4.
Predicted cis-acting elements in the promoters of PmNF-Y genes. (A) The 2000 bp sequences upstream of the 27 PmNF-Y genes were analyzed with PlantCARE. (B) Types and numbers of cis-acting elements in the promoter regions of PmNF-Y genes.
Moreover, among all the cis-acting elements, the number of different types of cis-acting elements varied greatly (Figure 5). The most abundant cis-acting elements are hormones, light, and the stress response; however, there are few types for plant growth and development. Hence, we concluded that the PmNF-Y plays a crucial role in the hormone, light, and stress responses in Prunus mume. The majority of the promoters comprised two cis-acting elements, GA and ABA, according to further examination. ABA, for instance, is currently assumed to have a major role in seed and bud dormancy. It also contains a chemical that prevents the release of bud dormancy and is believed to be responsible for initiating the “switch” of bud dormancy [25]. Gibberellin, as a dormancy breaker, can substitute some components of the cold-required conditions and promote/induce the early dormancy-breaking of fruit tree flower buds, with unexpected results [26]. This implies that members of the PmNF-Y family may be involved in plant dormancy modulation.
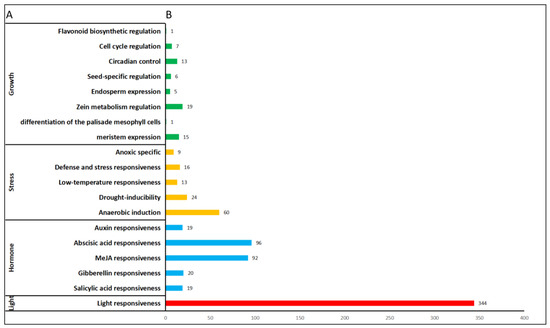
Figure 5.
Statistics and classification of the cis-acting elements of PmNF-Y. (A) The cis-acting elements in the 2000 bp region upstream of PmNF-Y were divided into four groups according to their function. (B) The numbers of each cis-acting element of PmNF-Y.
3.5. Phylogenetic Tree of the PmNF-Y Genes
The phylogenetic tree of 158 NF-Y genes, including 27, 36, 34, 29, and 32 from Prunus mume, Arabidopsis thaliana, Prunus salicina, Prunus armeniaca, and Prunus persica, respectively, was constructed based on their amino acid sequences using IQ-TREE. On the basis/foundation of their sequences’ similarities, these proteins, including 27 PmNF-Y in Prunus mume, were grouped into three groups (Figure 6). The integrated phylogenetic tree’s classification results were essentially consistent with that generated via PmNF-Y protein sequences (Figure 1). Meanwhile, the conserved motif and protein conserved domain results indicate that the same subfamily has similar numbers and types of conserved motifs and protein conserved domains, which further supports the phylogenetic tree clustering results (Figure 1). The number of conserved motifs and protein conserved domains within the same subfamily suggests that they are relatively conserved and undertake/conduct similar functions.
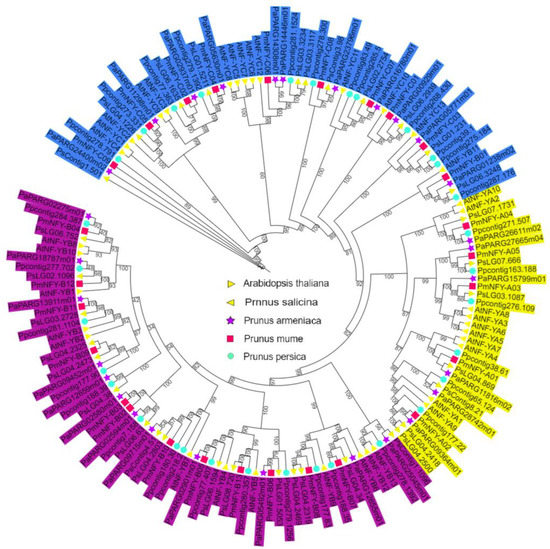
Figure 6.
Phylogenetic analysis of NF-Y proteins from Prunus mume and four other plant species.
3.6. Prediction of Protein–Protein Interaction Network
PmNF-Y gene regulatory relationships were predicted/hypothesized and elucidated. Almost every gene in the PmNF-Y gene family is a member of the regulatory network. In addition, three genes, PmNF-YA03, PmNF-YB07, and PmNF-YC09, interacted with regulatory network genes including PmABI5, PmKEG, PmDR1-AC, PmATRX-like, PmB3-LEC2, and PmRGL2 (Figure 7).
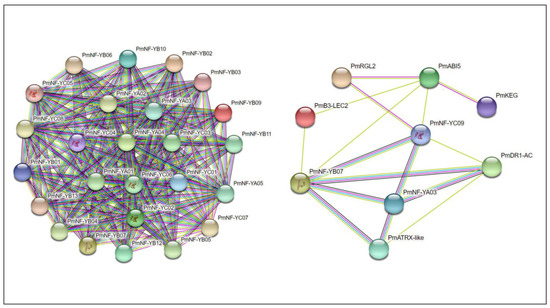
Figure 7.
Prediction and analysis of protein–protein interactions of PmNF-Y genes.
3.7. Expression Profile Analysis of PmNF-Y Genes in Different Tissues and Dormancy Periods
Using the already-existing RNA-seq data, we investigated the expression levels of the identified 27 PmNF-Y genes in leaves, roots, buds, stems, and fruits to determine their specific expression patterns in different tissues. The findings revealed that the expression levels of these genes differed significantly in Prunus mume tissues. However, the PmNF-YC02 gene had the highest expression level in roots, whereas PmNF-YB03, PmNF-YB07, PmNF-YB10, and PmNF-YC06 genes were not expressed in roots. The PmNF-YA01 gene had the highest expression levels in the stem, while PmNF-YB05, PmNF-YB07, PmNF-YB08, PmNF-YB10, PmNF-YC06, and PmNF-YC08 were not expressed in the stem. PmNF-YB04 was found to be the highest-expressed in leaves, buds, and fruit, while PmNF-YC06 was not expressed in all tissues. Furthermore, PmNF-YB10 was exclusively expressed in the bud and fruit, whereas the PmNF-YB08 gene was only expressed in the root.
Moreover, we used the already-existing RNA-seq data to determine the expression of PmNF-Y genes at distinct dormancy stages. The expression level of the PmNF-YB04 gene was the highest at all four stages, as shown in Figure 8B, with no significant difference between them. At the paradormancy, ecodormancy, and endodormancy stages, the PmNF-YC06 gene was not expressed. Four genes (PmNF-YB06, PmNF-YB08, PmNF-YC06, PmNF-YC08) were not expressed during dormancy breaking. The expression of PmNF-YB07 and PmNF-YA05 increased from the first to the second, third, and fourth stages. PmNF-YB08, PmNF-YC08, and PmNF-YC09 expression levels initially increased at the first, second, and third stages, then decreased at the fourth stage. PmNF-YA03, PmNF-YB10, and PmNF-YB13 expression levels, on the other hand, initially dropped/fell and subsequently rose. As a result of the significant differences in their expression levels at different stages of dormancy, these genes may play a key role in the control/modulation of the dormancy process.

Figure 8.
Expression levels of PmNF-Y genes in different tissues and at different stages of dormancy: (A) expression profiles of NF-Y genes in five different tissues; (B) expression profile of NF-Y genes in the four different dormancy stages.
3.8. Expression Pattern Analysis of PmNF-Y Genes under GA4, ABA, MeJA, and Low-Temperature Treatments
To further study the role of PmNF-Y genes in the dormancy process, we screened the transcriptome for differentially expressed PmNF-Y genes at distinct stages of dormancy in the bud tissue and performed qRT-PCR to examine their expression levels under the 200 mol/L GA4, 200 mol/L ABA, 200 mol/L MeJA, and 4 °C low-temperature treatments. However, under the GA4 hormone treatment, the expression level of PmNF-YA03, PmNF-YB07, and PmNF-YC09 genes initially increased, then decreased, and then stabilized, whereas the expression levels of the PmNF-YB08 and PmNF-YB10 genes increased. On the fifth day, the expression levels of PmNF-YB13 and PmNF-YC08 increased significantly, whereas the expression level of the PmNF-YA05 gene decreased. Under the ABA hormone treatment, the expression levels of the PmNF-YA03 and PmNF-YA05 genes also initially increased and then decreased, while the PmNF-YB13, PmNF-YC08, and PmNF-YC09 genes initially increased, then decreased, and finally increased. Additionally, the expression level of the PmNF-YB07 gene decreased, whereas the PmNF-YB08 and PmNF-YB10 genes increased. Under the MeJA hormone treatment, the expression levels of PmNF-YA03, PmNF-YA05, PmNF-YB07, and PmNF-YB10 decreased and reached approximately zero. On the fifth day, the expression levels of PmNF-YB13 increased significantly. Except for the PmNF-YA03 gene, the levels of all other genes increased in response to the 4 °C low-temperature treatment. Eight members of the PmNF-Y gene family from flower buds (in the endodormancy stage) showed distinct expression responses to various exogenous hormones and low-temperature treatments. The results are presented in Figure 9.
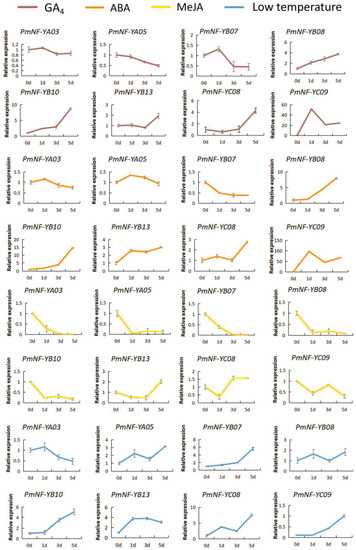
Figure 9.
Expression analysis of the PmNF-Y genes in Prunus mume under GA4, ABA, MeJA, and low-temperature treatments.
4. Discussion
NF-Y is encoded by a single gene in yeast and human genomes; however, in plants, NF-Y Subsets are encoded by a gene family of many genes, resulting in a large number of potential NF-Y genes. These distinct complexes may exhibit functional differentiation and redundancy due to their trimer composition. With the NF-YA and NF-YC subgroups, one subset of the NF-Y family forms a heterotrimer. To activate or regulate downstream gene expression, the heterotrimer complex interacts with the CCAAT box [27,28,29]. NF-YS had been reported to be associated with the regulation of plant growth and development processes as well as biological and abiotic stress responses, including embryonic development, seed maturation, post-embryonic development, flowering time, root growth and development, fatty acid synthesis, endoplasmic reticulum stress response, and others [30,31,32,33,34]. However, it has been reported in recent years that CO can substitute NF-YA and interact with NF-YB/NF-YC [35]. It forms a heterotrimer, specifically binds particularly to CCACA components, and is involved in flowering modulation [36,37]. Recent studies [38] have indicated that the PtNF-YB1 gene can regulate flowering time. Moreover, the NF-Y complex composed of NF-YA2, NF-YB2/NF-YB3, and NF-YC3/NF-YC9 directly regulates the flowering regulation genes SOC1 and FT, and the complex can interact with the CO gene of the photoperiodic pathway and the DELLAS gene of the gibberellin inhibitor in the flowering regulation pathway to cooperatively regulate the flowering time of Arabidopsis thaliana [39]. As an inhibitor of GA signals, the DELLA protein, has been proven to be related to NF-Y in several studies/reports. At the same time studies in tobacco found that GA3 treatment elevated AsNF-YB3 levels, which were similarly influenced by ABA and MeJA [40]. The NF-Y complex has recently been discovered to be implicated in the GA-induced dormancy transition in Arabidopsis. RGA inhibits flowering by preventing the interaction between NF-Y with CO [36]. NF-YC and RGL2 synergistically restricted the GA-mediated germination of seeds, according to Liu et al. (2016). They also noted that distinct DELLA-NF-Y (C) combinations have played a role in various biological processes. Consequently, these genes and their interaction are necessary for the regulation of flowering time and dormancy, and they also play significant roles in several diverse biological processes in plant species.
In this current study, HMMER 3.0 was used to identify PmNF-Y genes, and a detailed list of 27 PmNF-Y genes was established in Prunus mume (Table 1). Their physicochemical features were analyzed to predict their subcellular location. Different PmNF-Y proteins vary substantially in molecular weight, and their structure and composition may vary, indicating that their functions could be different. The majority of PmNF-Y genes are located in the nucleus, with a few in the chloroplasts and cytoplasm. Transcription factors are essential controllers of gene expression in the nucleus. The replication of genes serves a crucial function in biological evolution [41]. However, its related analysis revealed one gene pair (PmNF-YB02/PmNF-YB03) as a tandem repeat and two gene pairs (PmNF-YB10/PmNF-YB11, PmNF-YB06/PmNF-YB13) as segmental repeats (Figure 3), indicating that tandem and segmental repeats additionally support the PmNF-Y gene family’s evolution and diversity. Moreover, the segmental gene pairs’ Ka/Ks values were calculated. Here, we noticed that the Ka/Ks ratio of all PmNF-Y gene pairs is less than 1, implying that they are subjected to significant purification selection during evolution and might evolve slowly, which could avoid/prevent the transmission of genetic variations or mutations. The 27 PmNF-Y genes were subdivided into three subfamilies based on the phylogenetic tree. The greater level of homology between the PmNF-Y protein sequences of various Rosaceae plants within the same subfamily indicates that the gene structure of the NF-Y protein in Prunus plants is highly conserved. However, there are significant disparities in the number of motifs, protein conserved domains, and gene structure amongst subfamilies, indicating that they have distinct biological functions in plant species. We predicted the interactions between genes within PmNF-Y and other related key genes and found that PmNF-Y genes were not only highly interconnected within the family, but PmNF-YA03, PmNF-YB07, and PmNF-YC09 also interacted with PmABI5, PmKEG, PmDR1-AC, PmATRX-like, PmB3-LEC2, and PmRGL2. ABI5 is an ABA-signaling controller factor that integrates several signals, controls the gene expression of stress responses, and controls a variety of mechanisms: lateral root development, seed germination, photosynthesis, and seedling growth [42,43,44]. Meanwhile, E3 ligase KEG has been found to regulate plant immunity in Arabidopsis [45]. However, the Aux/IAAs gene family member, DR1-AC genes, have been expressed during fruit development and ripening, which are regulated via ethylene and auxin [7]. PmB3-LEC2 is involved in seed development, and miR395abcd’s control of its target gene ATRX plays a positive protective role in the low-temperature response in cassava [46,47]. RGL2 contributes to GA-induced floral development, including stamen growth/expansion and anther dehiscence, as well as being a negative regulator of GA-mediated seed germination in Arabidopsis [48]. The upstream promoter region of the PmNF-Y gene analysis results revealed that cis-acting elements could be divided into four categories: light response, growth metabolism, stress, and hormones. The PmNF-Y gene plays a significant role in several stress and hormone responses, suggesting that the PmNF-Y gene could be involved in the growth and development of Prunus mume. Except for photo-response, most cis-acting elements are hormones, while GA, ABA, and MeJA are, respectively, the main hormones. GA, ABA, and MeJA have always been considered the most closely related hormones in the process of plant dormancy [26,49]. Plant hormones play an important role in the regulation of plant dormancy. In the previous research of our research group, we found that the content of GA was the highest in the period of flower bud dormancy release, and there was a significant difference in the four dormancy stages. It was speculated that GA was related to dormancy release: ABA content was the highest in the paradormancy stage, decreased in the endodormancy and ecodormancy stages, and was the lowest in the dormancy release stage. The content of JA was relatively low in the period of paradormancy and endodormancy, while the content of JA was significantly higher in the period of ecodormancy than that of endodormancy. The content of JA was the highest at the dormancy release stage, which indicated that JA had an inhibitory effect on the flower bud dormancy of Prunus mume. Moreover, the expression levels of several genes (PmNF-YA03, PmNF-YA05, PmNF-YB07, PmNF-YB08, PmNF-YB10, PmNF-YB13, PmNF-YC08, and PmNF-YC09) varied significantly at different stages of dormancy. Furthermore, specific expression responses to different exogenous hormones and low-temperature treatments were identified in eight PmNF-Y genes. Therefore, we hypothesized that these genes may exhibit diverse mechanisms and functions that regulate plant dormancy.
5. Conclusions
We identified 27 PmNF-Y genes in the Prunus mume genome. Based on their evolutionary relationship, we grouped them into three subgroups, PmNF-YA, PmNF-YB, and PmNF-YC. The genes contain one (1) pair of tandem repeats and two (2) pairs of segmental repeats, with a Ka/Ks of less than one (1). The studied PmNF-Y gene has four types of cis-acting elements upstream: light response, growth metabolism, stress, and hormones, with hormones and light response containing the most cis-acting elements. According to RNA-seq data, most genes’ expression varied slightly in different tissues. PmNF-YA03, PmNF-YA05, PmNF-YB07, PmNF-YB08, PmNF-YB10, PmNF-YB13, PmNF-YC08, and PmNF-YC09 genes varied during dormancy. The qRT-PCR results showed that differentially expressed genes responded specifically to GA4, ABA, MeJA, and low temperature. The genome-wide identification of the NF-Y transcription factor helps to understand the evolution and function of the NF-Y gene family in Prunus mume and provides a basis for further functional studies of these genes in Prunus genus species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8121180/s1, Table S1: Expression profile analysis of PmNF-Y genes in different tissues; Table S2: Expression profile analysis of PmNF-Y genes in different dormancy periods; Table S3: Primer sequences used in qRT-PCR.
Author Contributions
Z.G. and T.S. designed and supervised the study. F.G., X.H., R.W., Y.M., C.M. and Y.B. performed the experiment. Z.N. and P.Z. prepared the materials. F.G., X.H. and W.T. performed data analysis. F.G. drafted the manuscript. D.C. edited the manuscript. F.G., S.S., D.C., T.S. and Z.G. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key Research and Development Program of China (2020YFE0202900), the “JBGS” Project of Seed Industry Revitalization in Jiangsu Province (JBGS (2021) 019), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) in material collection, data analysis, and experiment.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This manuscript and Supplemental Materials include all data analyzed or generated during the present study. The Prunus mume NF-Y gene family sequences used in this study were downloaded from the NCBI Genbank (https://www.ncbi.nlm.nih.gov, accessed on 1 January 2021).
Acknowledgments
Thanks to all the researchers for their contributions to this study.
Conflicts of Interest
All the authors declare no conflict of interest.
Abbreviations
| ABA | Abscisic acid |
| GA | Gibberellic acid |
| MeJA | Methyl jasmonate |
| NF-Y | Nuclear transcription factor Y |
| Pm | Prunus mume |
| NCBI | National Center for Biotechnology Information |
References
- Mantovani, R. The Molecular Biology of the CCAAT-Binding Factor NF-Y. Gene 1999, 239, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Zhang, S.; Olesen, J.T.; Rich, A.; Guarente, L. Subunit Interaction in the CCAAT-Binding Heteromeric Complex Is Mediated by a Very Short Alpha-Helix in HAP2. Proc. Natl. Acad. Sci. USA 1994, 91, 3009–3013. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Fikes, J.D.; Guarente, L. Mutations in Yeast HAP2/HAP3 Define a Hybrid CCAAT Box Binding Domain. EMBO J. 1993, 12, 4647–4655. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, R.; Li, X.-Y.; Pessara, U.; Van Huisjduijnen, R.H.; Benoist, C.; Mathis, D. Dominant Negative Analogs of NF-YA. J. Biol. Chem. 1994, 269, 20340–20346. [Google Scholar] [CrossRef] [PubMed]
- Siriwardana, C.L.; Gnesutta, N.; Kumimoto, R.W.; Jones, D.S.; Myers, Z.A.; Mantovani, R.; Holt III, B.F. Nuclear Factor Y, Subunit A (NF-YA) Proteins Positively Regulate Flowering and Act through Flowering Locus T. PLoS Genet. 2016, 12, e1006496. [Google Scholar] [CrossRef]
- Hwang, K.; Susila, H.; Nasim, Z.; Jung, J.-Y.; Ahn, J.H. Arabidopsis ABF3 and ABF4 Transcription Factors Act with the NF-YC Complex to Regulate SOC1 Expression and Mediate Drought-Accelerated Flowering. Mol. Plant 2019, 12, 489–505. [Google Scholar] [CrossRef]
- Xu, D.Q. Construction of Plant Overexpression Vector of Tomato DR1 Gene and Its Transformation into Tomato. Master’s Thesis, Southwest Agricultural University, Chongqing, China, 2004. [Google Scholar]
- Kumimoto, R.W.; Zhang, Y.; Siefers, N.; Holt III, B.F. NF–YC3, NF–YC4 and NF–YC9 Are Required for CONSTANS-mediated, Photoperiod-dependent Flowering in Arabidopsis Thaliana. Plant J. 2010, 63, 379–391. [Google Scholar] [CrossRef]
- Fornari, M.; Calvenzani, V.; Masiero, S.; Tonelli, C.; Petroni, K. The Arabidopsis NF-YA3 and NF-YA8 Genes Are Functionally Redundant and Are Required in Early Embryogenesis. PLoS One 2013, 8, e82043. [Google Scholar] [CrossRef]
- Li, C.; Distelfeld, A.; Comis, A.; Dubcovsky, J. Wheat Flowering Repressor VRN2 and Promoter CO2 Compete for Interactions with NUCLEAR FACTOR-Y Complexes. Plant J. 2011, 67, 763–773. [Google Scholar] [CrossRef]
- Hou, X.; Zhou, J.; Liu, C.; Liu, L.; Shen, L.; Yu, H. Nuclear Factor Y-Mediated H3K27me3 Demethylation of the SOC1 Locus Orchestrates Flowering Responses of Arabidopsis. Nat. Commun. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Xu, F.; Li, T.; Xu, P.; Li, L.; Du, S.; Lian, H.; Yang, H. DELLA Proteins Physically Interact with CONSTANS to Regulate Flowering under Long Days in Arabidopsis. FEBS Lett. 2016, 590, 541–549. [Google Scholar] [CrossRef]
- Chu, M. Chinese Fruit Trees-Prunus Mume [M]; China Forestry Press: Beijing, China, 1999. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-Wide Analysis of VQ Motif-Containing Proteins in Moso Bamboo (Phyllostachys Edulis). Planta 2017, 246, 165–181. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G. The Genome of Prunus Mume. Nat. Commun. 2012, 3, 1–8. [Google Scholar] [CrossRef]
- Gao, J.; Ni, X.; Li, H.; Hayat, F.; Shi, T.; Gao, Z. MiR169 and PmRGL2 Synergistically Regulate the NF-Y Complex to Activate Dormancy Release in Japanese Apricot (Prunus Mume Sieb. et Zucc.). Plant Mol. Biol. 2021, 105, 83–97. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, L.; Mai, Y.; Han, W.; Suo, Y.; Diao, S.; Sun, P.; Fu, J. Phytohormone and Integrated MRNA and MiRNA Transcriptome Analyses and Differentiation of Male between Hermaphroditic Floral Buds of Andromonoecious Diospyros Kaki Thunb. BMC Genom. 2021, 22, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Zhong, W.; Huo, X.; Zhuang, W.; Ni, Z.; Gao, Z. Expression Analysis of ABA-and GA-Related Genes during Four Stages of Bud Dormancy in Japanese Apricot (Prunus Mume Sieb. et Zucc). J. Hortic. Sci. Biotechnol. 2016, 91, 362–369. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wang, L.; Zhong, W.; Ni, Z.; Zhang, Z. Comparative Proteomic and Transcriptomic Approaches to Address the Active Role of GA4 in Japanese Apricot Flower Bud Dormancy Release. J. Exp. Bot. 2013, 64, 4953–4966. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hu, P.; Huang, M.; Tang, Y.; Li, Y.; Li, L.; Hou, X. The NF-YC–RGL2 Module Integrates GA and ABA Signalling to Regulate Seed Germination in Arabidopsis. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Martyn, G.E.; Quinlan, K.G.; Crossley, M. The Regulation of Human Globin Promoters by CCAAT Box Elements and the Recruitment of NF-Y. Biochim. Et Biophys. Acta (BBA)-Gene Regul. Mech. 2017, 1860, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, A.; Bellorini, M.; Mongelli, N.; Mantovani, R. CCAAT-Box Binding Protein NF-Y (CBF, CP1) Recognizes the Minor Groove and Distorts DNA. Nucleic Acids Res. 1995, 23, 4565–4572. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Howell, S.H. BZIP28 and NF-Y Transcription Factors Are Activated by ER Stress and Assemble into a Transcriptional Complex to Regulate Stress Response Genes in Arabidopsis. Plant Cell 2010, 22, 782–796. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; De Mita, S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT-Box Binding Transcription Factors in Plants: Y so Many? Trends Plant Sci. 2013, 18, 157–166. [Google Scholar] [CrossRef]
- Myers, Z.A.; Holt III, B.F. NUCLEAR FACTOR-Y: Still Complex after All These Years? Curr. Opin. Plant Biol. 2018, 45, 96–102. [Google Scholar] [CrossRef]
- Romier, C.; Cocchiarella, F.; Mantovani, R.; Moras, D. The NF-YB/NF-YC Structure Gives Insight into DNA Binding and Transcription Regulation by CCAAT Factor NF-Y. J. Biol. Chem. 2003, 278, 1336–1345. [Google Scholar] [CrossRef]
- Sato, H.; Mizoi, J.; Tanaka, H.; Maruyama, K.; Qin, F.; Osakabe, Y.; Morimoto, K.; Ohori, T.; Kusakabe, K.; Nagata, M. Arabidopsis DPB3-1, a DREB2A Interactor, Specifically Enhances Heat Stress-Induced Gene Expression by Forming a Heat Stress-Specific Transcriptional Complex with NF-Y Subunits. Plant Cell 2014, 26, 4954–4973. [Google Scholar] [CrossRef]
- Luo, X.; Gao, Z.; Wang, Y.; Chen, Z.; Zhang, W.; Huang, J.; Yu, H.; He, Y. The NUCLEAR FACTOR-CONSTANS Complex Antagonizes Polycomb Repression to De-repress FLOWERING LOCUS T Expression in Response to Inductive Long Days in Arabidopsis. Plant J. 2018, 95, 17–29. [Google Scholar] [CrossRef]
- Mach, J. CONSTANS Companion: CO Binds the NF-YB/NF-YC Dimer and Confers Sequence-Specific DNA Binding. Plant Cell 2017, 29, 1183. [Google Scholar] [CrossRef]
- Brambilla, V.; Fornara, F. Y Flowering? Regulation and Activity of CONSTANS and CCT-Domain Proteins in Arabidopsis and Crop Species. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2017, 1860, 655–660. [Google Scholar] [CrossRef]
- Wang, R.; Zhu, L.; Zhang, Y.; Fan, J.; Li, L. Genome-Wide Analysis of Poplar NF-YB Gene Family and Identified PtNF-YB1 Important in Regulate Flowering Timing in Transgenic Plants. BMC Plant Biol. 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Cao, S.; Kumimoto, R.W.; Gnesutta, N.; Calogero, A.M.; Mantovani, R.; Holt III, B.F. A Distal CCAAT/NUCLEAR FACTOR Y Complex Promotes Chromatin Looping at the FLOWERING LOCUS T Promoter and Regulates the Timing of Flowering in Arabidopsis. Plant Cell 2014, 26, 1009–1017. [Google Scholar] [CrossRef]
- Sun, X.; Ren, Y.; Zhang, X.; Lian, H.; Zhou, S.; Liu, S. Overexpression of a Garlic Nuclear Factor Y (NF-Y) B Gene, AsNF-YB3, Affects Seed Germination and Plant Growth in Transgenic Tobacco. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 513–523. [Google Scholar] [CrossRef]
- Casola, C.; Betrán, E. The Genomic Impact of Gene Retrocopies: What Have We Learned from Comparative Genomics, Population Genomics, and Transcriptomic Analyses? Genome Biol. Evol. 2017, 9, 1351–1373. [Google Scholar] [CrossRef]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Takumi, S. Positive Role of a Wheat HvABI5 Ortholog in Abiotic Stress Response of Seedlings. Physiol. Plant. 2008, 134, 74–86. [Google Scholar] [CrossRef]
- Utsugi, S.; Ashikawa, I.; Nakamura, S.; Shibasaka, M. TaABI5, a Wheat Homolog of Arabidopsis Thaliana ABA Insensitive 5, Controls Seed Germination. J. Plant Res. 2020, 133, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Zinsmeister, J.; Lalanne, D.; Terrasson, E.; Chatelain, E.; Vandecasteele, C.; Vu, B.L.; Dubois-Laurent, C.; Geoffriau, E.; Signor, C.L.; Dalmais, M. ABI5 Is a Regulator of Seed Maturation and Longevity in Legumes. Plant Cell 2016, 28, 2735–2754. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Sun, P.; Wang, W.; Tang, D. Arabidopsis E3 Ligase KEG Associates with and Ubiquitinates MKK4 and MKK5 to Regulate Plant Immunity. J. Integr. Plant Biol. 2021, 63, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Zhou, Y.; Peng, M. The Differential Analysis of MiR395abcd and Its Target Gene ATRX Response to Low Temperature in Different Cold Tolerance Cassava Varieties. Genom. Appl. Biol. 2015, 34, 2441–2447. [Google Scholar]
- Li, Y. Origin and Evolution of Seed Development-Related B3 Genes. Master’s Thesis, Fudan University of China, Shanghai, China, 2011. [Google Scholar]
- Gómez, M.D.; Fuster-Almunia, C.; Ocaña-Cuesta, J.; Alonso, J.M.; Pérez-Amador, M.A. RGL2 Controls Flower Development, Ovule Number and Fertility in Arabidopsis. Plant Sci. 2019, 281, 82–92. [Google Scholar] [CrossRef]
- Ríos, G.; Leida, C.; Conejero, A.; Badenes, M.L. Epigenetic Regulation of Bud Dormancy Events in Perennial Plants. Front. Plant Sci. 2014, 5, 247. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).