
Unlocking Plum Genetic Potential: Where Are We At?

 ,
,  , and
, and
Abstract
:1. Introduction
1.1. Plum Production and Market Aspects
1.2. The Rationale for Plum Breeding
2. Prunus Taxonomy, Species Identification and Diversity
3. Plum Species Characteristics, Origin and Diversity
3.1. European Plum (P. Domestica L.)
3.2. Japanese Plum (Prunus Salicina Lindl.)
4. Plum Varietal Diversity
5. New Plum Cultivars Obtained by Breeding
6. Modern Breeding Objectives and Programs
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chin, S.W.; Shaw, J.; Haberle, R.; Wen, J.; Potter, D. Diversification of almonds, peaches, plums and cherries—Molecular systematics and biogeographic history of Prunus (Rosaceae). Mol. Phylogenet. Evol. 2014, 76, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Huang, C.H.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of rosaceae fruit types based on nuclear phylogeny in the context of geological times and genome duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, M.; Suranyi, D. Origin and Dissemination of Plums. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1999; Volume 23, pp. 179–231. ISBN 9780471254454. [Google Scholar]
- Topp, B.L.; Russell, D.M.; Neumüller, M.; Dalbó, M.A.; Liu, W.P. Fruit Breeding, Handbook of Plant Breeding 8; Badenes, M.L., Byrne, D.H., Eds.; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2012; pp. 571–621. ISBN 9781441907639. [Google Scholar]
- Sottile, F.; Peano, C.; Mezzetti, B.; Capocasa, F.; Bellini, E.; Nencetti, V.; Palara, U.; Pirazzini, P.; Mennone, C.; Catalano, L. Plum Production in Italy: State of Art and Perspectives. Acta Hort. 2010, 874, 25–36. [Google Scholar] [CrossRef]
- Centre for the Promotion of Imports from Developing Countries (CBI). The European Market Potential for Fresh Plums and Other Stone Fruit; CBI: The Hague, The Netherlands, 2020; Available online: https://www.cbi.eu (accessed on 11 November 2021).
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 21 November 2021).
- USDA. Fruit and Tree Nuts Situation and Outlook Yearbook. In Market and Trade, Economics Division, Economic Research Service; U.S. Department of Agriculture: Singapore, 2021. [Google Scholar]
- Okie, W.R.; Hancock, J.F. Plums. In Temperate Fruit Crop Breeding; Hancock, J.F., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 337–358. [Google Scholar]
- Sottile, F.; D’Aquino, S.; Peano, C.; Girgenti, V.; Giuggioli, N.R.; Palma, A. Effect of Palletized Map Storage on the Quality and Nutritional Compounds of the Japanese Plum cv. Angeleno (Prunus salicina Lindl.). J. Food Processing Preserv. 2016, 41, 1–10. [Google Scholar]
- Giuggioli, N.; Sottile, F.; Peano, C. Quality indicators for modified atmosphere packaging (MAP) storage of high-quality European plum (Prunus domestica L.) cultivars. Ital. J. Food Sci. 2016, 28, 376–390. [Google Scholar]
- Taiti, C.; Pandolfi, C.; Caparrotta, S.; Dei, M.; Giordani, E.; Mancuso, S.; Nencetti, V. Fruit aroma and sensorial characteristics of traditional and innovative Japanese plum (Prunus salicina Lindl.) cultivars grown in Italy. Eur. Food Res. Technol. 2019, 245, 2655–2668. [Google Scholar] [CrossRef]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Kader, A. Antioxidant Capacities, Phenolic Compounds, Carotenoids, and Vitamin C Contents of Nectarine, Peach, and Plum Cultivars from California. J. Agric. Food Chem. 2002, 50, 4976–4982. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Kang, X.; He, X.; Dong, M.; Zhang, Q.; Liu, R.H. Cellular antioxidant activity of common vegetables. J. Agric. Food Chem. 2008, 56, 8418–8426. [Google Scholar] [CrossRef]
- Impallari, F.M.; Monte, M.; Girgenti, V.; Del Signore, M.B.; Sottile, F. Biodiversity of Sicilian Fruit Trees: Studies on Plums. Acta Hort. 2010, 874, 37–44. [Google Scholar] [CrossRef]
- Sottile, F.; Peano, C.; Giuggioli, N.R.; Girgenti, V. The effect of modified atmosphere packaging on the physical and chemical quality of fresh yellow plum cultivars. Int. J. Food Agric. Environ. 2013, 11, 363–367. [Google Scholar]
- Wolf, J.; Göttingerová, M.; Kaplan, J.; Kiss, T.; Venuta, R.; Nečas, T. Determination of the pomological and nutritional properties of selected plum cultivars and minor fruit species. Hortic. Sci. 2020, 47, 181–193. [Google Scholar] [CrossRef]
- Igwe, E.O.; Charlton, K.E. A Systematic Review on the Health Effects of Plums (Prunus domestica and Prunus salicina). Phyther. Res. 2016, 30, 701–731. [Google Scholar] [CrossRef] [PubMed]
- Zhebentyayeva, T.; Shankar, V.; Scorza, R.; Callahan, A.; Ravelonandro, M.; Castro, S.; DeJong, T.; Saski, C.A.; Dardick, C. Genetic characterization of worldwide Prunus domestica (plum) germplasm using sequence-based genotyping. Hortic. Res. 2019, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prunus Crop Germplasm Committee (PCGC). Prunus Vulnerability Statement; PCGC: Santa Ana, CA, USA, 2017; pp. 1–13. Available online: https://www.ars-grin.gov/npgs/cgc_reports/prunusvuln2017.pdf (accessed on 23 November 2021).
- Boonprakob, U.; Byrne, D.H.; Graham, C.J.; Okie, W.R.; Beckman, T.; Smith, B.R. Genetic relationships among cultivated diploid plums and their progenitors as determined by RAPD markers. J. Am. Soc. Hortic. Sci. 2001, 126, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.J.; Gross, B.L. From forest to field: Perennial fruit crop domestication. Am. J. Bot. 2011, 98, 1389–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, B.I.; Guerra, M.E.; Herrera, S.; Irisarri, P.; Pina, A.; Rodrigo, J. Genetic diversity and population structure of japanese plum-type (Hybrids of P. salicina) accessions assessed by ssr markers. Agronomy 2021, 11, 1748. [Google Scholar] [CrossRef]
- DeBuse, C.J.; Shaw, D.V.; DeJong, T.M. Response to inbreeding of early seedling growth and fruit traits in a Prunus domestica L. breeding population. Acta Hortic. 2013, 985, 87–96. [Google Scholar] [CrossRef]
- Goldschmidt, E.E. The Evolution of Fruit Tree Productivity: A Review. Econ. Bot. 2013, 67, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Lansari, A.; Kester, D.E.; Iezzoni, A.F. Inbreeding, Coancestry, and Founding Clones of Almonds of California, Mediterranean Shores, and Russia. J. Am. Soc. Hortic. Sci. 1994, 119, 1279–1285. [Google Scholar] [CrossRef] [Green Version]
- Okie, W.R.; Ramming, D.W. Plum breeding worldwide. HortTechnology 1999, 9, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Motisi, A.; Pernice, F.; Sottile, F.; Caruso, T. Rootstock effect on stem water potential gradients in cv. ‘armking’ nectarine trees. Acta Hortic. 2004, 658, 75–79. [Google Scholar] [CrossRef]
- Gianní, S.; Sottile, F. In vitro storage of plum germplasm by slow growth. J. Hortic. Sci. (Prague) 2015, 42, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Sottile, F.; Del Signore, M.B.; Massai, R.; Capocasa, F.; Mezzetti, B. Rootstocks evaluation for European and Japanese plums in Italy. Acta Hort. 2012, 968, 137–146. [Google Scholar] [CrossRef]
- Sottile, F.; Del Signore, M.B.; Girgenti, V.; Giuggioli, N.R.; Peano, C. Quality of autochthonous Sicilian plums. Ital. J. Food Sci. 2015, 27, 320–329. [Google Scholar]
- Monte, M.; Impallari, F.M.; Sala, G.; De Michele, A.; Sottile, F. The Italian Plum Rootstock Trial: Results for Sicilian Environmental Conditions. Acta Hortic. 2010, 874, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Kell, S.; Ford-Lloyd, B.; Dulloo, E.; Toledo, Á. Toward the systematic conservation of global crop wild relative diversity. Crop Sci. 2012, 52, 774–785. [Google Scholar] [CrossRef]
- Okie, W.R. Introgression of Prunus species in plum. N. Y. Fruit Q. 2006, 14, 29–37. [Google Scholar]
- García-Gómez, B.E.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular bases of fruit quality in Prunus species: An integrated genomic, transcriptomic, and metabolic review with a breeding perspective. Int. J. Mol. Sci. 2021, 22, 1–38. [Google Scholar] [CrossRef]
- Castro, S.; DeJong, T.M. Developing new prune cultivars for the California dried prune industry. Acta Hortic. 2021, 1322, 13–18. [Google Scholar] [CrossRef]
- Ryugo, K. Stone Fruits. In Fruitculture. Its Science and Art; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1988. [Google Scholar]
- Aranzana, M.J.; Decroocq, V.; Dirlewanger, E.; Eduardo, I.; Gao, Z.S.; Gasic, K.; Iezzoni, A.; Jung, S.; Peace, C.; Prieto, H.; et al. Prunus genetics and applications after de novo genome sequencing: Achievements and prospects. Hortic. Res. 2019, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehder, A. Manual of Cultivated Trees and Shrubs Hardy in North America Exclusive of the Subtropical and Warmer Temperate Regions, 2nd ed.; MacMillan: New York, NY, USA, 1940. [Google Scholar]
- Shaw, J.; Small, R. Addressing the “Hardest Puzzle in American Pomology”: Phylogeny of Prunus Sect. Prunocerasus (Rosaceae) based on seven noncoding chloroplast DNA regions. Am. J. Bot. 2004, 91, 985–996. [Google Scholar] [PubMed] [Green Version]
- Bortiri, E.; Heuvel, B.V.; Potter, D. Phylogenetic analysis of morphology in Prunus reveals extensive homoplasy. Plant Syst. Evol. 2006, 259, 53–71. [Google Scholar] [CrossRef]
- Watkins, R. Cherry, plum, peach, apricot and almond. Prunus spp. In Evolution of Crop Plants; Simmonds, N.W., Ed.; Longman: London, UK, 1976; pp. 242–247. [Google Scholar]
- Aradhya, M.K.; Weeks, C.; Simon, C.J. Molecular characterization of variability and relationships among seven cultivated and selected wild species of Prunus L. using amplified fragment length polymorphism. Sci. Hortic. (Amst.) 2004, 103, 131–144. [Google Scholar] [CrossRef]
- Depypere, L.; Chaerle, P.; Breyne, P.; Vander Mijnsbrugge, K.; Goetghebeur, P. A combined morphometric and AFLP based diversity study challenges the taxonomy of the European members of the complex Prunus L. section Prunus. Plant Syst. Evol. 2009, 279, 219–231. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Turcsán, A.; Depypere, L.; Steenackers, M. Variance, genetic control, and spatial phenotypic plasticity of morphological and phenological traits in Prunus spinosa and its large fruited forms (P. x fruticans). Front. Plant Sci. 2016, 7, 1641. [Google Scholar] [CrossRef] [Green Version]
- Hodel, R.G.I.; Zimmer, E.; Wen, J. A phylogenomic approach resolves the backbone of Prunus (Rosaceae) and identifies signals of hybridization and allopolyploidy. Mol. Phylogenet. Evol. 2021, 160, 107118. [Google Scholar] [CrossRef]
- Okie, W.R. Plum crazy: Rediscovering our lost Prunus resources. HortScience 2001, 36, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Gaši, F.; Sehic, J.; Grahic, J.; Hjeltnes, S.H.; Ordidge, M.; Benedikova, D.; Blouin-Delmas, M.; Drogoudi, P.; Giovannini, D.; Höfer, M.; et al. Genetic assessment of the pomological classification of plum Prunus domestica L. accessions sampled across Europe. Genet. Resour. Crop Evol. 2020, 67, 1137–1161. [Google Scholar] [CrossRef]
- Horvath, A.; Balsemin, E.; Barbot, J.C.; Christmann, H.; Manzano, G.; Reynet, P.; Laigret, F.; Mariette, S. Phenotypic variability and genetic structure in plum (Prunus domestica L.), cherry plum (P. cerasifera Ehrh.) and sloe (P. spinosa L.). Sci. Hortic. (Amst.) 2011, 129, 283–293. [Google Scholar] [CrossRef]
- Woldring, H. On the origin of plums: A study of sloe, damson, cherry plum, domestic plums and their intermediate forms. Palaeohistoria 1997, 39, 535–562. [Google Scholar]
- Pollmann, B.; Jacomet, S.; Schlumbaum, A. Morphological and genetic studies of waterlogged Prunus species from the Roman vicus Tasgetium (Eschenz, Switzerland). J. Archaeol. Sci. 2005, 32, 1471–1480. [Google Scholar] [CrossRef]
- Ucchesu, M.; Sarigu, M.; Del Vais, C.; Sanna, I.; d’Hallewin, G.; Grillo, O.; Bacchetta, G. First finds of Prunus domestica L. In Italy from the Phoenician and Punic periods (6th–2nd centuries bc). Veg. Hist. Archaeobot. 2017, 26, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.; Olrik, D.C. A morphometric analysis of Prunus spinosa, P. Domestica ssp. Insititia, and their putative hybrids in Denmark. Nord. J. Bot. 2001, 21, 349–363. [Google Scholar] [CrossRef]
- Gómez, E.; Ledbetter, C.A. Comparative study of the aromatic profiles of two different plum species: Prunus salicina Lindl and Prunus simonii L. J. Sci. Food Agric. 1994, 65, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wei, X.; Liu, W.; Liu, N.; Zhang, Y.; Xu, M.; Liu, S.; Zhang, Y.; Ma, X.; Dong, W. The genetic relationship and structure of some natural interspecific hybrids in Prunus subgenus Prunophora, based on nuclear and chloroplast simple sequence repeats. Genet. Resour. Crop Evol. 2018, 65, 625–636. [Google Scholar] [CrossRef]
- Milošević, T.; Miloševíc, N. Plum (Prunus spp.) breeding. In Advances in Plant Breeding Strategies: Fruits: Volume 3; Al-Khayri, J., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2018; pp. 165–215. [Google Scholar]
- Neumüller, M. Fundamental and applied aspects of plum (Prunus domestica) breeding. Fruit Veg. Cereal Sci. Biotechnol. 2011, 5, 139–156. [Google Scholar]
- Liu, C.; Feng, C.; Peng, W.; Hao, J.; Wang, J.; Pan, J.; He, Y. Chromosome-level draft genome of a diploid plum (Prunus salicina). Gigascience 2020, 9, giaa130. [Google Scholar] [CrossRef]
- Hartmann, W.; Neumüller, M. Plum Breeding. In Breeding Plantation Tree Crops: Temperate Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2009; pp. 161–232. [Google Scholar]
- Okie, W.; Weinberger, J.H. Fruit Breeding: Tree and Tropical Fruits; Janick, J.M.J., Ed.; Wiley: New York, NY, USA, 1996. [Google Scholar]
- Reales, A.; Sargent, D.J.; Tobutt, K.R.; Rivera, D. Phylogenetics of Eurasian plums, Prunus L. section Prunus (Rosaceae), according to coding and non-coding chloroplast DNA sequences. Tree Genet. Genomes 2010, 6, 37–45. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World, 3rd ed.; Oxford University Press: New York, NY, USA, 2000; Volume 316. [Google Scholar]
- Kazija, D.H.; Jelačić, T.; Vujević, P.; Milinović, B.; Čiček, D.; Biško, A.; Pejić, I.; Šimon, S.; Žulj Mihaljević, M.; Pecina, M.; et al. Plum germplasm in Croatia and neighboring countries assessed by microsatellites and DUS descriptors. Tree Genet. Genomes 2014, 10, 761–778. [Google Scholar] [CrossRef]
- Norton, M.V.; Krueger, W.H. Growing Prunes (Dried Plums) in California: An Overview. ANR Publ. 2007, 7, 8264. [Google Scholar]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and Heat Requirements of Temperate Stone. Agronomy 2020, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- Janick, J. The Origins of Fruits, Fruit Growing, and Fruit Breeding. Plant Breed. Rev. 2005, 25, 255–321. [Google Scholar]
- Hartmann, W. Three new sharka resistant (hypersensitive) plum cultivars from Hohenheim plum breeding programme. Acta Hortic. 2019, 1260, 9–11. [Google Scholar] [CrossRef]
- Doyle, J.F.; Debuse, C.J.; Dejong, T.M. Prune varieties and their origins. In Prune Production Manual; Buchner, R., Ed.; Agriculture and Natural Resources Publication; University of California: Davis, CA, USA, 2012; pp. 75–92. [Google Scholar]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Wang, J.; Yang, W.; Tao, Y.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef]
- Byrne, D.H. Inbreeding, coancestry, and founding clones of Japanese-type plums of California and the southeastern United States. J. Am. Soc. Hortic. Sci. 1989, 114, 699–705. [Google Scholar]
- Guerra, M.E.; Rodrigo, J. Japanese plum pollination: A review. Sci. Hortic. (Amst.) 2015, 197, 674–686. [Google Scholar] [CrossRef]
- Radice, S.; Giordani, E.; Nencetti, V.; Bellini, E. Phenological expression in Prunus salicina Lindl. Genotypes and its relation with insect attraction and pollination. Acta Hort. 2010, 874, 151–156. [Google Scholar] [CrossRef]
- Myracle, A.D.; Castonguay, Z.J.; Elwell, A.; Moran, R.E. Fruit quality and consumer acceptability of three plum types and 14 plum cultivars grown in Maine for a local market. HortTechnology 2018, 28, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Fogle, H.W. Plum Improvement in the United States. Acta Hortic. 1978, 74, 35–40. [Google Scholar] [CrossRef]
- Bellini, E. Cultivar. In Il susino. Frutticoltura anni 80; Baldini, E., Scaramuzzi, F., Eds.; Reda: Firenze, Italy, 1980; pp. 9–73. [Google Scholar]
- Blažek, J. A survey of the genetic resources used in plum breeding. Acta Hortic. 2007, 734, 31–45. [Google Scholar] [CrossRef]
- Nybom, H.; Lācis, G. Recent large-scale genotyping and phenotyping of plant genetic resources of vegetatively propagated crops. Plants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Weise, S.; Oppermann, M.; Maggioni, L.; Van Hintum, T.; Knupffer, H. EURISCO: The European search catalogue for plant genetic resources. Nucleic Acids Res. 2017, 45, D1003–D1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benediková, D.; Giovannini, D. Review on genetic resources in the ECPGR prunus working group. Acta Hortic. 2012, 981, 43–51. [Google Scholar] [CrossRef]
- Zanetto, A.; Maggioni, L.; Tobutt, K.R.; Dosba, F. Prunus genetic resources in Europe: Achievement and perspectives of a networking activity. Genet. Resour. Crop Evol. 2002, 49, 331–337. [Google Scholar] [CrossRef]
- Maggioni, L.; Lipman, E. (Compiler) Report of a Working Group on Prunus. In Proceedings of the Seventh Meeting of the Working Group on Prunus, Larnaca, Cyprus, 1–3 December 2005; ECPGR (Ed.) Bioversity International: Rome, Italy, 2006. [Google Scholar]
- Maggioni, L.; Lateur, M.; Balsemin, E.; Lipman, E. Report of a Working Group on Prunus. In Proceedings of the Eighth Meeting, Forlì, Italy, 7–9 September 2010; ECPGR, Bioversity International: Rome, Italy, 2011. [Google Scholar]
- ECPGR. Identification of a Representative Set of Prunus Domestica Accessions of European Origin, Well Documented and Characterized, to be Included into the AEGIS System. 2016. Available online: https://www.ecpgr.cgiar.org/resources/ecpgr-publications/publication/identification-of-a-representative-set-of-prunus-domestica-accessions-of-european-origin-well-documented-and-characterized-to-be-included-into-the-aegis-system-prundoc-2016 (accessed on 21 November 2021).
- Ordidge, M.; Lācis, G.; Delmas, M.; Höfer, M.; Drogoudi, P.; Hjeltnes, S.H.; Kaldmäe, H.; Toldam-andersen, T.; Békefi, Z.; Fernández, F.F.; et al. Testing, Use and Alignment of Genetic Data to Distinguish Unique and Characterized Accessions in Prunus (Prunus Alignment). Activity Report. 2021. Available online: https://www.ecpgr.cgiar.org/resources/ecpgr-publications/publication/testing-use-and-alignment-of-genetic-data-to-distinguish-unique-and-characterized-accessions-in-prunus-prunus-alignment-1-january-2018-30-april-2020-2021 (accessed on 23 November 2021).
- Hjeltnes, S.H.; Giovannini, D.; Blouin, M.; Benedikova, D.; Drogoudi, P.; Höfer, M.; Lacis, G.; Ognjanov, V.; Lateur, M.; Engels, J.M.; et al. PRUNDOC—A project to define accessions for the European Collection. Acta Hortic. 2017, 1175, 19–24. [Google Scholar] [CrossRef]
- Nybom, H.; Giovannini, D.; Ordidge, M.; Hjeltnes, S.H.; Grahić, J.; Gaši, F. ECPGR recommended SSR loci for analyses of European plum (Prunus domestica) collections. Genet. Resour. 2020, 1, 40–48. [Google Scholar] [CrossRef]
- Sehic, J.; Nybom, H.; Hjeltnes, S.H.; Gaši, F. Genetic diversity and structure of Nordic plum germplasm preserved ex situ and on-farm. Sci. Hortic. (Amst.) 2015, 190, 195–202. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Errea, P.; Miranda, C.; Santesteban, L.G.; Pina, A. Genetic diversity of Spanish Prunus domestica L. germplasm reveals a complex genetic structure underlying. PLoS ONE 2018, 13, e0195591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ternjak, T.; Barreneche, T.; SŠušek, A.; SŠisko, M.; Ivančič, A.; Quero-Garcıá, J. The Slovenian traditional plum germplasm and its diversity compared to the French National Plum Collection. Acta Hortic. 2021, 1322, 69–76. [Google Scholar] [CrossRef]
- Milošević, T.; Miloševíc, N. Phenotypic diversity of autochthonous European (Prunus domestica L.) and Damson (Prunus insititia L.) plum accessions based on multivariate analysis. Hortic. Sci. 2012, 39, 8–20. [Google Scholar] [CrossRef]
- Ivanova, D.; Dimkova, S.; Vitanova, I.; Marinova, N. Yield and chemical composition of seven local plum varieties. Acta Hortic. 2009, 825, 471–474. [Google Scholar] [CrossRef]
- Bozhkova, V. Plum genetic resources and breeding. AgroLife Sci. J. 2013, 2, 83–88. [Google Scholar]
- Makovics-Zsohár, N.; Tóth, M.; Surányi, D.; Kovács, S.; Hegedűs, A.; Halász, J. Simple sequence repeat markers reveal Hungarian plum (Prunus domestica L.) germplasm as a valuable gene resource. HortScience 2017, 52, 1655–1660. [Google Scholar] [CrossRef]
- Butac, M.; Botu, M.; Militaru, M.; Chitu, E.; Plopa, C.; Sumedrea, M.; Sumedrea, D. Old autochthonous Romanian plum genotypes—A source for breeding program. Acta Hortic. 2017, 1175, 29–36. [Google Scholar] [CrossRef]
- Butac, M.; Botu, M.; Militaru, M.; Mazilu, C.; Dutu, I.; Nicolae, S. Plum Germplasm Resources and Breeding in Romania. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2019, 73, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Basso, M.; Faccioli, F. Le Principali Prugne Coltivate in Italia; Istituto Coltivazioni Arboree: Bologna, Italy, 1978; Volume 163. [Google Scholar]
- Nencetti, V.; Picardi, E. Susino. In Atlante dei Fruttiferi Autoctoni Italiani; Fideghelli, C., Ed.; MIPAAF-CREA: Rome, Italy, 2016; pp. 1575–1651. ISBN 978-88-99595-35-7. [Google Scholar]
- Manco, R.; Basile, B.; Capuozzo, C.; Scognamiglio, P.; Forlani, M.; Rao, R.; Corrado, G. Molecular and phenotypic diversity of traditional European plum (Prunus domestica L.) germplasm of Southern Italy. Sustainability 2019, 11, 4112. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, D.; Antonucci, F.; Talento, C.; Ciccoritti, R. Chemical characterization in the selection of Italian autochthonous genotypes of plum. Sci. Hortic. (Amst.) 2021, 281, 109922. [Google Scholar] [CrossRef]
- Gharbi, O.; Wünsch, A.; Rodrigo, J. Characterization of accessions of “Reine Claude Verte” plum using Prunus SRR and phenotypic traits. Sci. Hortic. (Amst.) 2014, 169, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, D.; Baraket, G.; Perez, V.; Ben Mustapha, S.; Salhi-Hannachi, A.; Hormaza, J.I. Analysis of self-incompatibility and genetic diversity in diploid and hexaploid plum genotypes. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fernandez i Marti, A.; Castro, S.; DeJong, T.M.; Dodd, R.S. Evaluation of the S-locus in Prunus domestica, characterization, phylogeny and 3D modelling. PLoS ONE 2021, 16, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.F.; Volk, G.M.; Gardner, C.; Gore, M.A.; Simon, P.W.; Smith, S. Sustaining the future of plant breeding: The critical role of the USDA-ARS national plant germplasm system. Crop Sci. 2018, 58, 451–468. [Google Scholar] [CrossRef] [Green Version]
- Wight, W.F. The Varieties of Plums Derived from Native American Species; U.S. Department of Agriculture: Singapore, 1915; Volume 172.
- Karp, D.; Breeding, B.S.P. Luther Burbank’s Plums. HortScience 2015, 50, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Sherman, W.B.; Rouse, R.E. Characteristics of Plums from the University of Florida Breeding Program. Pro. Fla. State Hort. Soc. 2001, 114, 30–32. [Google Scholar]
- Della Strada, G.; Fideghelli, C. Le Varietà dei Fruttiferi Introdotte nel Mondo dal 1980 al 2008; MiPAAF, CRA-FRU: Roma, Italy, 2011. [Google Scholar]
- Byrne, D.H. Trends in Stone Fruit Cultivar Development. Horttechnology 2005, 15, 494–500. [Google Scholar] [CrossRef]
- Coe, M.T.; Evans, K.M.; Gasic, K.; Main, D. Plant breeding capacity in U.S. public institutions. Crop Sci. 2020, 60, 2373–2385. [Google Scholar] [CrossRef]
- Scorza, R.; Ravelonandro, M.; Callahan, A.; Zagrai, L.; Polak, J.; Malinowski, T.; Cambra, M.; Levy, L.; Damsteegt, V.; Krška, B.; et al. Honeysweet (C5), the first genetically engineered plum pox virus-resistant plum (Prunus domestica L.) cultivar. HortScience 2016, 51, 601–603. [Google Scholar] [CrossRef] [Green Version]
- Paunovic, S.A.; Glisic, I.S.; Karaklajic-Stajic, Z.; Milosevic, N.; Jevremovic, D. Performance of “Lana”, a new plum cultivar developed at Fruit Research Institute, Cacak. Acta Hortic. 2021, 1322, 25–32. [Google Scholar] [CrossRef]
- Lugli, S.; Correale, R.; Grandi, M.; Sansavini, S.; Bertolazzi, M.; Pallotti, G.; Tartarini, S. Bologna University’s plum breeding program. Acta Hortic. 2019, 1260, 23–28. [Google Scholar] [CrossRef]
- Ruiz, D.; Cos, J.; Nicolás-Almansa, M.; Egea, J.; Garcıá, F.; Carrillo, A.; Rubio, M.; López, D.; Salazar, J.A.; Guevara, A. New promising Japanese plum cultivars for warm areas from CEBAS-CSIC/IMIDA breeding programme. Acta Hortic. 2021, 1322, 55–60. [Google Scholar] [CrossRef]
- Botu, M. Stone fruit germplasm resource and exploitation and links with ECPGR and EUFRIN. In Proceedings of the Joint Meeting of European COST & ISHS—1st International Congress for Bacterial Diseases of Stone Fruits and Nuts, Zurich, Switzerland, 14–17 February 2012. [Google Scholar]
- Butac, M.; Bozhkova, V.; Zhivondov, A.; Milošević, N.; Bellini, E.; Nencetti, V.; Blazek, J.; Lafarque, B.; Kaufmane, E.; Gravite, I.; et al. Overview of plum breeding in Europe. Acta Hortic. 2013, 981, 91–98. [Google Scholar] [CrossRef]
- Novotna, I.; Dankova, V.; Pravcova, G.; Vavra, R. Fruit characteristic of new plum cultivars bred in Czech Republic. Acta Hortic. 2021, 1322, 109–112. [Google Scholar] [CrossRef]
- Community Plant Variety Office (CPVO). Annual Report 2020 Annex; Publications Office of the European Union: Luxembourg, 2021; Available online: www.cpvo.europa.eu (accessed on 14 November 2021).
- Dalbó, M.A.; Menezes-Netto, A.C.; Bruna, E.D.; Thomazi-Kleina, H.; May-De-Mio, L.L. Advances in plum breeding for resistance to Xylella fastidiosa in Brazil. Acta Hortic. 2021, 1322, 19–24. [Google Scholar] [CrossRef]
- Zurawicz, E.; Pruski, K.; Szymajda, M.; Lewandowski, M.; Seliga, L.; Malinowski, T. Prunus domestica ‘Jojo’—Good Parent for Breeding of New Plum Cultivars Resistant to Plum pox Virus. J. Agric. Sci. 2013, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Glisic, I.; Karaklajic-Stajic, Z.; Paunovic, S.A.; Lukic, M. Plum cultivars Zlatka and Pozna Plava (Prunus domestica L.) bred at the Fruit Research Institute in Èaèak. Hortic. Sci. 2016, 43, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Neshev, N.; Nesheva, M.; Akova, V. Agronomic and fruit quality characteristics of the Bulgarian plum cultivar “Pagane”. Acta Hortic. 2021, 1322, 33–39. [Google Scholar] [CrossRef]
- Milošević, N.; Glišić, I.S.; Đorđević, M.; Radičević, S.; Jevremović, D. An overview of plum breeding at Fruit Research Institute, Čačak. Acta Hortic. 2021, 1322, 7–12. [Google Scholar] [CrossRef]
- Nencetti, V.; Morelli, D. Il miglioramento genetico del susino in Italia. Frutticoltura 2014, 5, 14–18. [Google Scholar]
- Pirazzini, P.; Mezzetti, B.; Insero, O.; Bellini, E.; Nencetti, V. Susino Liste varietali. Terra E Vita 2003, 7, 75–80. [Google Scholar]
- Liverani, A.; Sirri, S.; Nencetti, V. Susino Liste Varietali. Terra E Vita 2013, 5, 24–29. [Google Scholar]
- Fideghelli, C.; Ancarani, V.; Antonacci, D. Analisi e rinnovamento delle specie e delle varietà frutticole. Frutticoltura 2018, 82, 54–63. [Google Scholar]
- Lugli, S.; Correale, R.; Grandi, M.; Sansavini, S. Breeding high-quality plums at Bologna University’s CMVF-DCA. Acta Hortic. 2010, 874, 69–76. [Google Scholar] [CrossRef]
- Byrne, D.H.; Sherman, W.B.; Bacon, T.A. Stone fruit genetic pool and its exploitation for growing under warm winter conditions. In Temperate Fruit Crops in Warm Climates; Erez, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 157–230. [Google Scholar]
- Nicolas-Almansa, M.; Salazar, J.A.; Rubio, M.; Guevara, A.; Carrillo, A.; Garcia, F.; Cos, J.; Martinez-Gomez, P.; Ruiz, D. Inheritance and correlation of main phenological and pomological traits in Japanese plum (Prunus salicina Lindl.). Acta Hortic. 2021, 1322, 61–67. [Google Scholar] [CrossRef]
- Botu, I.; Botu, M. Limits and perspectives in plum cultivar breeding using conventional methods. Acta Hortic. 2007, 734, 321–325. [Google Scholar] [CrossRef]
- Zaremuk, R.S.; Dolya, Y.A.; Kopnina, T.A.; Kochubey, A.A. Efficiency of the breeding use of the genus Prunus L. biodiversity. E3S Web Conf. 2021, 254, 01027. [Google Scholar] [CrossRef]
- Petri, C.; Alburquerque, N.; Faize, M.; Scorza, R.; Dardick, C. Current achievements and future directions in genetic engineering of European plum (Prunus domestica L.). Transgenic Res. 2018, 27, 225–240. [Google Scholar] [CrossRef] [Green Version]
- International Union for the Protection of New Varieties of Plants (UPOV). Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability, European Plum (Prunus Domestica). 2002. Available online: https://www.upov.int/en/publications/tg-rom/tg041/tg_41_5.pdf (accessed on 3 December 2021).
- Neumüller, M.; Dittrich, F.; Holzapfel, C. Recent achievements and future challenges in breeding for European plum cultivars: Finding the suitable genetic resources for important traits. Acta Hortic. 2021, 1322, 1–5. [Google Scholar] [CrossRef]
- Register of New Fruit and Nut Varieties—Lists 40–50. Hort. Sci. Volume 35; 37; 39; 41; 43; 45; 47; 49; 51; 53; 55. 2000–2020. Available online: https://journals.ashs.org/hortsci/view/journals/hortsci/55/7/article-p1164.xml (accessed on 11 November 2021).
- Koebner, R.; Ortiz, R. Fishing in the gene pool—How useful was the catch? Plant Genet. Resour. 2013, 11, 283–287. [Google Scholar] [CrossRef]
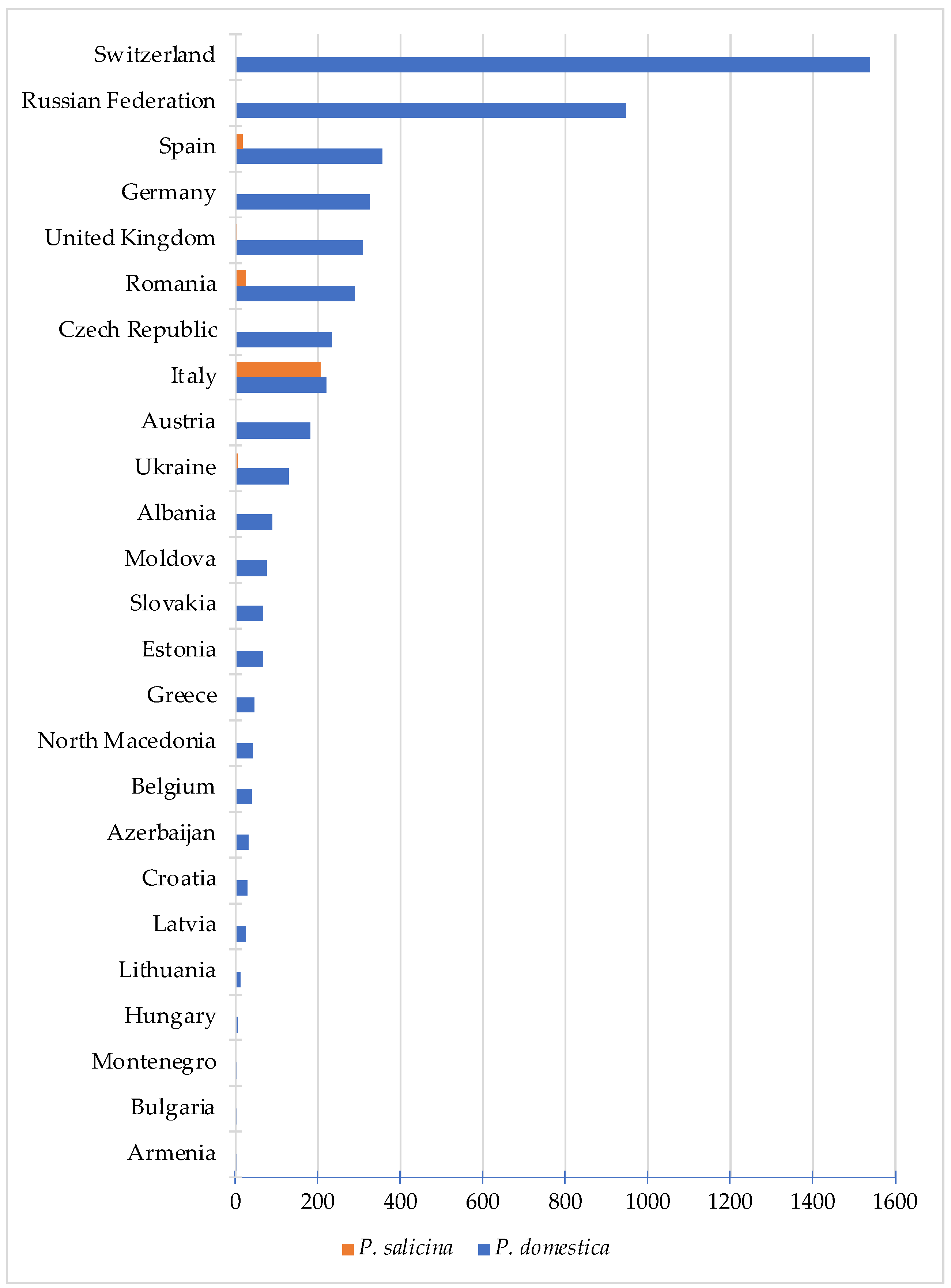
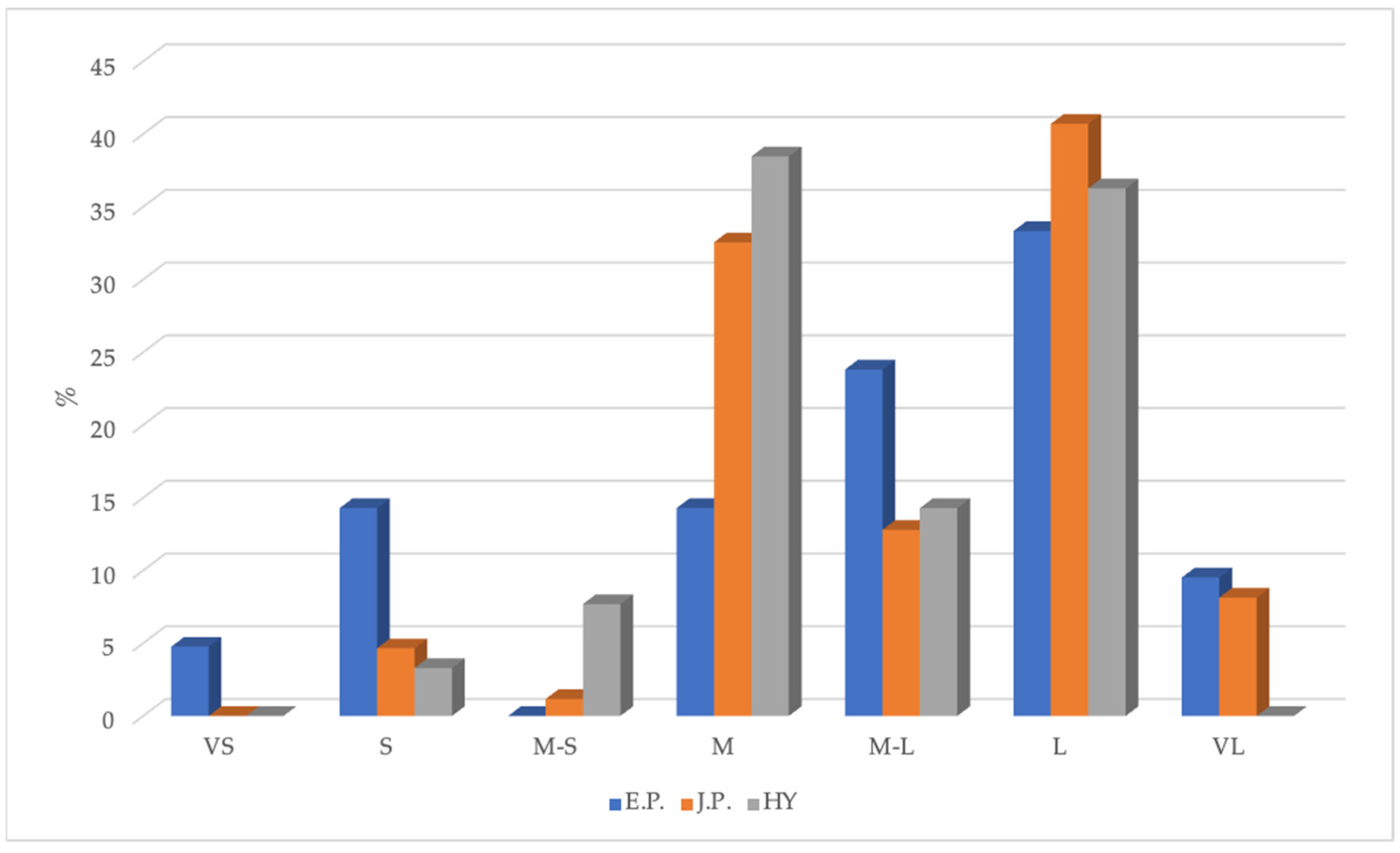

{kind=link}
{kind=link}
| Related Species | Positive Traits | Negative Traits | Ref.: | |
|---|---|---|---|---|
| Tree | Fruit | |||
| P. simonii | flat fruit shape, small stone, flesh firmness, acidity, aroma | [55,56] | ||
| P. cathayana | sweetness | [56] | ||
| P. limeixing | late-flowering or frost-resistance | pubescent fruit surface, flesh firmness, aroma | [56] | |
| P. mume | early blooming | [59] | ||
| P. cerasifera | cold hardiness good productivity, resistance to bacterial spot, drought and heat, early maturity, and tolerance to unfavorable conditions | medium to small fruit size | [35,60] | |
| P. spinosa | drought resistance, cold hardiness dwarfing | blue fruit color | [60] | |
| P. microcarpa | drought resistance; heat resistance | [60] | ||
| P. americana | cold hardiness; tough skin | suckering | [9,60] | |
| P. angustifolia | low chilling requirement; resistance to bacterial spot | suckering, limited tolerance to plum leaf scald; small fruit size | [20,48,60] | |
| P. ussuriensis | cold hardiness | [60] | ||
| P. besseyi | late bloom, frost resistance, resistance to crown gall | sensitivity to Monilinia spp. | [20,48,60] | |
| P. maritima | frost resistance, late bloom, high heat requirement; salt tolerance | [48,60] | ||
| P. alleghaniensis | resistance to crown gall | small fruit size; bitterness | [20,48] | |
| P. geniculata | drought tolerant, low chill | small fruit size | [20,48] | |
| P. hortulana | resistance to bacterial spot | bright skin color | small fruit size | [20,48] |
| P. munsoniana | productive | good fruit quality | [20] | |
| P. subcordata | drought tolerance | high chill requirement | [20,48] | |
| P. nigra | cold hardiness | [20] | ||
| P. umbellata | resistance to crown gall | poor fruit quality | [20,28] | |
| P. texana | precocity | fair quality | [4,48] | |
| Group and Species | 1989 | 1999 | 2009 | 2017 | 2017/1989 |
|---|---|---|---|---|---|
| % | |||||
| European: | |||||
| P. domestica (European plum) | 0 | 141 | 154 | 193 | |
| P. bokhariensis | 3 | 3 | 2 | 2 | |
| P. spinosa | 21 | 5 | 8 | 23 | |
| P. cerasifera | 14 | 32 | 45 | 66 | |
| P. cerasifera var. divaricata | 27 | ||||
| P. insititia | 0 | 1 | 3 | 0 | |
| Subtotal | 38 | 182 | 212 | 311 | 818 |
| Asian: | |||||
| P. salicina (Japanese plum) | 92 | 40 | 63 | 77 | |
| P. salicina var. mandshurica | 1 | ||||
| P. salicina var. salicina | 3 | ||||
| P. simonii * | 2 | 3 | 3 | 8 | |
| P. hybrid (plumcot, aprium, pluot, etc.) | 59 | 130 | |||
| Subtotal | 94 | 102 | 66 | 219 | 233 |
| New World Plums: | |||||
| P. americana | 1 | 4 | 11 | ||
| P. rivularis | 0 | 0 | 0 | 3 | |
| P. hortulana * | 3 | 0 | 1 | 6 | |
| P. mexicana | 15 | 2 | 2 | 3 | |
| P. maritima * | 50 | 2 | 21 | 3 | |
| P. geniculate * | 0 | 0 | 0 | 1 | |
| P. alleghaniensis * | 0 | 3 | 2 | 0 | |
| P. angustifolia | 2 | 15 | 4 | 20 | |
| P. gracilis | 0 | 0 | 0 | 0 | |
| P. munsoniana * | 0 | 1 | 2 | 0 | |
| P. nigra | 0 | 1 | 0 | 1 | |
| P. umbellata | 0 | 0 | 0 | 7 | |
| Subtotal | 71 | 28 | 32 | 55 | 77 |
| Other: | |||||
| P. subcordata | 26 | 12 | 7 | 12 | |
| P. texana | 0 | 0 | 0 | 2 | |
| P. bifrons | 0 | 0 | 0 | 4 | |
| P. pumila | 1 | 0 | 1 | 0 | |
| P. pumula var. besseyi | 5 | 1 | 0 | 4 | |
| P. microcarpa | 0 | 0 | 0 | 3 | |
| P. tomentosa | 55 | 6 | 9 | 11 | |
| P. andersonii | 0 | 3 | 0 | 4 | |
| P. fremontii | 1 | 2 | 0 | 2 | |
| P. × cistena (P. cerasifera × P. pumila) | 1 | 0 | 0 | 1 | |
| Subtotal | 89 | 24 | 17 | 43 | 48 |
| Total | 292 | 336 | 327 | 628 | 215 |
| EPz | JPz | Hybr. | TOT. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yr. | Ey | My | Ly | Ey | My | Ly | Ey | My | Ly | Ey | My | Ly | Total |
| 2000 | 2 | 2 | 0 | 0 | 2 | ||||||||
| 2002 | 3 | 2 | 2 | 3 | 5 | 1 | 5 | 10 | 1 | 16 | |||
| 2004 | 1 | 1 | 1 | 11 | 3 | 2 | 7 | 1 | 2 | 19 | 5 | 5 | 29 |
| 2006 | 1 | 1 | 1 | 9 | 7 | 4 | 4 | 2 | 14 | 10 | 5 | 29 | |
| 2008 | 1 | 2 | 1 | 2 | 2 | 1 | 1 | 3 | 2 | 4 | 7 | 4 | 15 |
| 2010 | 3 | 3 | 3 | 2 | 17 | 7 | 5 | 20 | 10 | 35 | |||
| 2012 | 1 | 2 | 3 | 2 | 3 | 2 | 3 | 5 | 5 | 13 | |||
| 2014 | 1 | 13 | 8 | 1 | 13 | 8 | 22 | ||||||
| 2016 | 7 | 1 | 1 | 2 | 1 | 2 | 1 | 2 | 11 | 2 | 15 | ||
| 2018 | 3 | 7 | 3 | 3 | 7 | 3 | 13 | ||||||
| 2020 | 3 | 3 | 1 | 2 | 3 | 5 | 1 | 9 | |||||
| Total | 3 | 14 | 4 | 38 | 31 | 17 | 20 | 48 | 23 | 61 | 93 | 44 | 198 |
| Pub. | Priv. | Pub. | Priv. | USA | Other Count. | |
|---|---|---|---|---|---|---|
| Year | N° | % | N° | |||
| 2000 | 2 | 100 | 0 | 2 | 0 | |
| 2002 | 5 | 11 | 31.3 | 68.7 | 15 | 1 |
| 2004 | 4 | 25 | 13.8 | 86.2 | 29 | 0 |
| 2006 | 2 | 27 | 6.9 | 93.1 | 29 | 0 |
| 2008 | 6 | 9 | 40.0 | 60.0 | 9 | 6 |
| 2010 | 1 | 34 | 2.9 | 97.1 | 32 | 3 |
| 2012 | 13 | 0.0 | 100.0 | 13 | 0 | |
| 2014 | 22 | 0.0 | 100.0 | 22 | 0 | |
| 2016 | 3 | 12 | 20.0 | 80.0 | 9 | 6 |
| 2018 | 13 | 0.0 | 100.0 | 13 | 0 | |
| 2020 | 9 | 0.0 | 100.0 | 8 | 1 | |
| Total | 23 | 175 | 11.6 | 88.4 | 181 | 17 |
| Cultivar | Plum Type(z) | Bacterial Spot | Bacterial Canker | Leaf Scald | Black Knot | PPV |
|---|---|---|---|---|---|---|
| Spring Satin | Hy | R | R | T | ||
| Gulfbeauty | JP | T | R | |||
| Gulfblaze | JP | T | R | |||
| Gulfrose | JP | T | R | |||
| Mann | JP | R | ||||
| Queen Garnet | JP | R | ||||
| Ruby Queen | JP | R | R | |||
| Vampire | JP | T | T | |||
| Honey Sweet | EP | R | R | |||
| Valerie | EP | R | R | |||
| Vandor | EP | R | R | |||
| Vanette | EP | R | R | |||
| Vibrant | EP | R | R | |||
| Violette | EP | R | R |
| Problem/Researched Trait | Potential Donors with Improved/High Trait | References | |
|---|---|---|---|
| European Plums | Japanese Plums | ||
| Primary/Traditional | |||
| Early blooming | ‘J2N-127′, ‘Graf Bruhl’, ‘Lutzelsachser Fruhzwetsche’ | [37,134] | |
| Late Blooming | ‘Blue Bell’, ‘Italian Prune’, ‘Pitestean’ | [60,134] | |
| Low chilling requirement | ‘Gulfblaze’, ‘Gulfbeauty’.‘Victoria Myrtea’ | [107,114] | |
| Early maturing | ‘J15S-22′, ‘J16N-95′, ‘Ruth Gerstetter’ | ‘Spring Satin’, ‘Lucia Myrtea’, ‘Victoria Myrtea’ | [35,37,114,129] |
| Late maturing | ‘Elena’, ‘Reine Claude de Bavay’, ‘Presenta’ | ‘Fallette’, ‘Holiday’, ‘Ruby Queen’ | [35,60,134] |
| Climatic adaptation | ‘Prune d’Agen’, ‘Italian Prune’, ‘Stanley’, ‘German Prune’ | [58] | |
| Fruit size | ‘J17S-30′, ‘President’, ‘Jubileum’, ‘Tophit’, ‘Haganta’, ‘Lana’, ‘Pagane’ ‘Grossa di Felisio’, ‘Kabardinskaya Rannyaya’ | ‘Plumcandy XIV’, ‘Suplum Fiftyfour’ | [37,60,111,122,124,128] |
| Self-compatibility | ‘Muir Beauty’, ‘Stanley’, ‘Anna Späth’, ‘Bluefre’, ‘Ialomiza’, ‘Diana’, ‘Andreea’ | ‘Victoria Myrtea’ | [69,96,114,135] |
| Productivity | ‘Muir Beauty’, ‘Grase de Pesteana’, ‘Stanley’, ‘Anna Späth’, ‘Bluefre’, ‘Standard’, ‘Grase de Becs’, ‘Čačanska Lepotica’ | [95,96] | |
| Fruit taste and quality | ‘Nada’, ‘Mildora’, ‘Kabardinskaya Rannyaya’, ‘Oneida’, ‘Andreea’, ‘Grase de Becs’ | ‘Hanita’ | [60,96,123,128] |
| High sugar content | ‘F11S-38′, ‘J4N-119′, ‘Sugar Top’, ‘Mildora’ | [37,123,124] | |
| High flesh firmness | ‘Hauszwetsche’, Nordens, ‘Katinka’, ‘Tegera’, ‘Čačanska Lepotica’ | [60,134] | |
| Floral precocity | ‘H13S-58′, ‘I12S-6′ ‘Stanley’, ‘Čačanska Lepotica’, ‘Čačanska Rodna’, ‘Verity’ | [37,60] | |
| Resistance to PPV | ‘Moni’, ‘HoneySweet’, ‘Jojo’, ‘Boranka’, ‘Timočanka’, ‘Mildora’, ‘Krina’, ‘Zlatk’a, ‘Pozna’ ‘Plava’, ‘Nada’, ‘Jofela’, ‘Jolinda’, ‘Jocanta’, ‘Divna’, ‘Petra’, ‘Lana’, ‘Grase de Becs’, ‘Uriase de Sibiu’ | [58,69,95,96,111,120,121,123,135] | |
| Resistance to leaf scald | ‘SC7′ | [119] | |
| New/Additional | |||
| Low fresh to dry fruit weight loss | ‘HoneySweet’, ‘F11S-38′, ‘F11S-38′ | [37,111] | |
| Storage ability | ‘Vampire’ | [136] | |
| Abnormal June fruit drop | ‘Katinka’, ‘Juna’, ‘Moni’, ‘Haroma’ | [135] | |
| Frost tolerance | ‘Franzi’, ‘Hauszwetsche’, ‘Mirabelle de Nancy’, ‘Schönberger’, ‘Italian Prune’, ‘German Prune’ | [60,135] | |
| Winter hardiness | ‘Vengerka Moskovskaya’, ‘Zuysinskaya’, ‘Reine Claude Reform’ | [58,135] | |
| Heat susceptibility (damage in the flesh) | ‘Moni’ | [60,131] | |
| Stone cracking | [135] | ||
| Twin fruits | ‘Hauszwetsche’, ‘Katinka’, ‘Juna’ ‘Čačanska Rodna’ | [58,60,135] | |
| Orange flesh color | ‘Hanita’, Gulfblaze, ‘John W.’, ‘Sugar Top’ | [58,136] | |
| Red flesh color | ‘Lucia Myrtea’, ‘Victoria Myrtea’, ‘Vampire’, ‘Plumred X, Plumsweet (series)’, ‘Suplumfortyseven to Suplumfiftyone (series)’ | [114,136] | |
| Caverns and pectin inclusions | ‘Moni’ | [135] | |
| Freestone pit | ‘Hauszwetsche’, ‘Kirke’s’, ‘Čačanska Lepotica’, ‘Tegera’, ‘Katinka’, ‘Sutter’ | [60,69,134] | |
| Processing suitability(z) | [69] | ||
| Good tree structure(z) | [37] | ||
| Uniformity in fruit maturation(z) | [37] | ||
| Slow fruit softening near harvest time(z) | [37] | ||
| Plum decline: Pseudomonas syringae resistance(z) | [135] | ||
| Drosophila suzukii resistance(z) | [135] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sottile, F.; Caltagirone, C.; Giacalone, G.; Peano, C.; Barone, E. Unlocking Plum Genetic Potential: Where Are We At? Horticulturae 2022, 8, 128. https://doi.org/10.3390/horticulturae8020128
Sottile F, Caltagirone C, Giacalone G, Peano C, Barone E. Unlocking Plum Genetic Potential: Where Are We At? Horticulturae. 2022; 8(2):128. https://doi.org/10.3390/horticulturae8020128
Chicago/Turabian StyleSottile, Francesco, Chiara Caltagirone, Giovanna Giacalone, Cristiana Peano, and Ettore Barone. 2022. "Unlocking Plum Genetic Potential: Where Are We At?" Horticulturae 8, no. 2: 128. https://doi.org/10.3390/horticulturae8020128