
Metabolic Response of Malus domestica Borkh cv. Rubin Apple to Canopy Training Treatments in Intensive Orchards



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
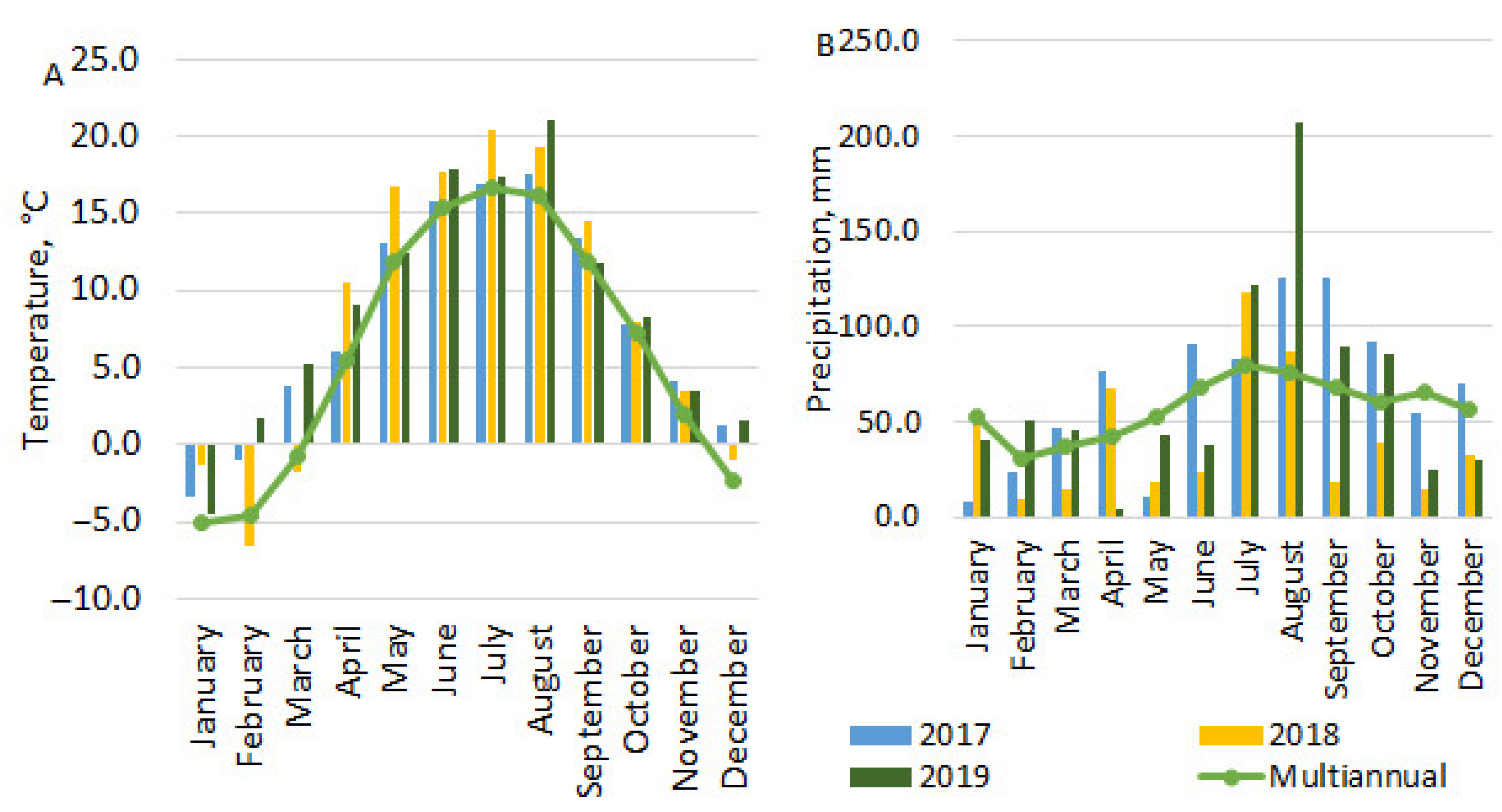
2.1. Meteorological Conditions
2.2. Sample Preparation
2.3. Biochemical Analyses
2.3.1. Assessment of Photosynthetic Pigments Using High-Performance Liquid Chromatography (HPLC)
2.3.2. Assessment of Soluble Sugars Using Ultra-Performance Liquid Chromatography (UPLC)
2.3.3. Bioassay of Total Starch by Colorimetric Method
2.3.4. Bioassay of Total Phenolic Compounds Using Colorimetric Method
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Masouleh, S.S.S.; Sassine, Y.N. Molecular and biochemical responses of horticultural plants and crops to heat stress. Ornam. Hortic. 2020, 26, 148–158. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant Salt Stress: Adaptive Responses, Tolerance Mechanism and Bioengineering for Salt Tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Kamrun Nahar, K.; Alam Md, M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Kooyers, N.J.; Greenlee, A.B.; Colicchio, J.M.; Oh, M.; Blackman, B.K. Replicate altitudinal clines reveal that evolutionary flexibility underlies adaptation to drought stress in annual Mimulus guttatus. New Phytol. 2015, 206, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Bohra, A.; Pandey, A.K.; Pandey, M.K.; Kumar, A. Metabolomics for plant improvement: Status and prospects. Front. Plant Sci. 2017, 8, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saud, S.; Fahad, S.; Yajun, C.; Ihsan, M.Z.; Hammad, H.M.; Nasim, W.; Amanullah, Jr.; Arif, M.; Alharby, H. Effects of Nitrogen Supply on Water Stress and Recovery Mechanisms in Kentucky Bluegrass Plants. Front. Plant Sci. 2017, 8, 983. [Google Scholar] [CrossRef] [Green Version]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [Green Version]
- Mazid, M.; Khan, T.A.; Mohammad, F. Role of secondary metabolites in defense mechanisms of plants. Exp. Biol. Med. 2011, 3, 232–249. [Google Scholar]
- Popa, V.I. Wood bark as valuable raw material for compounds with biological activity. Celul. Hârtie 2015, 64, 5–17. [Google Scholar]
- Ma, D.; Li, Y.; Zhang, J.; Wang, C.; Qin, H.; Ding, H.; Xie, Y.; Guo, T. Accumulation of Phenolic Compounds and Expression Profiles of Phenolic Acid Biosynthesis-Related Genes in Developing Grains of White, Purple, and Red Wheat. Front. Plant Sci. 2016, 7, 528. [Google Scholar] [CrossRef]
- Slatnar, A.; Mikulič-Petkovešek, M.; Veberič, R.; Štampar, F. Research on the involment of phenoloics in the defence of horticultural plants. Acta Agric. Slov. 2017, 107, 183. [Google Scholar] [CrossRef] [Green Version]
- Arici, S.E.; Kafkas, E.; Kaymak, S. Phenolic compounds of apple cultivars resistant or susceptible to Venturia inaequalis. Pharm. Biol. 2014, 52, 904–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolić, M.T.; Krbavčić, I.P.; Vujević, P.; Milinović, B.; Jurčević, I.L.; Vahčić, N. Effects of Weather Conditions on Phenolic Content and Antioxidant Capacity in Juice of Chokeberries (Aronia melanocarpa L.). Polish J. Food. Nutr. Sci. 2017, 67, 67–74. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Zhu, L.; Huang, Y.; Lu, J. Influence of growing season on phenolic compounds and antioxidant properties of grape berries from vines grown in subtropical climate. J. Agric. Food Chem. 2011, 59, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W. Effects of production and processing factors on major fruit and vegetable antioxidants. J. Food Sci. 2005, 70, 11–19. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Dupont, N.; Guillermin, P. Concentrations and characteristics of procyanidins and other phenolics in apples during fruit growth. Phytochemistry 2007, 68, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Liaudanskas, M.; Viškelis, P.; Kviklys, D.; Raudonis, R.; Janulis, V. A comparative study of phenolic content in apple fruits. Int. J. Food Prop. 2015, 18, 945–953. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Ruta, C.; Fortunato, I.M.; Lattanzio, V.M.T.; Linsalata, V.; Cicco, N. Relationship of secondary metabolism to growth in oregano (Origanum vulgare L.) shoot cultures under nutritional stress. Environ. Exp. Bot. 2009, 65, 54–62. [Google Scholar] [CrossRef]
- Treutter, D. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regul. 2001, 34, 71–89. [Google Scholar] [CrossRef]
- Skłodowska, M.; Mikiciński, A.; Wielanek, M.; Kuźniak, E.; Sobiczewski, P. Phenolic profiles in apple leaves and the efficacy of selected phenols against fire blight (Erwinia amylovora). Eur. J. Plant Pathol. 2017, 151, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. Phenolic compounds in apple leaves after infection with apple scab. Biol. Plant 2011, 55, 725–732. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars--metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benyas, E.; Hassanpouraghdam, M.B.; Zehtbsalmasi, S.; Khatamian Oskooei, O.S. Allelopathic effects of Xanthium strumarium L. shoots aqueous extracts on germination, seedlings growth and chlorophyll content of lentil (Lens culinaris Medic.). Rom. Biotechnol. Lett. 2010, 15, 5223–5228. [Google Scholar]
- Zepka, L.Q.; Jacob-Lopes, E.; Roca, M. Catabolism and bioactive properties of chlorophylls. Curr. Opin. Food Sci. 2019, 26, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Alexieva, V.; Ivanov, S.; Sergiev, I.; Karanov, E. Interaction between stresses. Bulg. J. Plant Physiol. 2003, 29, 1–17. [Google Scholar]
- Khanal, N.; Bray, G.E.; Grisnich, A.; Moffatt, B.A.; Gray, G.R. Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema. Salsugineum. Plants 2017, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. J. Nutr. 2004, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Gharras, H. Polythenols: Food sources, properties and applications. Int. J. Food Sci. Technol. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- FAOSTAT. 2020. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 1 January 2022).
- Brons, C.; Olieman, C. Study of the high-performance liquid chromatographic separation of reducing sugars, applied to the determination of lactose in milk. J. Chromatogr. A 1983, 259, 79–86. [Google Scholar] [CrossRef]
- McCleary, B.V.; Gibson, T.S.; Mugford, D.C. Measurement of Total Starch in Cereal Products by Amyloglucosidase-α-Amylase Method: Collaborative Study. J. AOAC Int. 1997, 80, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Roca, M.; Chen, K.; Pérez-Gálvez, A. Chlorophylls. In Handbook on Natural Pigments in Food and Beverages: Industrial Applications for Improving Food Color; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; pp. 125–158. [Google Scholar]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hort. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picinelli, A.; Dapena, E.; Mangas, J.J. Polyphenolic pattern in apple tree leaves in relation to scab resistance: A preliminary study. J. Agric. Food Chem. 1995, 43, 2273–2278. [Google Scholar] [CrossRef] [Green Version]
- Petkovsek, M.M.; Slatnar, A.; Stampar, F.; Veberic, R. The influence of organic/integrated production on the content of phenolic compounds in apple leaves and fruits in four different varieties over a 2-year period. J. Sci. Food Agric. 2010, 90, 2366–2378. [Google Scholar] [CrossRef] [PubMed]
- Viškelis, J.; Liaudanskas, M.; Uselis, N.; Kviklys, D. Internal fruit quality depends on the regulation of apple-tree vegetative growth. In Proceedings of the XXX International Horticultural Congress IHC2018: International Symposium on Cultivars, Rootstocks and Management Systems of 1281, Istanbul, Turkey, 12–16 August 2018; pp. 643–648. [Google Scholar] [CrossRef]
- Hua, Y.; Guo, H.; Zhou, X.G.; Li, X.; Yang, S.; Song, Y.; Ma, N.; Chai, C.; Qiao, X.; Li, L. Correlations between soluble sugar and phenol contents in leaves and pear scab resistance. J. Plant Pathol. Microbiol. 2014, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Sharma, M.; Ali, R.F.; Yousuf, M.; Hussain, A. In vitro antioxidant activity and spectrophotometric quantification of total phenolic and flavonoid contents of Malus domestica. Trop J. Pharm. Res. 2013, 3, 452–471. [Google Scholar]
- Magel, E.; Einig, W.; Hampp, R. Carbohydrates in trees. Dev. Crop. Sci. 2000, 26, 317–336. [Google Scholar] [CrossRef]
- Kviklys, D.; Samuolienė, G. Relationships among the rootstock, crop load, and sugar hormone signaling of apple tree, and their effects on biennial bearing. Front. Plant Sci. 2020, 11, 1213. [Google Scholar] [CrossRef]
- Samuolienė, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between apple tree rootstock, crop-load, plant nutritional status and yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Loescher, W.H.; Mccamant, T.; Keller, J.D. Carbohydrate reserves, translocation, and storage in woody plant roots. Hortscience 1990, 25, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Zhou, R.; Reider, E.J.; Sharkey, T.D.; Dandekar, A.M. Antisense inhibition of sorbitol synthesis leads to up-regulation of starch synthesis without altering CO2 assimilation in apple leaves. Planta 2005, 220, 767–776. [Google Scholar] [CrossRef]
- Li, M.; Li, P.; Ma, F.; Dandekar, A.M.; Cheng, L. Sugar metabolism and accumulation in the fruit of transgenic apple trees with decreased sorbitol synthesis. Hort. Res. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nii, N. Changes of Starch and Sorbitol in Leaves Before and After Removal of Fruits from Peach Trees. Ann. Bot. 1997, 79, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Kelc, D.; Lakota, M.; Vindis, P. Mesurments of sugars in leaves on apple trees. In DAAAM International Scientific Book; Katalinic, B., Ed.; DAAAM International: Vienna, Austria, 2016; pp. 207–216. [Google Scholar] [CrossRef]
- Wibbe, M.L.; Blanke, M.M. Effects of defruiting on source–sink relationship, carbon budget, leaf carbohydrate content and water use efficiency of apple trees. Plant Physiol. 1995, 94, 529–533. [Google Scholar] [CrossRef]
- Hudina, M.; Colaric, M.; Stampar, F. Primary metabolites in the leaves and fruits of three pear cultivars during the growing season. Can. J. Plant Sci. 2006, 87, 327–332. [Google Scholar] [CrossRef]
- Wang, Z.; Stutte, G.W. The role of carbohydrates in active osmotic adjustment in apple under water stress. J. Am. Soc. Hortic. Sci. 1992, 117, 816–823. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, G.K.; Fioravanço, J.C. Yield of ‘Royal Gala’ apple trees in response to pruning before or after leaf drop. Pesqui. Agropecu. Bras. 2018, 53, 427–434. [Google Scholar] [CrossRef]
- Greer, D.H.; Wünsche, J.N.; Halligan, E.A. Influence of postharvest temperatures on leaf gas exchange, carbohydrate reserves and allocations, subsequent budbreak, and fruit yield of ‘Braeburn’ apple (Malus domestica) trees. N. Z. J. Crop Hortic. Sci. 2002, 30, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H. Limitations to photosynthesis of leaves of apple (Malus domestica) trees across the growing season prior to and after harvest. Photosynthetica 2019, 57, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Fanwoua, J.; Bairam, E.; Delaire, M.; Buck-Sorlin, G. The role of branch architecture in assimilate production and partitioning: The example of apple (Malus domestica). Front. Plant Sci. 2014, 5, 338. [Google Scholar] [CrossRef] [Green Version]
- Melke, A. The physiology of chilling temperature requirements for dormancy release and bud-break in temperate fruit trees grown at mild winter tropical climate. J. Plant Res. 2015, 4, 110–156. [Google Scholar] [CrossRef]
- Yamaki, S. Isolation of vacuoles from immature apple fruit flesh and compartmentation of sugars, organic acids, phenolic compounds and amino acids. Plant Cell Physiol. 1984, 25, 151–166. [Google Scholar] [CrossRef]
- Colaric, M.; Veberic, R.; Stampar, F.; Hudina, M. Evaluation of peach and nectarine fruit quality and correlations between sensory and chemical attributes. J. Sci. Food Agric. 2005, 85, 2611–2616. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, E.; Boo, K.H.; Kim, H.Y.; Eigenheer, R.A.; Phinney, B.S.; Shulaev, V.; Negre-Zakharov, F.; Sadka, A.W. Label-free shotgun proteomics and metabolite analysis reveal a significant metabolic shift during citrus fruit development. J. Exp. Bot. 2011, 62, 5367–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardozza, S.; Boldingh, H.L.; Osorio, S.; Höhne, M.; Wohlers, M.; Gleave, A.P.; MacRae, E.A.; Richardson, A.C.; Atkinson, R.G.; Sulpice, R.; et al. Metabolic analysis of kiwifruit (Actinidia deliciosa) berries from extreme genotypes reveals hallmarks for fruit starch metabolism. J. Exp. Bot. 2013, 64, 5049–5063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desnoues, E.; Gibon, Y.; Baldazzi, V.; Signoret, V.; Génard, M.; Quilot-Turion, B. Profiling sugar metabolism during fruit development in a peach progeny with different fructose-to-glucose ratios. BMC Plant Biol. 2014, 14, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Li, D.; Feng, F.; Zhang, S.; Ma, F.; Cheng, L. Proteomic analysis reveals dynamic regulation of fruit development and sugar and acid accumulation in apple. J. Exp. Bot. 2016, 67, 5145–5157. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; He, M.; Bai, Y.; Xu, H.; Dandekar, A.M.; Fei, Z.; Cheng, L. Decreased sorbitol synthesis leads to abnormal stamen development and reduced pollen tube growth via a MYB transcription factor, MdMYB39L, in apple (Malus domestica). New Phytol. 2018, 217, 641–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.; Li, C.; Park, H.J.; González, J.; Wang, J.; Dandekar, A.M.; Turgeon, B.G.; Cheng, L. Sorbitol modulates resistance to Alternaria alternata by regulating the expression of an NLR resistance gene in apple. Plant Cell 2018, 30, 1562–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirgedaitė-Šėžienė, V.; Laužikė, K.; Uselis, N.; Samuolienė, G. Metabolic Response of Malus domestica Borkh cv. Rubin Apple to Canopy Training Treatments in Intensive Orchards. Horticulturae 2022, 8, 300. https://doi.org/10.3390/horticulturae8040300
Sirgedaitė-Šėžienė V, Laužikė K, Uselis N, Samuolienė G. Metabolic Response of Malus domestica Borkh cv. Rubin Apple to Canopy Training Treatments in Intensive Orchards. Horticulturae. 2022; 8(4):300. https://doi.org/10.3390/horticulturae8040300
Chicago/Turabian StyleSirgedaitė-Šėžienė, Vaida, Kristina Laužikė, Nobertas Uselis, and Giedrė Samuolienė. 2022. "Metabolic Response of Malus domestica Borkh cv. Rubin Apple to Canopy Training Treatments in Intensive Orchards" Horticulturae 8, no. 4: 300. https://doi.org/10.3390/horticulturae8040300
APA StyleSirgedaitė-Šėžienė, V., Laužikė, K., Uselis, N., & Samuolienė, G. (2022). Metabolic Response of Malus domestica Borkh cv. Rubin Apple to Canopy Training Treatments in Intensive Orchards. Horticulturae, 8(4), 300. https://doi.org/10.3390/horticulturae8040300