
Woody Canker and Shoot Blight Caused by Botryosphaeriaceae and Diaporthaceae on Mango and Litchi in Italy

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Survey and Isolation
2.2. Morphological Characterization
2.3. Molecular Characterization
2.4. Phylogenetic Analysis
2.5. Pathogenicity Tests
3. Results
3.1. Field Survey and Morphological Characterization
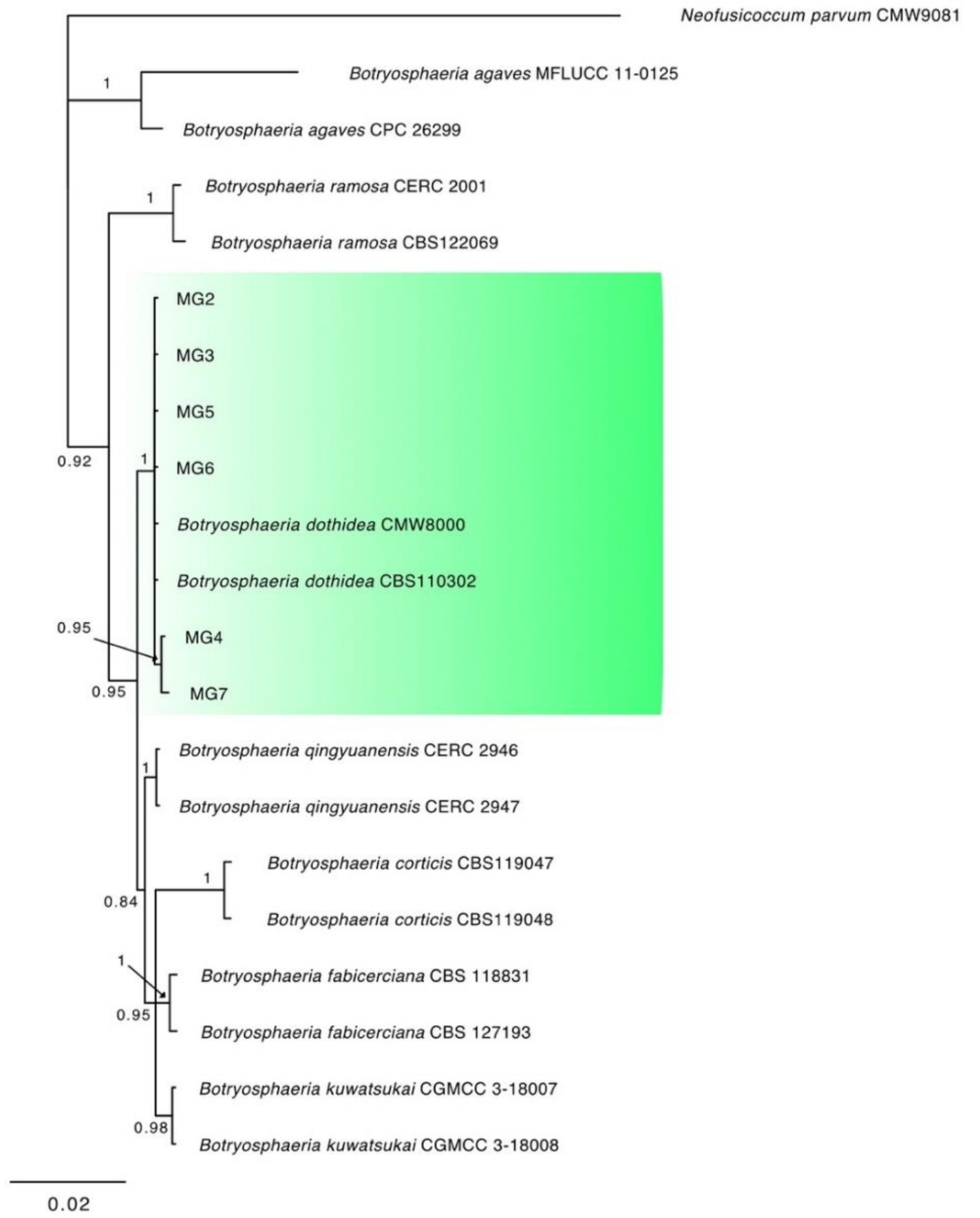
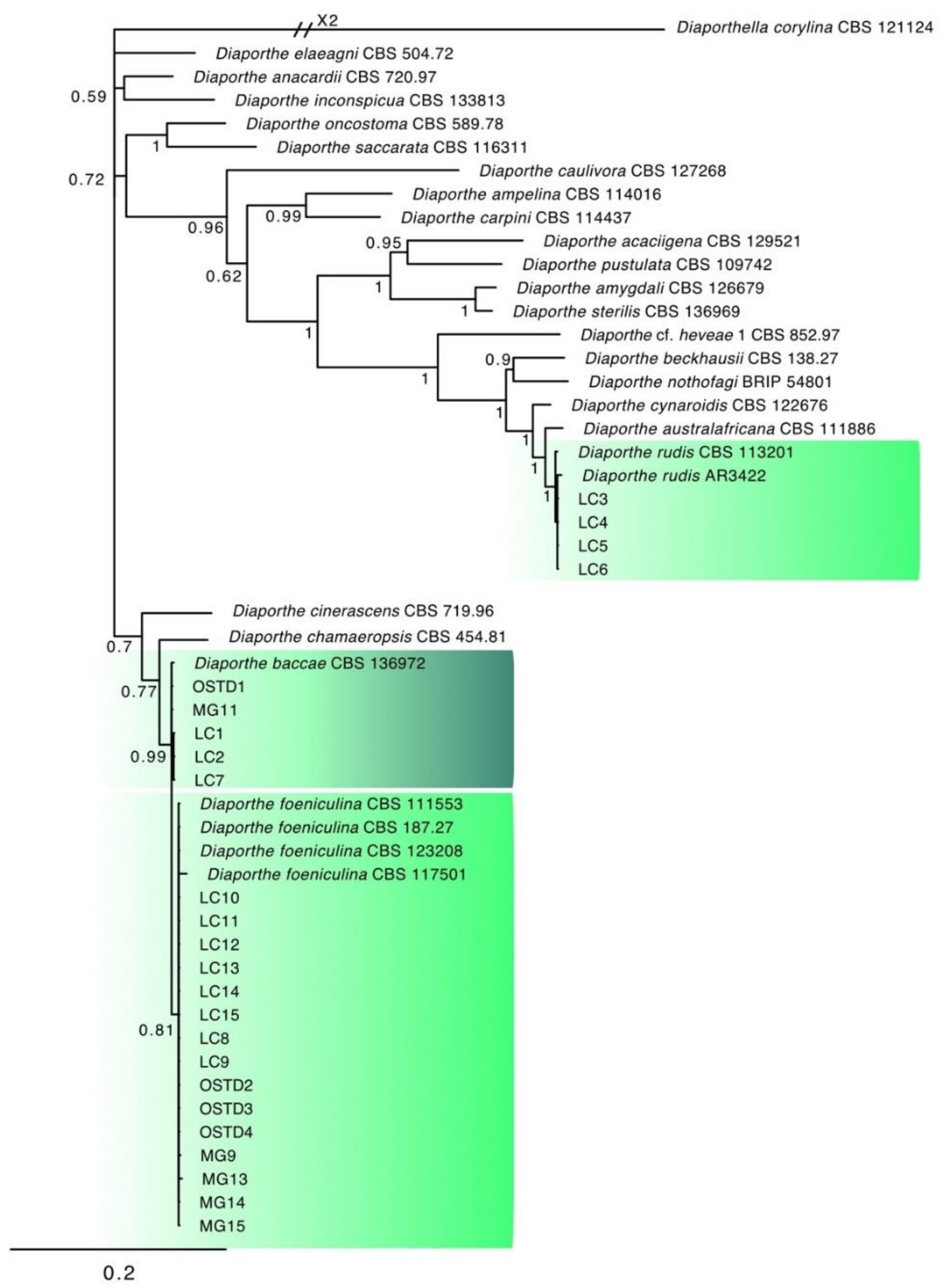
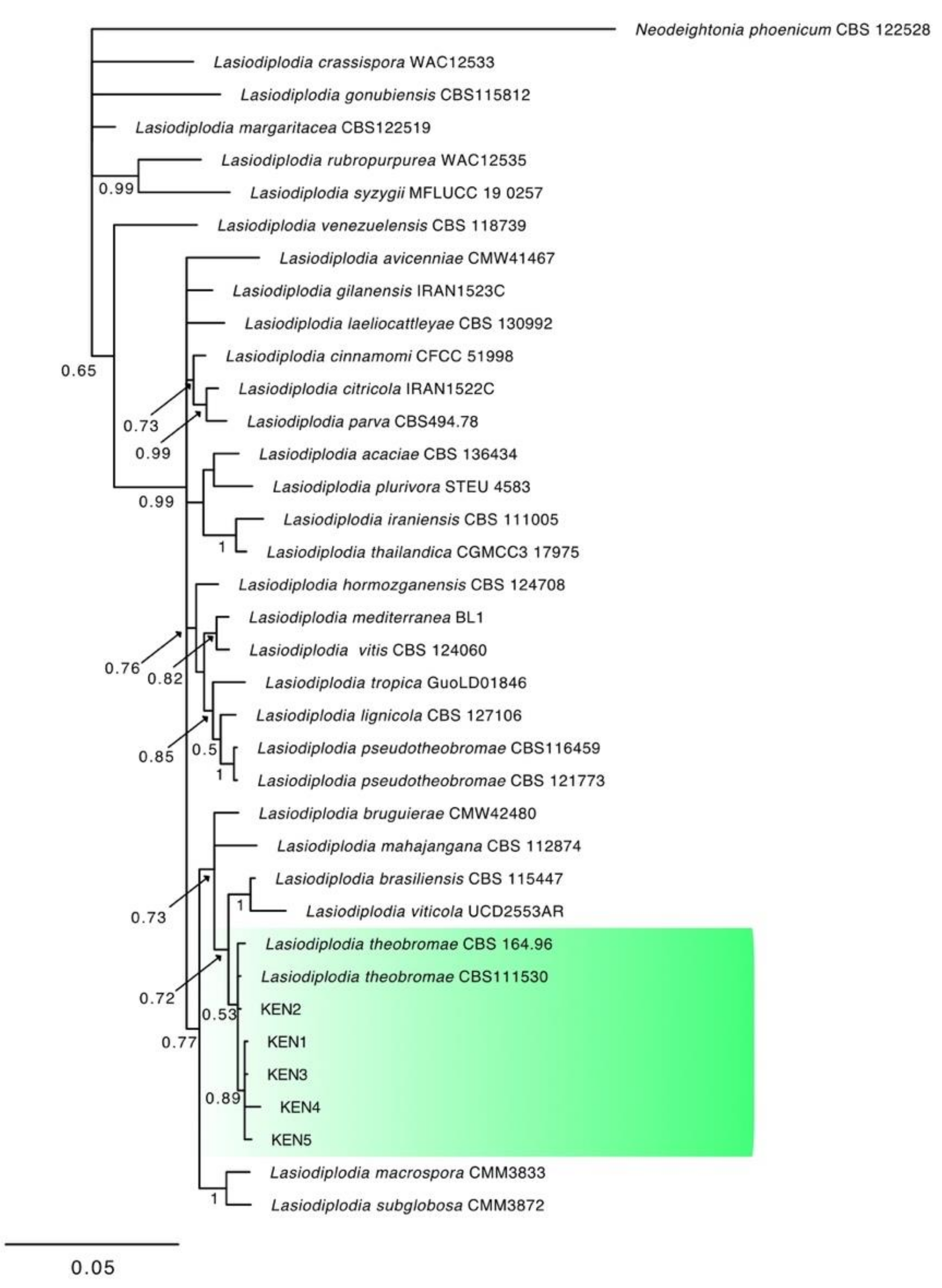
3.2. Molecular Characterization and Phylogenetic Analysis
3.3. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maldonado-Celis, M.E.; Yahia, E.M.; Bedoya, R.; Landázuri, P.; Loango, N.; Aguillón, J.; Restrepo, B.; Guerrero Ospina, J.C. Chemical Composition of Mango (Mangifera indica L.) Fruit: Nutritional and Phytochemical Compounds. Front. Plant Sci. 2019, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Ara, R.; Motalab, M.; Uddin, M.N.; Fakhruddin, A.N.M.; Saha, B.K. Nutritional evaluation of different mango varieties available in Bangladesh. Int. Food Res. J. 2014, 21, 2169–2174. [Google Scholar]
- Zhao, L.; Wang, K.; Wang, K.; Zhu, J.; Hu, Z. Nutrient components, health benefits, and safety of litchi (Litchi chinensis Sonn.): A review. Compr. Rev. Food. Sci. F. 2020, 19, 2139–2163. [Google Scholar] [CrossRef]
- Marçal, S.; Pintado, M. Mango peels as food ingredient/additive: Nutritional value, processing, safety and applications. Trends Food Sci. Technol. 2021, 114, 472–489. [Google Scholar] [CrossRef]
- FAO. Major Tropical Fruits—Statistical Compendium 2017; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; p. 38. [Google Scholar]
- Farina, V.; D’Asaro, A.; Mazzaglia, A.; Gianguzzi, G.; Palazzolo, E. Chemical-physical and nutritional characteristics of mature-green and mature-ripe ‘Kensington Pride’ mango fruit cultivated in Mediterranean area during cold storage. Fruits 2017, 72, 221–229. [Google Scholar] [CrossRef]
- Ismail, A.M.; Cirvilleri, G.; Lombard, L.; Crous, P.W.; Groenewald, J.Z.; Polizzi, G. Characterisation of Neofusicoccum species causing mango dieback in Italy. J. Plant Pathol. 2013, 95, 549–557. [Google Scholar]
- Ismail, A.M.; Cirvilleri, G.; Yaseen, T.; Epifani, F.; Perrone, G.; Polizzi, G. Characterisation of Colletotrichum species causing anthracnose disease of mango in Italy. J. Plant Pathol. 2015, 97, 167–171. [Google Scholar]
- Ismail, A.M.; Cirvilleri, G.; Polizzi, G. Characterisation and pathogenicity of Pestalotiopsis uvicola and Pestalotiopsis clavispora causing grey leaf spot of mango (Mangifera indica L.) in Italy. Eur. J. Plant Pathol. 2013, 135, 619–625. [Google Scholar] [CrossRef]
- Ahmed, Y.; Cirvilleri, G.; D’Onghia, A.M.; Yaseen, T. First report of Verticillium wilt of Mango (Mangifera indica) caused by Verticillium dahliae in Italy. Plant Dis. 2014, 98, 1156. [Google Scholar] [CrossRef]
- Trantas, E.A.; Mpalantinaki, E.; Pagoulatou, M.; Markakis, E.; Sarris, P.F.; Ververidis, F.; Goumas, D.E. First report of bacterial apical necrosis of mango caused by Pseudomonas syringae pv. syringae in Greece. Plant Dis. 2017, 101, 1541. [Google Scholar] [CrossRef]
- Torta, L.; Lo Piccolo, S.; Burruano, S.; Lo Cantore, P.; Iacobellis, N.S. Necrosi apicale del mango (Mangifera indica L.) causata da Pseudomonas syringae pv. syringae van Hall in Sicilia. Info. Fitopat. 2003, 11, 44–46. [Google Scholar]
- Aiello, A.; Ferrante, P.; Vitale, A.; Polizzi, G.; Scortichini, M.; Cirvilleri, G. Characterisation of Pseudomonas syringae pv. syringae isolated from mango in Sicily and occurrence of copper-resistant strains. J. Plant Pathol. 2015, 97, 273–282. [Google Scholar]
- Cazorla, F.M.; Tores, J.A.; Olalla, L.; Perez-Garcıa, A.; Farre, J.M.; de Vicente, A. Bacterial apical necrosis of mango in southern Spain: A disease caused by Pseudomonas syringae pv. syringae. Phytopathology 1998, 88, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Subhadrabandhu, S.; Mitra, S.K.; Ben-Arie, R.; Stern, R.A. Origin, History, Production and Processing. In Litchi and Longan. Botany, Production and Uses; Menzel, C.M., Waite, G.K., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 1–23. [Google Scholar]
- Ye, W.; Wang, Y.; Shen, D.; Li, D.; Pu, T.; Jiang, Z.; Zhang, Z.; Zheng, X.; Tyler, B.M.; Wang, Y. Sequencing of the litchi downy blight pathogen reveals it is a Phytophthora species with downy mildew-like characteristics. Mol. Plant-Microbe Interact. 2016, 29, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, J.F.; Song, X.B.; Xi, P.G.; Cheng, B.P.; Cui, Y.P.; Chen, X.; Peng, A.T.; Jiang, Z.D.; Zhang, L.H. Identification of Colletotrichum siamense causing litchi pepper spot disease in mainland China. Plant Pathol. 2019, 68, 1533–1542. [Google Scholar] [CrossRef]
- Anderson, J.M.; Aitken, E.A.B.; Dann, E.K.; Coates, L.M. Morphological and molecular diversity of Colletotrichum spp. causing pepper spot and anthracnose of lychee (Litchi chinensis) in Australia. Plant Pathol. 2013, 62, 279–288. [Google Scholar] [CrossRef]
- Kumar, V.; Anal, A.K.D.; Rai, S.; Nath, V. Leaf, panicle and fruit blight of litchi (Litchi chinensis) caused by Alternaria alternata in Bihar state, India. Can. J. Plant Pathol. 2018, 40, 84–89. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin–Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F.; et al. Genera of phytopathogenic fungi: GOPHY2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v. 2. Program Distributed by the Author; Uppsala Evolutionary Biology Centre Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Phillips, A.J.L. Resolving the complex of Diaporthe (Phomopsis) species occurring on Foeniculum vulgare in Portugal. Fungal Divers. 2009, 34, 111–125. [Google Scholar]
- Turco, E.; Marianelli, L.; Vizzuso, C.; Ragazzi, A.; Gini, R.; Selleri, B.; Tucci, R. First report of Botryosphaeria dothidea on sycamore, red oak, and english oak in North western Italy. Plant Dis. 2006, 90, 1106. [Google Scholar] [CrossRef]
- Carlucci, A.; Lops, F.; Raimondo, M.L.; Gentile, V.; Mucci, M.; Frisullo, S. The Botryosphaeria species from vineyards of Apulia. Phytopathol. Mediterr. 2009, 48, 180. [Google Scholar]
- Carlucci, A.; Cibelli, F.; Lops, F.; Raimondo, M.L. Characterisation of Botryosphaeriaceae species as causal agents of trunk diseases on grapevines. Plant Dis. 2015, 99, 1678–1688. [Google Scholar] [CrossRef] [Green Version]
- Deidda, A.; Buffa, F.; Linaldeddu, B.T.; Pinna, C.; Scanu, B.; Deiana, V.; Satta, A.; Franceschini, A.; Floris, I. Emerging pests and diseases threaten Eucalyptus camaldulensis plantations in Sardinia, Italy. Forest 2016, 9, 883–891. [Google Scholar] [CrossRef]
- Gusella, G.; Lawrence, D.P.; Aiello, D.; Luo, Y.; Polizzi, G.; Michailides, T. Etiology of Botryosphaeria Panicle and Shoot Blight of Pistachio (Pistacia vera) caused by Botryosphaeriaceae in Italy. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fiorenza, A.; Aiello, D.; Costanzo, M.B.; Gusella, G.; Polizzi, G. A New Disease for Europe of Ficus microcarpa Caused by Botryosphaeriaceae Species. Plants 2022, 11, 727. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Van Leeuwen, G.C.M.; Guarnaccia, V.; Polizzi, G.; Van Rijswick, P.C.; Rosendahl, K.C.H.M.; Gabler, J.; Crous, P.W. Diaporthe species associated with Vaccinium, with specific reference to Europe. Phytopathol. Mediterr. 2014, 53, 287–299. [Google Scholar]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarnaccia, V.; Martino, I.; Tabone, G.; Brondino, L.; Gullino, M.L. Fungal pathogens associated with stem blight and dieback of blueberry in northern Italy. Phytopathol. Mediterr. 2020, 59, 229–245. [Google Scholar]
- Guarnaccia, V.; Vitale, A.; Cirvilleri, G.; Aiello, D.; Susca, A.; Epifani, F.; Perrone, G.; Polizzi, G. Characterisation and pathogenicity of fungal species associated with branch cankers and stem-end rot of avocado in Italy. Eur. J. Plant Pathol. 2016, 146, 963–976. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Crous, P.W.; Aiello, D.; Gullino, M.L.; Polizzi, G.; Guarnaccia, V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants 2021, 10, 492. [Google Scholar] [CrossRef]
- Burruano, S.; Mondello, V.; Conigliaro, G.; Alfonzo, A.; Spagnolo, A.; Mugnai, L. Grapevine decline in Italy caused by Lasiodiplodia theobromae. Phytopathol. Mediterr. 2008, 47, 132–136. [Google Scholar]
- Garibaldi, A.; Bertetti, D.; Amatulli, M.T.; Cardinale, J.; Gullino, M.L. First report of postharvest fruit rot in avocado (Persea americana) caused by Lasiodiplodia theobromae in Italy. Plant Dis. 2012, 96, 460. [Google Scholar] [CrossRef]
- Aiello, D.; Gusella, G.; Fiorenza, A.; Guarnaccia, V.; Polizzi, G. Identification of Neofusicoccum parvum causing Canker and Twig Blight on Ficus carica in Italy. Phytopathol. Mediterr. 2020, 59, 213–218. [Google Scholar] [CrossRef]
- Lorenzini, M.; Zapparoli, G. Diaporthe rudis associated with berry rot of postharvest grapes in Italy. Plant Dis. 2019, 103, 1030. [Google Scholar] [CrossRef]
- Ploetz, R.C. The major diseases of mango: Strategies and potential for sustainable management. Acta Hortic. 2004, 645, 137–150. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Benscher, D.; Vázquez, A.; Colls, A.; Nagel, J.; Schaffer, B. A re-examination of mango decline in Florida. Plant Dis. 1996, 80, 664–668. [Google Scholar] [CrossRef]
- Slippers, B.; Johnson, G.I.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Phylogenetic and Morphological Re-Evaluation of the Botryosphaeria Species Causing Diseases of Mangifera indica. Mycologia 2005, 97, 99–110. [Google Scholar] [CrossRef]
- Johnson, G.I. Status of mango postharvest disease management R&D: Options and solutions for the Australian mango industry. Hortic. Aust. 2008, 4, 1–130. [Google Scholar]
- de Oliveira Costa, V.S.; Michereff, S.J.; Martins, R.B.; Gava, C.A.T.; Mizubuti, E.S.G.; Camara, M.P.S. Species of Botryosphaeriaceae associated on mango in Brazil. Eur. J. Plant Pathol. 2010, 127, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E.S.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Wingield, M.J.; Crous, P.W.; Coutinho, T.A. Sphaeropsis sapinea and Botryosphaeria dothidea endophytic in Pinus spp. and Eucalyptus spp. in South Africa. S. Afr. J. Bot. 1996, 62, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Wingfield, M.J.; Petrini, O. Botryosphaeria dothidea endophytic in Eucalyptus grandis and Eucalyptus nitens in South Africa. For. Ecol. Manag. 1996, 89, 189–195. [Google Scholar] [CrossRef]
- Stanosz, G.R.; Smith, D.R.; Albers, A.J. Surveys for asymptomatic persistence of Sphaeropsis sapinea on or in stems of red pine seedlings from seven Great Lakes region nurseries. For. Pathol. 2005, 35, 233–244. [Google Scholar] [CrossRef]
- Lavalle, C.; Micale, F.; Houston, T.D.; Camia, A.; Hiederer, R.; Lazar, C.; Conte, C.; Amatulli, G.; Genovese, G. Climate change in Europe. 3. Impact on agriculture and forestry. A review. Agron. Sustain. Dev. 2009, 29, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Ma, Z.; Morgan, D.P.; Michailides, T.J. Effects of water stress on Botryosphaeria blight of pistachio caused by Botryosphaeria dothidea. Plant Dis. 2001, 85, 745–749. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.A.; Hendrix, F.F. Pathogenicity and histopathology of Botryosphaeria dothidea on apple stems. Phytopathology 1981, 71, 375–379. [Google Scholar] [CrossRef]
- Blodgett, J.T.; Kruger, E.L.; Stanosz, G.R. Effects of moderate water stress on disease development by Sphaeropsis sapinea on red pine. Phytopathology 1997, 87, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Blodgett, J.T.; Kruger, E.L.; Stanosz, G.R. Sphaeropsis sapinea and water stress in a red pine plantation in central Wisconsin. Phytopathology 1997, 87, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, E.; Danti, R.; Strati, S. Pre- and post-inoculation water stress affects Sphaeropsis sapinea canker length in Pinus halepensis. For. Pathol. 2001, 31, 209–218. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; Mckenzie, E.; Chukeatirote, E.; Bahkali, A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Lim, L.; Mohd, M.H.; Zakaria, L. Identification and pathogenicity of Diaporthe species associated with stem-end rot of mango (Mangifera indica L.). Eur. J. Plant Pathol. 2019, 155, 687–696. [Google Scholar] [CrossRef]
- Serrato-Diaz, L.M.; Rivera-Vargas, L.I.; French-Monar, R.D. First report of Diaporthe pseudomangiferae causing inflorescence rot, rachis canker and flower abortion on mango. Plant Dis. 2014, 98, 1004. [Google Scholar] [CrossRef] [PubMed]
- Ajitomi, A.; Minoshima, A.; Takushi, T.; Truong, H.H.; Ooshiro, A.; Yamashiro, M.; Arasaki, C.; Hirooka, Y. First report of mango (Mangifera indica) stem-end rot caused by two Diaporthe species and their susceptibility to procymidone. J. Gen. Plant Pathol. 2020, 86, 237–244. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandecic, K.; Cosic, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcmillan, J.R.R.T. Diseases of Litchi chinensis in south Florida. Proc. Fla. State Hortic. Soc. 1994, 107, 360–361. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Culture No. | Host/Cultivar | Genbank Accession Number | ||
|---|---|---|---|---|---|
| ITS | tef1 | tub2 | |||
| Lasiodiplodia theobromae | KEN1 | Mangifera indica cv. Kent | OL470874 | OL891701 | OL891738 |
| L. theobromae | KEN2 | M. indica cv. Kent | OL470875 | OL891699 | OL891742 |
| L. theobromae | KEN3 | M. indica cv. Kent | OL470876 | OL891700 | OL891739 |
| L. theobromae | KEN4 | M. indica cv. Kent | OL470877 | OL891708 | OL891741 |
| L. theobromae | KEN5 | M. indica cv. Kent | OL470878 | OL891709 | OL891740 |
| Neofusicoccum parvum | KEI1 | M. indica cv. Keitt | OL470866 | OL891684 | OL891757 |
| N. parvum | KEI2 | M. indica cv. Keitt | OL470867 | OL891690 | OL891756 |
| N. parvum | KEI3 | M. indica cv. Keitt | OL470868 | OL891695 | OL955497 |
| N. parvum | KEI4 | M. indica cv. Keitt | OL470869 | OL891696 | OL891751 |
| N. parvum | KEI5 | M. indica cv. Keitt | OL470870 | OL891697 | OL891752 |
| N. parvum | KEI6 | M. indica cv. Keitt | OL470871 | OL891698 | OL891753 |
| N. parvum | KEI7 | M. indica cv. Keitt | OL470872 | OL891710 | OL891754 |
| N. parvum | KEI8 | M. indica cv. Keitt | OL470873 | OL891711 | OL891759 |
| N. parvum | SENS1 | M. indica cv. Sensation | OL470882 | OL891688 | OL955498 |
| N. parvum | SENS2 | M. indica cv. Sensation | OL470883 | OL891689 | OL955500 |
| N. parvum | SENS3 | M. indica cv. Sensation | OL470884 | OL891713 | OL891750 |
| N. parvum | SENS4 | M. indica cv. Sensation | OL470885 | OL891691 | OL955496 |
| N. parvum | SENS5 | M. indica cv. Sensation | OL470886 | OL891692 | OL891758 |
| N. parvum | SENS6 | M. indica cv. Sensation | OL470887 | OL891712 | OL955495 |
| N. parvum | SENS7 | M. indica cv. Sensation | OL470888 | OL891693 | OL955499 |
| N. parvum | SENS8 | M. indica cv. Sensation | OL470889 | OL891694 | OL955494 |
| N. parvum | OSTB1 | M. indica cv. Osteen | OL470879 | OL891685 | OL891749 |
| N. parvum | OSTB2 | M. indica cv. Osteen | OL470880 | OL891686 | OL891760 |
| N. parvum | OSTB3 | M. indica cv. Osteen | OL470881 | OL891687 | OL891755 |
| Diaporthe foeniculina | OSTD1 | M. indica cv. Osteen | OL477409 | OL891674 | OL891729 |
| D. foeniculina | OSTD2 | M. indica cv. Osteen | OL477410 | OL891675 | OL891730 |
| D. foeniculina | OSTD3 | M. indica cv. Osteen | OL477411 | OL891682 | OL891731 |
| D. foeniculina | OSTD4 | M. indica cv. Osteen | OL477412 | OL891676 | OL891720 |
| Botryosphaeria dothidea | MG2 | M. indica cv. Kensington Pride | OL470890 | OL891704 | OL891746 |
| B. dothidea | MG3 | M. indica cv. Kensington Pride | OL470891 | OL891706 | OL891743 |
| B. dothidea | MG4 | M. indica cv. Kensington Pride | OL470892 | OL891702 | OL891748 |
| B. dothidea | MG5 | M. indica cv. Kensington Pride | OL470893 | OL891705 | OL891744 |
| B. dothidea | MG6 | M. indica cv. Kensington Pride | OL470894 | OL891707 | OL891745 |
| B. dothidea | MG7 | M. indica cv. Kensington Pride | OL470895 | OL891703 | OL891747 |
| D. foeniculina | MG9 | M. indica cv. Kensington Pride | OL477413 | OL891678 | OL891722 |
| D. baccae | MG11 | M. indica cv. Kensington Pride | OL477414 | OL891681 | OL891733 |
| D. foeniculina | MG13 | M. indica cv. Kensington Pride | OL477415 | OL891677 | OL891727 |
| D. foeniculina | MG14 | M. indica cv. Kensington Pride | OL477416 | OL891679 | OL891728 |
| D. foeniculina | MG15 | M. indica cv. Kensington Pride | OL477417 | OL891683 | OL891721 |
| D. foeniculina | LC1 | Litchi chinensis cv. Way Chee | OL477395 | OL891664 | OL891714 |
| D. foeniculina | LC2 | L. chinensis cv. Way Chee | OL477402 | OL891670 | OL891723 |
| D. rudis | LC3 | L. chinensis cv. Way Chee | OL477403 | OL891660 | OL891737 |
| D. rudis | LC4 | L. chinensis cv. Way Chee | OL477404 | OL891662 | OL891734 |
| D. rudis | LC5 | L. chinensis cv. Way Chee | OL477405 | OL891663 | OL891736 |
| D. rudis | LC6 | L. chinensis cv. Way Chee | OL477406 | OL891661 | OL891735 |
| D. foeniculina | LC7 | L. chinensis cv. Way Chee | OL808652 | OL891671 | OL891724 |
| D. foeniculina | LC8 | L. chinensis cv. Way Chee | OL477407 | OL891672 | OL891725 |
| D. foeniculina | LC9 | L. chinensis cv. Kwai Mai Pink | OL477408 | OL891673 | OL891719 |
| D. foeniculina | LC10 | L. chinensis cv. Kwai Mai Pink | OL477396 | OL891665 | OL891732 |
| D. foeniculina | LC11 | L. chinensis cv. Kwai Mai Pink | OL477397 | OL891666 | OL891726 |
| D. foeniculina | LC12 | L. chinensis cv. Kwai Mai Pink | OL477398 | OL891667 | OL891715 |
| D. foeniculina | LC13 | L. chinensis cv. Kwai Mai Pink | OL477399 | OL891668 | OL891716 |
| D. foeniculina | LC14 | L. chinensis cv. Kwai Mai Pink | OL477400 | OL891680 | OL891717 |
| D. foeniculina | LC15 | L. chinensis cv. Kwai Mai Pink | OL477401 | OL891669 | OL891718 |
| Species | Morphological Characteristics | |||
|---|---|---|---|---|
| Colony on PDA | Pycnidia | Conidia | ||
| Diaporthe baccae | Cream colour becoming smoky gray, flattened, dense mycelium; grayish sepia on the lower surface | Brown to black, globose with yellowish masses on MEA | α conidia aseptate, hyaline, fusiform to ellipsoidal tapering toward both ends; 5.1–9.4 × 2–9.4 µm | β conidia aseptate, hyaline, apex acutely rounded, base truncate; 16.8–27.1 × 0.8–2.2 µm |
| D. foeniculina | White, sparse, cottony, aerial mycelium; greenish yellow on the lower surface | Black, globose to subglobose with yellowish conidial masses on PNA | α conidia aseptate, hyaline, ellipsoidal to fusiform, rarely with substruncate base; 3.9–11.1 × 1.5–3.4 µm | β conidia aseptate, hyaline, slightly curved with substruncate base; 15.8–27.1 × 0.8–2.2 µm |
| D. rudis | White, fluffy, aerial mycelium; whitish yellow on the lower surface | not observed | α conidia not observed | β conidia not observed |
| Botryosphaeria dothidea | Olivaceous becoming grayish-dark brown, dense mycelium; grayish-dark brown on the lower surface | Black to brown, globose to subglobose on PNA | Aseptate, hyaline, fusiform to subclavate with substruncate to blunty rounded base; 17.7–27.8 × 4.2–7.4 µm | - |
| Lasiodiplodia theobromae | White, dense aerial mycelium becoming grayish green; yellowish gray on the lower surface | Black, globose to subglobose on PNA | Aseptate, hyaline, subovoid to ellipsoid immature, become 1-septate with age, dark brown, thick walled, ellipsoidal; 22.1–26.6 × 12.9–14.5 µm | - |
| Neofusicoccum parvum | White, dense aerial mycelium becoming greenish gray; greenish gray on the lower surface | Dark to brown, globose to subglobose on PNA | Aseptate, old conidia becoming 1–2 septate, hyaline, ellipsoidal with apex round and base flat; 13.3–20.9 × 5.2–7.5 µm | - |
| Botryosphaeria | Diaporthe | Lasiodiplodia | Neofusicoccum | |
|---|---|---|---|---|
| Locus(i) | ITS + tef1 + tub2 | ITS + tef1 + tub2 | ITS + tef1 + tub2 | ITS + tef1 + tub2 |
| Unique site patterns of ITS | 92 | 318 | 56 | 44 |
| Unique site patterns of tef1 | 114 | 151 | 108 | 31 |
| Unique site patterns of tub2 | 75 | 241 | 37 | 39 |
| Generation ran | 845,000 | 1,790,000 | 880,000 | 3,195,000 |
| Generated trees | 1692 | 3582 | 1762 | 6392 |
| Sampled trees | 1270 | 2688 | 1322 | 4794 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, D.; Guarnaccia, V.; Costanzo, M.B.; Leonardi, G.R.; Epifani, F.; Perrone, G.; Polizzi, G. Woody Canker and Shoot Blight Caused by Botryosphaeriaceae and Diaporthaceae on Mango and Litchi in Italy. Horticulturae 2022, 8, 330. https://doi.org/10.3390/horticulturae8040330
Aiello D, Guarnaccia V, Costanzo MB, Leonardi GR, Epifani F, Perrone G, Polizzi G. Woody Canker and Shoot Blight Caused by Botryosphaeriaceae and Diaporthaceae on Mango and Litchi in Italy. Horticulturae. 2022; 8(4):330. https://doi.org/10.3390/horticulturae8040330
Chicago/Turabian StyleAiello, Dalia, Vladimiro Guarnaccia, Mariangela Benedetta Costanzo, Giuseppa Rosaria Leonardi, Filomena Epifani, Giancarlo Perrone, and Giancarlo Polizzi. 2022. "Woody Canker and Shoot Blight Caused by Botryosphaeriaceae and Diaporthaceae on Mango and Litchi in Italy" Horticulturae 8, no. 4: 330. https://doi.org/10.3390/horticulturae8040330
APA StyleAiello, D., Guarnaccia, V., Costanzo, M. B., Leonardi, G. R., Epifani, F., Perrone, G., & Polizzi, G. (2022). Woody Canker and Shoot Blight Caused by Botryosphaeriaceae and Diaporthaceae on Mango and Litchi in Italy. Horticulturae, 8(4), 330. https://doi.org/10.3390/horticulturae8040330