
Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Growing Conditions
2.2. Data Collection
2.3. Data Analysis
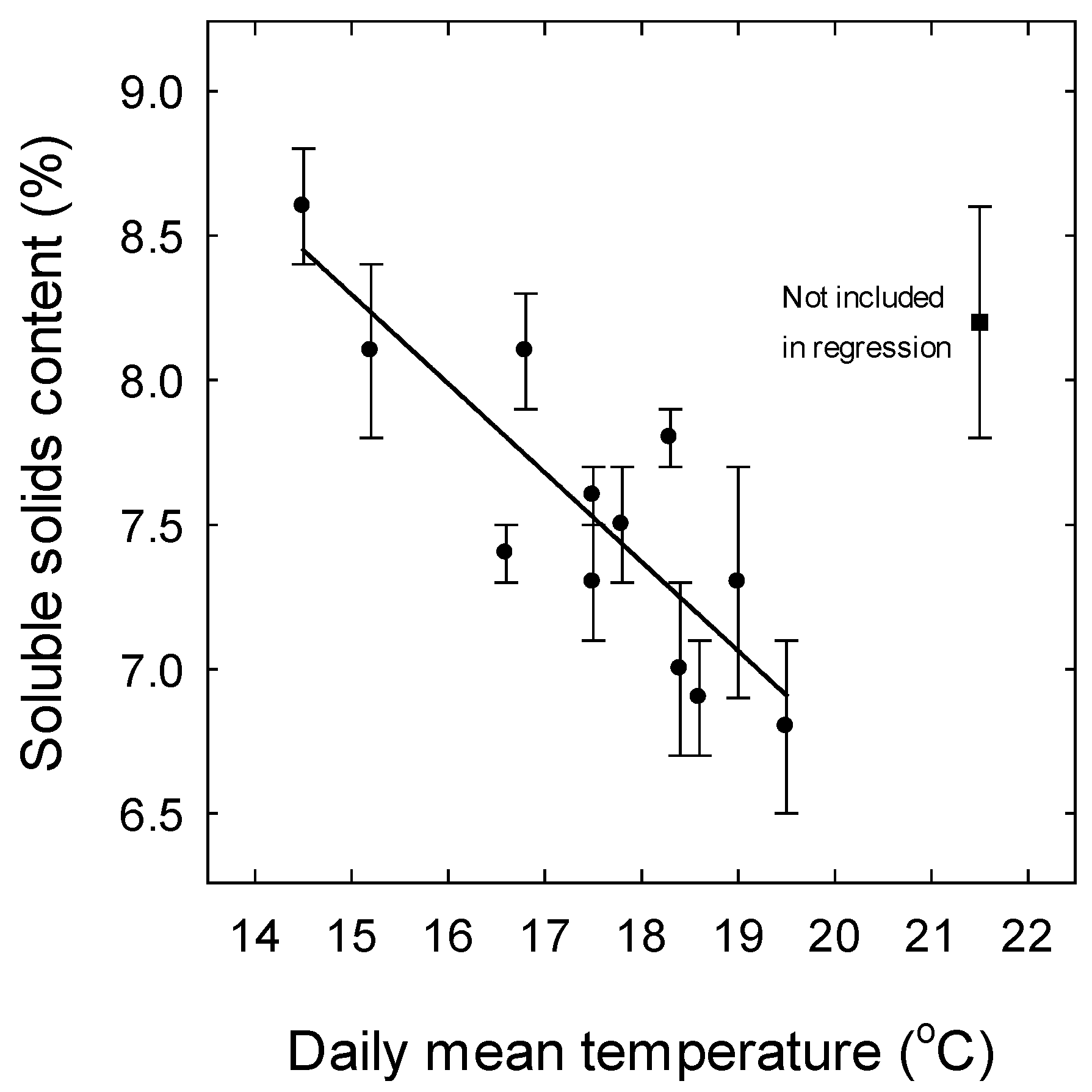
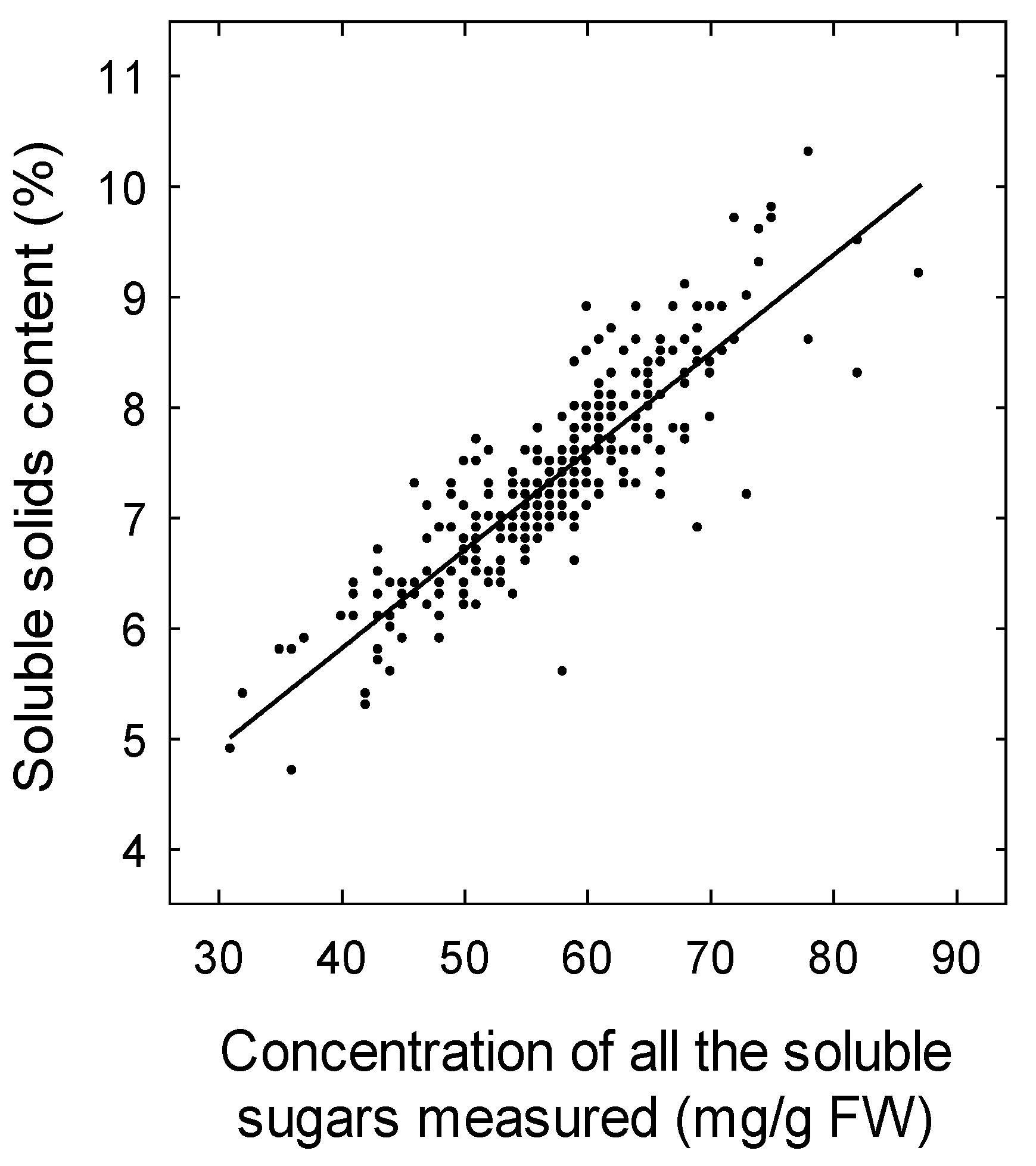
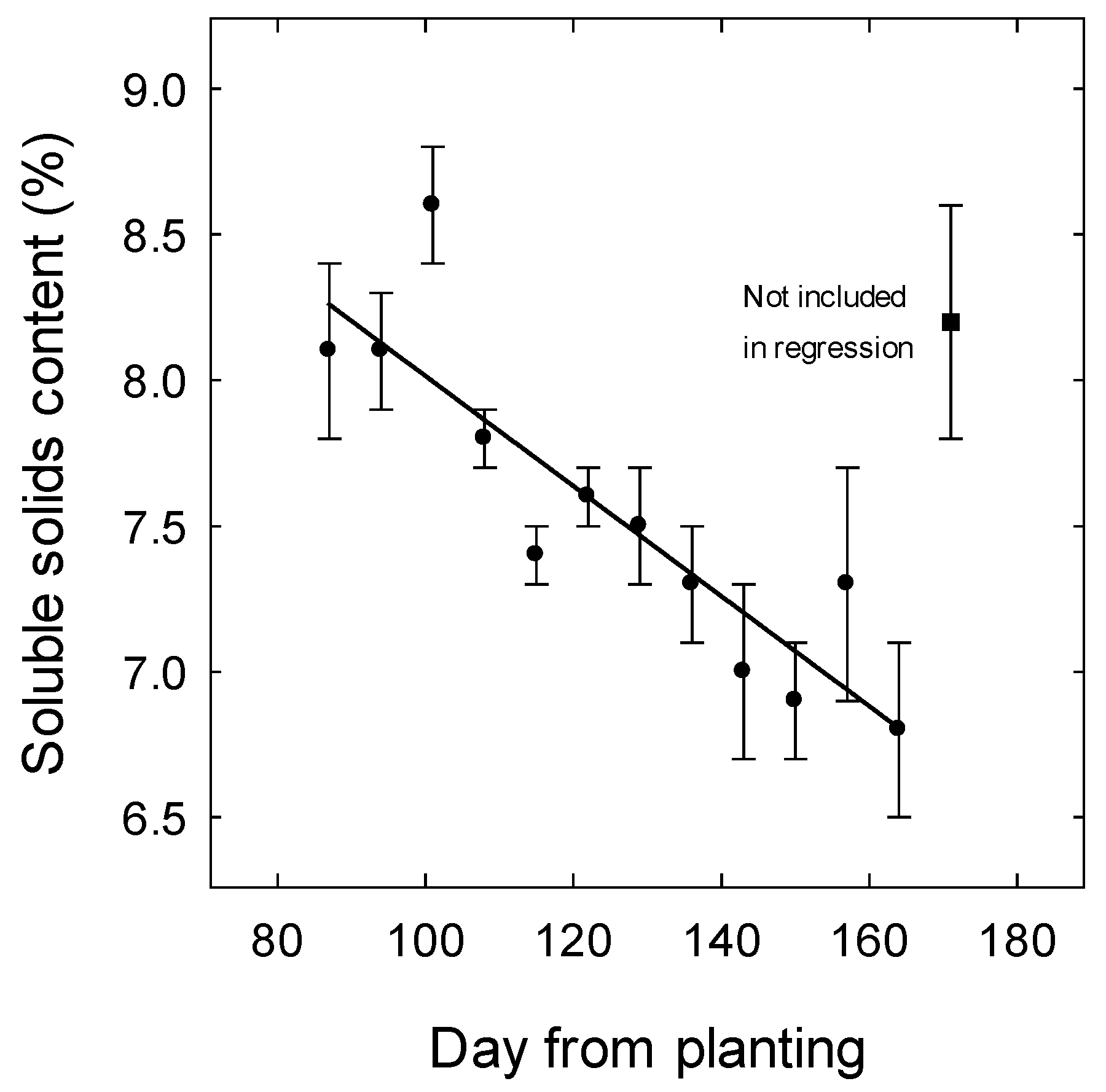
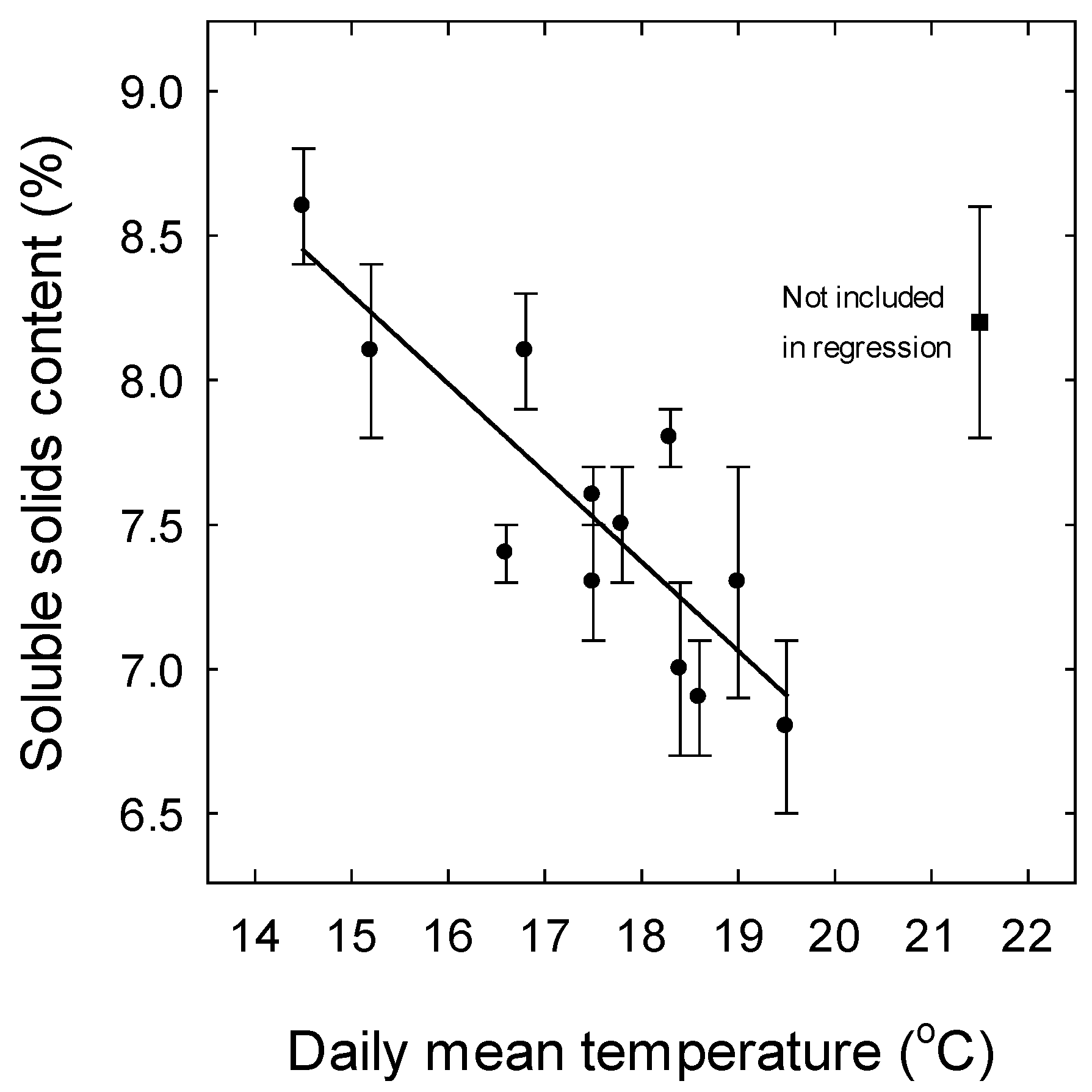
3. Results
3.1. Weather
3.2. Plant Growth
3.3. Yield and Fruit Quality
4. Discussion
4.1. Yields
4.2. Effect of Cultivar on Soluble Solids Content and Titratable Acidity
4.3. Effect of Temperature on Soluble Solids Content and Titratable Acidity
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lobell, D.; Field, C. California perennial crops in a changing climate. Clim. Chang. 2011, 109, 317–333. [Google Scholar] [CrossRef]
- Deschenes, O.; Kolstad, C. Economic impacts of climate change on California agriculture. Clim. Chang. 2011, 109, 365–386. [Google Scholar] [CrossRef] [Green Version]
- Lobell, D.; Cahill, K.N.; Field, C.B. Historical effects of temperature and precipitation on California crop yields. Change 2007, 81, 187–203. [Google Scholar] [CrossRef]
- Mackenzie, S.J.; Chandler, C.K.; Hasing, T.; Whitaker, V.M. The role of temperature in the late-season decline in soluble solids content of strawberry fruit in a subtropical production system. HortScience 2011, 46, 1562–1566. [Google Scholar] [CrossRef]
- Cui, M.; Pham, M.D.; Hwang, H.; Chun, C. Flower development and fruit malformation in strawberries after short-term exposure to high or low temperature. Sci. Hortic. 2021, 288, 110308. [Google Scholar] [CrossRef]
- Sone, K.; Mochizuki, T.; Noguchi, Y. Relationship between stability of some eating quality of strawberry cultivars and their sugar and organic acid contents. J. Jpn. Soc. Hortic. Sci. 2000, 69, 736–743. [Google Scholar] [CrossRef]
- Ikegaya, A.; Ohba, S.; Toyoizumi, T.; Arai, E. Quality evaluation of strawberries grown in various regions by Singaporeans and Japanese. Int. J. Fruit Sci. 2021, 21, 883–895. [Google Scholar] [CrossRef]
- Fan, Z.; Hasing, T.; Johnson, T.S.; Garner, D.M.; Schweiterman, M.L.; Barbey, C.R.; Colquhoun, T.A.; Sims, C.A.; Resende, M.F.R.; Whitaker, V.M. Strawberry sweetness and consumer preference are enhanced by specific volatile compounds. Hortic. Res. 2021, 8, 66. [Google Scholar] [CrossRef]
- Hagenguth, J.; Kanski, L.; Kahle, H.; Naumann, M.; Pawelzik, E.; Becker, H.C.; Horneburg, B. Breeders’ sensory test: A new tool for early selection in breeding for tomato (Solanum lycopersicum) flavour. Plant Breed. 2022, 141, 96–107. [Google Scholar] [CrossRef]
- Yan, J.-W.; Ban, Z.-J.; Lu, H.-Y.; Li, D.; Poverenov, E.; Luo, Z.-S.; Li, L. The aroma volatile repertoire in strawberry fruit: A review. J. Sci. Food Agric. 2018, 98, 4395–4402. [Google Scholar] [CrossRef]
- Akšić, M.F.; Tosti, T.; Sredojević, M.; Milivojević, J.; Meland, M.; Natić, M. Comparison of sugar profile between leaves and fruits of blueberry and strawberry cultivars grown in organic and integrated production system. Plants 2019, 8, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallio, H.; Hakala, M.; Pelkkikangas, A.; Lapveteläinen, A. Sugars and acids of strawberry varieties. Eur. Food Res. Technol. 2000, 212, 81–85. [Google Scholar] [CrossRef]
- Pistón, F.; Peréz, A.G.; Sanz, C.; Refoyo, A. Relationship between sugar content and brix degrees in strawberry. Acta Hortic. 2017, 1156, 491–496. [Google Scholar] [CrossRef]
- Osatuke, A.; Pritts, M. Strawberry flavor is influenced by the air temperature differential during fruit development but not management practices. Agronomy 2021, 11, 606. [Google Scholar] [CrossRef]
- Hoppula, K.B.; Karhu, S.T. Strawberry fruit quality response to the production environment. J. Food Agric. Environ. 2006, 4, 166–170. [Google Scholar]
- Kruger, E.; Josuttis, M.; Nestby, R.; Toldam-Andersen, T.B.; Carlen, C.; Mezzetti, B. Influence of growing conditions at different latitudes of Europe on strawberry growth performance, yield and quality. J. Berry Res. 2012, 2, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Fiorani, F.; Pieruschka, R.; Wojciechowski, T.; van der Putten, W.; Kleyer, M.; Schurr, U.; Postma, J. Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytol. 2016, 212, 838–855. [Google Scholar] [CrossRef]
- Forero, L.E.; Grenzer, J.; Heinze, J.; Schittko, C.; Kulmatiski, A. Greenhouse- and field-measured plant-soil feedbacks are not correlated. Front. Environ. Sci. 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Yan, L.; Xia, J. A threefold difference in plant growth response to nitrogen addition between the laboratory and field experiments. Ecosphere 2019, 10, e02572. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Quero, E.; López, M.G. Carbohydrate differences in strawberry crowns and fruit (Fragaria × ananassa) during plant development. J. Agric. Food Chem. 2002, 50, 3317–3321. [Google Scholar] [CrossRef]
- Damyeh, M.S.; Fernando, C.W.; Phan, A.D.T.; Hong, H.T.; Mereddy, R.; Netzel, M.E.; Chaliha, M.; Sultanbawa, Y. Post-harvest fungal occurrence on commercial strawberry cultivars grown in Australia: Impact of phytochemical composition. J. Food Meas. Charact. 2021, 15, 3811–3822. [Google Scholar] [CrossRef]
- Scott, G.; Williams, C.; Wallace, R.W.; Du, X. Exploring plant performance, fruit physiochemical characteristics, volatile profiles, and sensory properties of day-neutral and short-day strawberry cultivars grown in Texas. J. Agric. Food Chem. 2021, 69, 13299–13314. [Google Scholar] [CrossRef] [PubMed]
- Duval, J.R.; Chandler, C.K.; Golden, E. Planting date affects early season fruit yield of strawberry in a subtropical environment. J. Am. Pomolog. Soc. 2005, 59, 7–12. [Google Scholar]
- Torres-Quezada, E.A.; Zotarelli, L.; Whitaker, V.M.; Santos, B.M.; Hernandez-Ochoa, I. Initial crown diameter of strawberry bare-root transplants affects early and total fruit yield. HortTechnology 2015, 25, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Chandler, C.K.; Legard, D.E.; Dunigan, D.D.; Crocker, T.E.; Sims, C.A. ‘Strawberry Festival’ strawberry. HortScience 2000, 35, 1366–1367. [Google Scholar] [CrossRef] [Green Version]
- Chandler, C.K.; Santos, B.M.; Peres, N.A.; Jouquand, C.; Plotto, A.; Sims, C.A. ‘Florida Radiance’ strawberry. HortScience 2009, 44, 1769–1770. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, V.M.; Osorio, L.F.; Peres, N.A.; Fan, Z.; Herrington, M.; Nunes, M.C.N.; Plotto, A.; Sims, C.A. ‘Florida Beauty’ strawberry. HortScience 2017, 52, 1443–1447. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, V.M.; Peres, N.A.; Osorio, L.F.; Fan, Z.; Nunes, M.C.N.; Plotto, A.; Sims, C.A. ‘Florida Brilliance’ strawberry. HortScience 2019, 54, 2073–2077. [Google Scholar] [CrossRef] [Green Version]
- Ariza, M.T.; Miranda, L.; Martínez-Ferri, E.; Medina, J.J.; Gómez-Mora, J.A.; Cervantes, L.; Soria, C. Consistency of organoleptic and yield related traits of strawberry cultivars over time. J. Berry Res. 2020, 10, 623–636. [Google Scholar] [CrossRef]
- Ogiwara, I.; Habutsu, S.; Hakoda, N.; Shimura, I. Soluble sugar content in nine wild and forty-one cultivated strawberries. J. Jap. Soc. Hortic. Sci. 1998, 67, 406–412. [Google Scholar] [CrossRef]
- Shaw, D.V.; Bringhurst, R.S.; Voth, V. Genetic variation for quality traits in an advanced-cycle breeding population of strawberries. J. Am. Soc. Hortic. Sci. 1987, 112, 699–702. [Google Scholar]
- Shaw, D.V. Genotypic variation and genotypic correlations for sugars and organic acids of strawberries. J. Am. Soc. Hortic. Sci. 1988, 113, 770–774. [Google Scholar]
- Shaw, D.V. Response to selection and associated changes in genetic variance for soluble solids and titratable acids contents in strawberries. J. Am. Soc. Hortic. Sci. 1990, 115, 839–843. [Google Scholar] [CrossRef]
- Spangelo, L.P.S.; Hsu, C.S.; Fejer, S.O.; Bedard, P.R.; Rousselle, G.L. Heritability and genetic variance components for 20 fruit and plant characters in the cultivated strawberry. Can. J. Genet. Cytol. 1971, 13, 443–456. [Google Scholar] [CrossRef]
- Lerceteau-Köhler, E.; Moing, A.; Guérin, G.; Renaud, C.; Petit, A.; Rothan, C.; Denoyes, B. Genetic dissection of fruit quality traits in the octoploid cultivated strawberry highlights the role of homoeo-QLT in their control. Theor. Appl. Genet. 2012, 124, 1059–1077. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, V.M.; Osorio, L.F.; Hasing, T.; Gezan, S. Estimation of genetic parameters for 12 fruit and vegetative traits in the University of Florida strawberry breeding population. J. Am. Soc. Hortic. Sci. 2012, 137, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Hasing, T.N.; Osorio, L.F.; Whitaker, V.M. With-season stability of strawberry soluble solids content. J. Am. Soc. Hortic. Sci. 2013, 138, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Camp, M.J. Temperatures after bloom affect plant growth and fruit quality of strawberry. Sci. Hortic. 2000, 85, 183–199. [Google Scholar] [CrossRef]
- Davik, J.; Bakken, A.K.; Holte, K.; Blomhoff, R. Effects of genotype and environment on total anti-oxidant capacity and the content of sugars and acids in strawberries (Fragaria × ananassa Duch.). J. Hortic. Sci. Biotechnol. 2006, 81, 1057–1063. [Google Scholar] [CrossRef]
- Del Pozo-Insfran, D.; Duncan, C.E.; Yu, K.C.; Talcott, S.T.; Chandler, C.K. Polyphenols, ascorbic acid, and soluble solids concentrations of strawberry cultivars and selections grown in a winter annual hill production system. J. Am. Soc. Hortic. Sci. 2006, 131, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Kumakura, H.; Shishido, Y. The effect of daytime, nighttime, and mean diurnal temperatures on the growth of ‘Morioka-16’ strawberry fruit and plant. J. Jap. Soc. Hortic. Sci. 1994, 62, 827–832. [Google Scholar] [CrossRef]
- Ikeda, T.; Yamazaki, K.; Kumakura, H.; Hamamoto, H. Effect of high temperature on fruit quality of pot-grown strawberry plants. Acta Hortic. 2009, 842, 679–682. [Google Scholar] [CrossRef]
- Khammayom, N.; Maruyama, N.; Chaichana, C. The effect of climatic parameters on strawberry production in a small walk-in greenhouse. AgriEngineering 2022, 4, 104–121. [Google Scholar] [CrossRef]
- Cayo, Y.D.P.; Nunes, M.C.N.; Whitaker, V.M. Effect of harvest date on the soluble solids content and sugar profile of commercial strawberry cultivars and advanced selections from the University of Florida. Proc. Fla. State Hortic. Soc. 2013, 126, 180–183. [Google Scholar]
- Cayo, Y.P.; Sargent, S.; Nunes, C.D.; Whitaker, V. Composition of commercial strawberry cultivars and advanced selections as affected by season, harvest and postharvest storage. HortScience 2016, 51, 1134–1143. [Google Scholar] [CrossRef] [Green Version]
- Kadir, S.; Sidhu, G.; Al-Khatib, K. Strawberry (Fragaria × ananassa Duch.) growth and productivity as affected by temperature. HortScience 2006, 41, 1423–1430. [Google Scholar] [CrossRef] [Green Version]
- Josuttis, M.; Dietrich, H.; Patz, C.-D.; Krüger, E. Effects of air and soil temperatures on the chemical composition of fruit and agronomic performance in strawberry (Fragaria × ananassa Duch.). J. Hortic. Sci. Biotechnol. 2011, 86, 415–421. [Google Scholar] [CrossRef]
- Agüero, J.J.; Salazar, S.M.; Kirschbaum, D.S.; Jerez, E.F. Factors affecting fruit quality in strawberries grown in a subtropical environment. Int. J. Fruit Sci. 2015, 15, 223–234. [Google Scholar] [CrossRef]
- Cárdenas-Navarro, R.; López-Pérez, L.; Lobit, P. Effect of N application season and harvest period on the production and quality of strawberry fruits (Fragaria × ananassa Duch.). Sci. Agropecu. 2019, 10, 337–345. [Google Scholar] [CrossRef] [Green Version]
- McKean, T.W. Effects of Soilless Substrate Systems and Environmental Conditions on Yield, Total Soluble Solids, and Titratable Acidity of Greenhouse Strawberry (Fragaria × ananassa). Master of Science Thesis, Ohio State University, Columbus, OH, USA, 2019. [Google Scholar]
- Samykanno, K.; Pang, E.; Marriott, P.J. Genotypic and environmental effects on flavor attributes of ‘Albion’ and ‘Juliette’ strawberry fruits. Sci. Hortic. 2013, 164, 633–642. [Google Scholar] [CrossRef]
- Cervantes, L.; Ariza, M.T.; Miranda, L.; Lozano, D.; Medina, J.J.; Soria, C.; Martínez-Ferri, E. Stability of fruit quality traits of different strawberry varieties under variable environmental conditions. Agronomy 2020, 10, 1242. [Google Scholar] [CrossRef]
- Diamanti, J.; Capocasa, F.; Mezzetti, B.; Tulipani, S.; Battino, M. The interaction of plant genotype and temperature conditions at ripening stage affects strawberry nutritional quality. Acta Hortic. 2009, 838, 183–186. [Google Scholar] [CrossRef]
- Chiang, C.; Bånkestad, D.; Hoch, G. Reaching natural growth: The significance of light and temperature fluctuations in plant performance in indoor growth facilities. Plants 2020, 9, 1312. [Google Scholar] [CrossRef] [PubMed]
- Hammer, P.A.; Urquhart, N.S. Precision and replication: Critique II. In Controlled Environment Guidelines for Plant Research; Tibbitts, T.W., Kozlowski, T.T., Eds.; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Lee, C.; Rawlings, J.O. Design of experiments in growth chambers—Uniformity trials in the North Carolina State University phytotron. Crop Sci. 1982, 22, 551–558. [Google Scholar] [CrossRef]
- Lee, S.-W.; Chae, Y.-S. Changes in fruit weight and soluble solids content of ‘Seolhyang’ strawberry by fruit setting order of the flower cluster. J. Agric. Life Sci. 2012, 46, 105–111. [Google Scholar]
- Oohashi, T.; Shigeno, T.; Kojima, N.; Hatakeyama, A.; Tsurumi, R.; Nakanishi, T.; Oohashi, Y.; Ueki, M. Effect of flower thinning on fruit quality and yield in strawberry cultivar ‘Tochigi i27 gou’. Bull. Tochigi Agric. Exp. Stn. 2018, 77, 39–50. [Google Scholar]
- Nakai, H.; Yasutake, D.; Kimura, K.; Kengo, I.; Hidaka, K.; Eguchi, T.; Hirota, T.; Okayasu, T.; Ozaki, Y.; Kitano, M. Dynamics of carbon export from leaves as translocation affected by the coordination of carbohydrate availability in field strawberry. Environ. Exp. Bot. 2022, 196, 104806. [Google Scholar] [CrossRef]
- Campbell, D.E.; Young, R. Short-term CO2 exchange response to temperature, irradiance, and CO2 concentration in strawberry. Photosynth. Res. 1986, 8, 31–40. [Google Scholar] [CrossRef]
- Bunce, J.A. Seasonal patterns of photosynthetic response and acclimation to elevated carbon dioxide in field-grown strawberry. Photosynth. Res. 2001, 68, 237–246. [Google Scholar] [CrossRef]
- Jun, H.; Jung, H.; Imai, K. Gas exchange characteristics of a leading cultivar of Korean strawberry (Fragaria × ananassa, ‘Sulhyang’). Sci. Hortic. 2017, 221, 10–15. [Google Scholar] [CrossRef]
- Rivero, R.; Sønsteby, A.; Solhaug, K.A.; Heide, O.M.; Remberg, S.F. Effects of temperature and photoperiod on photosynthesis in everbearing strawberry. Acta Hortic. 2021, 1309, 379–386. [Google Scholar] [CrossRef]
- Barrios, S.; Lemna, P.; Lareo, C. Modeling respiration rate of strawberry (cv. San Andreas) for modified atmosphere packaging design. Int. J. Food Prop. 2014, 17, 2039–2051. [Google Scholar] [CrossRef]
- Wu, X.; Han, W.; Yang, Z.; Zhang, Y.; Zheng, Y. The difference in temperature between day and night affects the strawberry soluble sugar content by influencing the photosynthesis, respiration and sucrose phosphate synthase. Hortic. Sci. 2021, 48, 174–182. [Google Scholar] [CrossRef]
- MacKenzie, S.J.; Chandler, C.K. A method to predict weekly strawberry fruit yields from extended season production systems. Agron. J. 2009, 101, 278–287. [Google Scholar] [CrossRef]
- Ayala-Zavala, J.F.; Wang, S.Y.; Wang, C.Y.; González-Aguilar, G.A. Effect of storage temperature on antioxidant capacity and aroma compounds in strawberry fruit. LWT Food Sci. Technol. 2004, 37, 687–695. [Google Scholar] [CrossRef]
- Cordenunsi, B.R.; Genovese, M.I.; Nascimento, J.R.O.; Hassimotto, N.M.A.; Santos, R.J.; Lajolo, F.M. Effects of temperature on the chemical composition and antioxidant activity of three strawberry cultivars. Food Chem. 2005, 91, 113–121. [Google Scholar] [CrossRef]
- Lv, J.; Zheng, T.; Song, Z.; Pervaiz, T.; Dong, T.; Zhang, Y.; Jia, H.; Fang, J. Strawberry proteome responses to controlled hot and cold stress partly mimic post-harvest storage temperature effects on fruit quality. Front. Nutr. 2022, 8, 812666. [Google Scholar] [CrossRef]
- Kerr, A.; Dialesandro, J.; Steenwerth, K.; Lopez-Brody, N.; Elias, E. Vulnerability of California specialty crops to projected mid-century temperature changes. Clim. Chang. 2018, 148, 419–436. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Period | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|
| 2021 | |||||||
| Mean daily maximum temperature (°C) | 25.8 | 24.0 | 21.8 | 21.9 | 23.7 | 25.2 | 28.4 |
| Mean daily minimum temperature (°C) | 15.6 | 13.4 | 11.0 | 10.3 | 11.4 | 12.9 | 16.9 |
| Mean daily solar radiation (MJ/m2) | 15.7 | 13.8 | 12.3 | 13.2 | 16.0 | 18.9 | 20.8 |
| Total monthly rainfall (mm) | 190 | 125 | 59 | 71 | 4 | 20 | 58 |
| Long-term average | |||||||
| Mean daily maximum temperature (°C) | 26.1 | 23.5 | 21.3 | 20.8 | 22.3 | 24.6 | 26.5 |
| Mean daily minimum temperature (°C) | 15.0 | 11.7 | 8.5 | 7.0 | 7.4 | 9.8 | 13.2 |
| Mean daily solar radiation (MJ/m2) | 16.2 | 13.7 | 11.7 | 13.1 | 16.1 | 18.9 | 20.9 |
| Total monthly rainfall (mm) | 160 | 108 | 115 | 50 | 58 | 90 | 80 |
| Cultivar | No. of Leaves/Plant | Leaf Area (cm2/Plant) | Leaf Dry Weight (g/Plant) | Crown Dry Weight (g/Plant) | Root Dry Weight (g/Plant) |
|---|---|---|---|---|---|
| Festival | 26.8 ± 2.0 bc | 2010 ± 161 c | 17.1 ± 1.9 b | 4.8 ± 0.9 b | 1.6 ± 0.4 bc |
| Fortuna | 28.0 ± 5.5 c | 1516 ± 120 ab | 10.7 ± 1.2 a | 3.3 ± 0.6 a | 1.4 ± 0.4 ab |
| Brilliance | 26.2 ± 2.3 bc | 1474 ± 211 ab | 10.1 ± 0.8 a | 3.3 ± 0.4 a | 1.1 ± 0.1 a |
| Beauty | 21.1 ± 2.5 a | 1395 ± 233 a | 11.2 ± 2.1 a | 3.4 ± 0.5 a | 1.5 ± 0.3 bc |
| Red Rhapsody | 24.2 ± 5.6 ab | 1728 ± 368 bc | 15.4 ± 3.8 b | 4.4 ± 0.7 b | 1.8 ± 0.1 c |
| Cultivar | Marketable Yield (g/Plant) | Fruit Weight (g) | Soluble Solids Content (%) | Titratable Acidity (%) |
|---|---|---|---|---|
| Festival | 493 ± 58 b | 21.2 ± 0.6 a | 7.9 ± 0.2 b | 0.63 ± 0.01 d |
| Fortuna | 483 ± 13 b | 22.5 ± 0.6 b | 7.2 ± 0.3 a | 0.57 ± 0.01 b |
| Brilliance | 440 ± 53 b | 23.8 ± 0.8 c | 7.1 ± 0.2 a | 0.54 ± 0.02 a |
| Beauty | 367 ± 64 a | 21.5 ± 1.0 ab | 8.2 ± 0.2 c | 0.61 ± 0.01 c |
| Red Rhapsody | 507 ± 42 b | 25.0 ± 1.0 d | 7.5 ± 0.1 b | 0.61 ± 0.02 c |
| Cultivar | Fructose (mg/g FW) | Glucose (mg/g FW) | Sucrose (mg/g FW) | Total Sugars (mg/g FW) |
|---|---|---|---|---|
| Festival | 31.5 ± 3.6 c | 28.2 ± 4.3 c | 0 ± 0 a | 59.7 ± 7.8 c |
| Fortuna | 28.1 ± 3.6 a | 24.7 ± 3.8 a | 0.1 ± 0.5 a | 53.1 ± 7.4 a |
| Brilliance | 28.2 ± 3.6 a | 25.4 ± 4.3 ab | 0 ± 0 a | 53.6 ± 7.8 a |
| Beauty | 33.1 ± 3.7 d | 30.4 ± 4.0 d | 0.2 ± 0.7 a | 63.9 ± 7.7 d |
| Red Rhapsody | 30.1 ± 4.2 b | 26.6 ± 4.7 b | 0 ± 0 a | 56.6 ± 8.8 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menzel, C.M. Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia. Horticulturae 2022, 8, 367. https://doi.org/10.3390/horticulturae8050367
Menzel CM. Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia. Horticulturae. 2022; 8(5):367. https://doi.org/10.3390/horticulturae8050367
Chicago/Turabian StyleMenzel, Christopher Michael. 2022. "Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia" Horticulturae 8, no. 5: 367. https://doi.org/10.3390/horticulturae8050367
APA StyleMenzel, C. M. (2022). Effect of Temperature on Soluble Solids Content in Strawberry in Queensland, Australia. Horticulturae, 8(5), 367. https://doi.org/10.3390/horticulturae8050367