
Use of Piriformospora indica to Promote Growth of Strawberry Daughter Plants

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Fungi Preparation
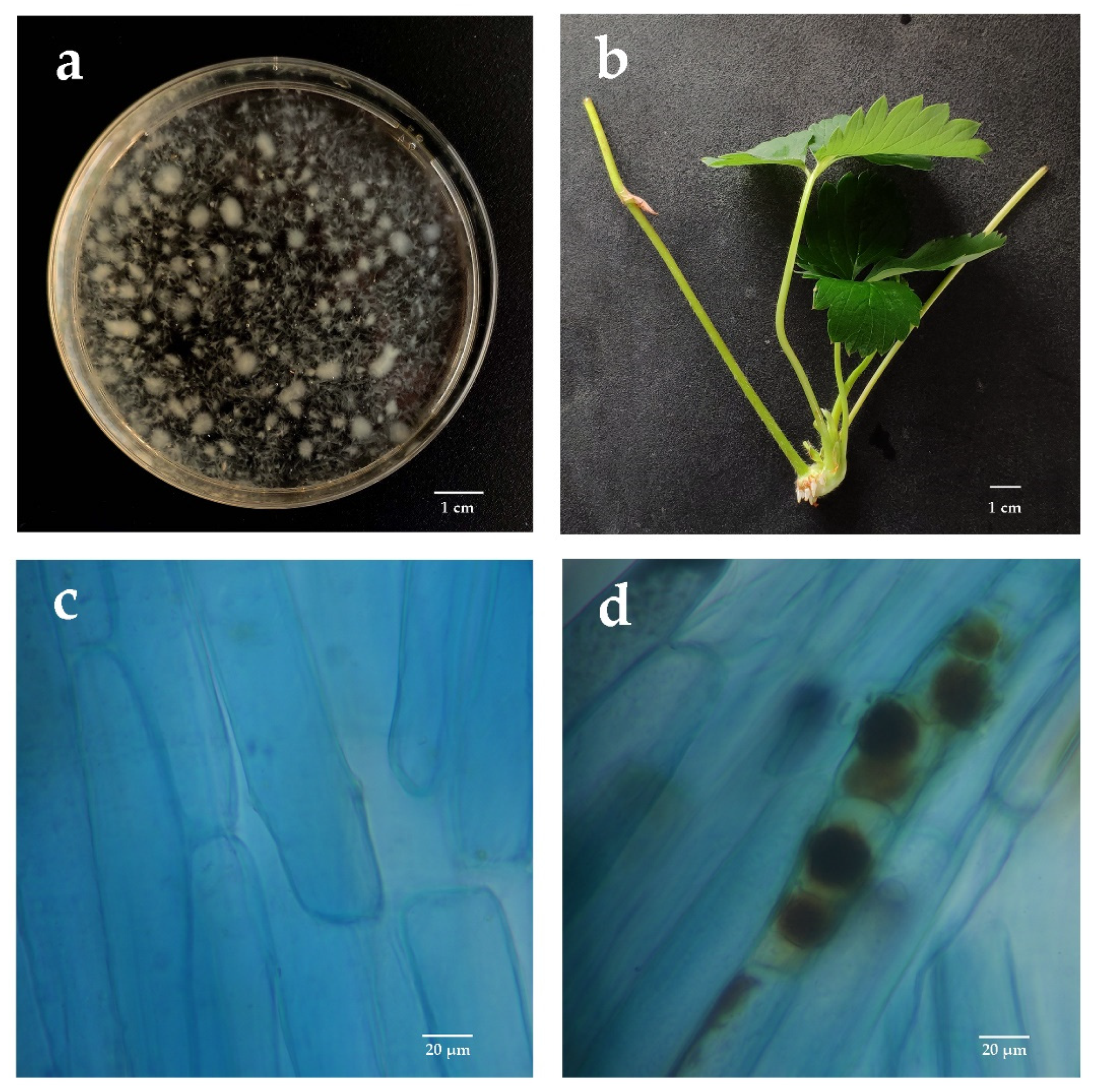
2.2. Detection of Piriformospora indica Colonization in Strawberry Roots
2.3. Measurement of Plant Growth-Related Parameters
2.4. Determination of Photosynthetic Pigments Contents
2.5. Determination of Leaf Nitrate Reductase Activity and Root Activity
2.6. Statistics Analysis
3. Results
3.1. P. indica Colonization Detection Results in Roots of Strawberry Daughter Plants
3.2. Effects of P. indica Colonization on the Growth of Strawberry Daughter Plants
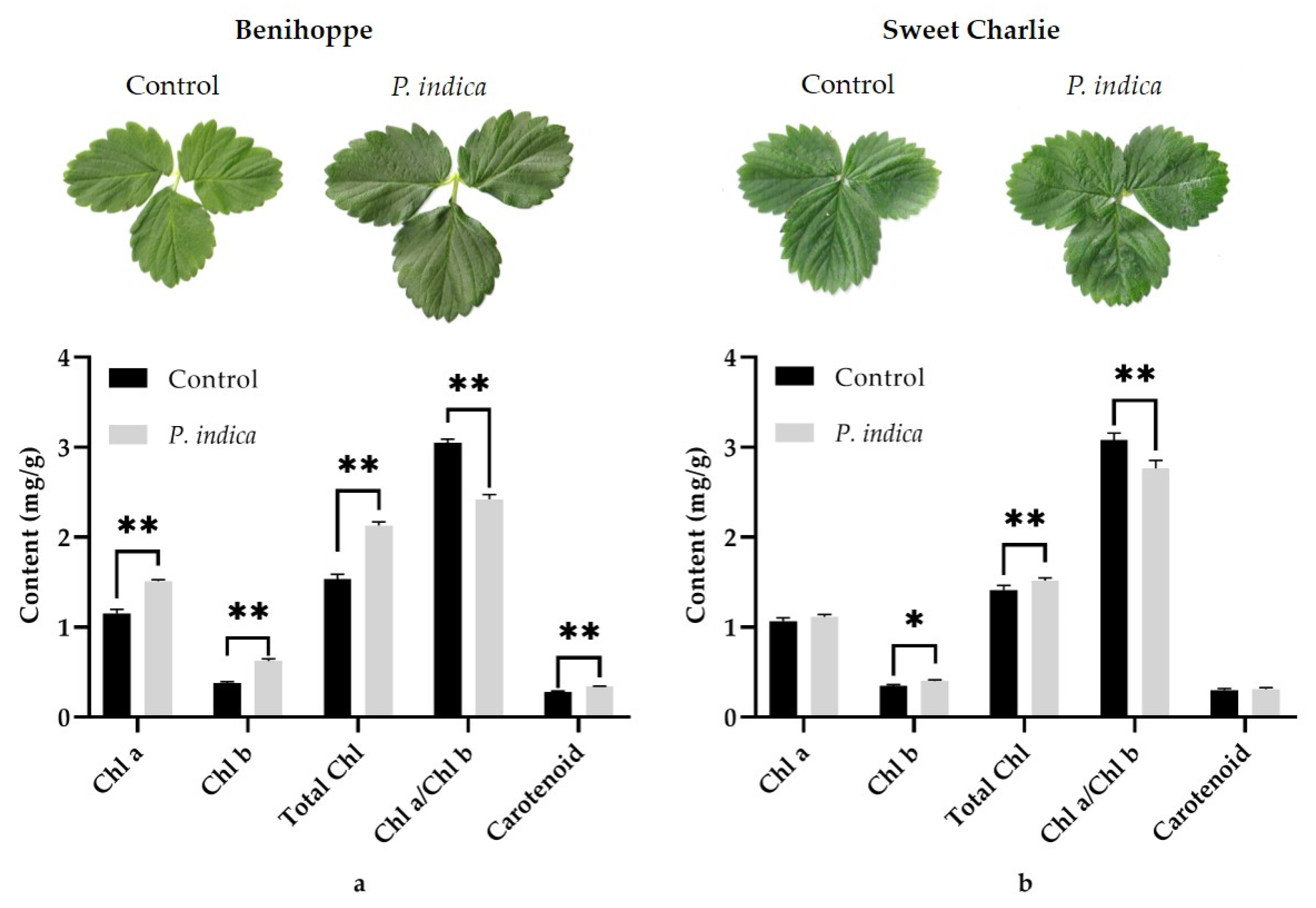
3.3. Effects of P. indica on Photosynthetic Pigments Accumulations in Leaves of Strawberry Daughter Plants
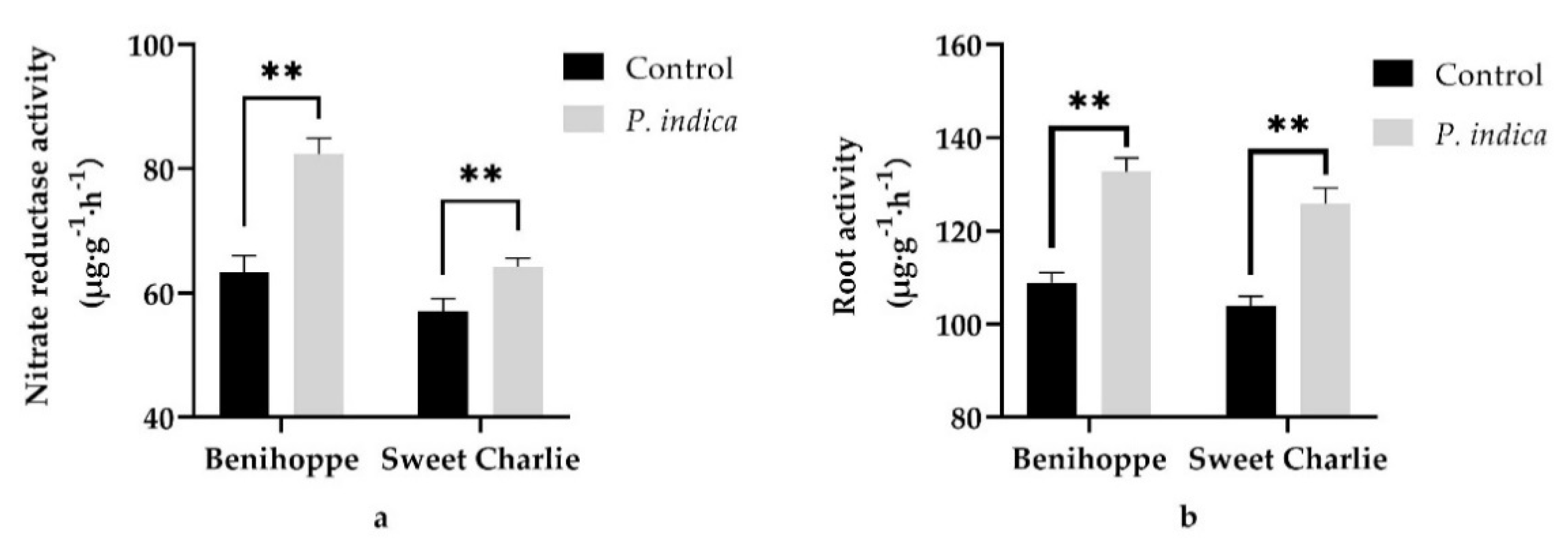
3.4. Effects of P. indica on Nitrate Reductase Activity and Root Activity
4. Discussion
4.1. Colonization of P. indica Promoted the Growth of Strawberry Daughter Plants, and Its Promoting Effects Varied in Different Varieties
4.2. P. indica Colonization Significantly Induces the Accumulation of Photosynthetic Pigments in Strawberry Leaves
4.3. P. indica Colonization Enhanced the Nutrient Uptake Ability of Strawberry Daughter Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Hrynkiewicz, K.; Baum, C. The potential of rhizosphere microorganisms to promote the plant growth in disturbed soils. In Environmental Protection Strategies for Sustainable Development; Malik, A., Grohmann, E., Eds.; Springer: Berlin, Germany, 2012; pp. 35–64. [Google Scholar]
- Jain, A.; Chakraborty, J.; Das, S. Underlying mechanism of plant-microbe crosstalk in shaping microbial ecology of the rhizosphere. Acta Physiol. Plant. 2020, 42, 8. [Google Scholar] [CrossRef]
- Shao, Y.D.; Zhang, D.J.; Hu, X.C.; Wu, Q.S.; Jiang, C.J.; Xia, T.J.; Gao, X.B.; Kuča, K. Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 2018, 64, 283–289. [Google Scholar]
- Rocha, I.; Ma, Y.; Carvalho, M.F.; Magalhães, C.; Janoušková, M.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed coating with inocula of arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria for nutritional enhancement of maize under different fertilisation regimes. Arch. Agron. Soil Sci. 2019, 65, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Cobb, A.B.; Wilson, G.W.T.; Goad, C.L.; Grusak, M.A. Influence of alternative soil amendments on mycorrhizal fungi and cowpea production. Heliyon 2018, 4, e00704. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; Iapichino, G.; Consentino, B.B.; D’Anna, F.; Rouphael, Y. Rootstock and arbuscular mycorrhiza combinatorial effects on eggplant crop performance and fruit quality under greenhouse conditions. Agronomy 2020, 10, 693. [Google Scholar] [CrossRef]
- Hart, M.; Ehret, D.L.; Krumbein, A.; Leung, C.; Murch, S.; Turi, C.; Franken, P. Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 2015, 25, 359–376. [Google Scholar] [CrossRef]
- Li, F.; Hao, Z.P.; Chen, B.D. Molecular mechanism for the adaption of arbuscular mycorrhizal symbiosis to phosphorus deficiency. J. Plant Nutr. Fertil. 2019, 25, 1989–1997. [Google Scholar]
- Rivero, J.; Álvarez, D.; Flors, V.; Azcón-Aguilar, C.; Pozo, M.J. Root metabolic plasticity underlies functional diversity in mycorrhiza-enhanced stress tolerance in tomato. New Phytol. 2018, 220, 1322–1336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.; Guo, J. Arbuscular mycorrhizal fungi improve plant growth of Ricinus communis by altering photosynthetic properties and increasing pigments under drought and salt stress. Ind. Crop. Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Mensah, R.A.; Li, D.; Liu, F.; Tian, N.; Sun, X.; Hao, X.; Lai, Z.; Cheng, C. Versatile Piriformospora indica and its potential applications in horticultural crops. Hortic. Plant J. 2020, 6, 111–121. [Google Scholar] [CrossRef]
- Varma, A.; Verma, S.; Sudha; Sahay, N.; Bütehorn, B.; Franken, P. Piriformospora indica, a cultivable plant-growth-promoting root endophyte. Appl. Environ. Microbiol. 1999, 65, 2741–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, F.; Wang, B.; Wu, H.; Wu, J.; Liu, J.; Sun, Y.; Cheng, C.; Qiu, D. Identification, characterization and expression analysis of anthocyanin biosynthesis-related bHLH genes in blueberry (Vaccinium corymbosum L.). Int. J. Mol. Sci. 2021, 22, 13274. [Google Scholar] [CrossRef] [PubMed]
- Mosse, B. Fructifications associated with mycorrhizal strawberry roots. Nature 1953, 171, 974. [Google Scholar] [CrossRef]
- Boyer, L.R.; Feng, W.; Gulbis, N.; Hajdu, K.; Harrison, R.J.; Jeffries, P.; Xu, X. The use of arbuscular mycorrhizal fungi to improve strawberry production in coir substrate. Front. Plant Sci. 2016, 7, 1237. [Google Scholar]
- Cordeiro, E.C.N.; De Resende, J.T.V.; Córdova, K.R.V.; Nascimento, D.A.; Júnior, O.J.S.; Zeist, A.R.; Favaro, R. Arbuscular mycorrhizal fungi action on the quality of strawberry fruits. Hortic. Bras. 2019, 37, 437–444. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Husaini, A.M.; Abdin, M.Z.; Khan, S.; Xu, Y.W.; Aquil, S.; Anis, M. Modifying strawberry for better adaptability to adverse impact of climate change. Curr. Sci. 2012, 102, 1660–1673. [Google Scholar]
- Chien, Y.; Lin, N. Effects of biohardening with Serendipita indica on strawberry growth and resistance to Colletotrichum gloeosporioides. J. Plant Med. 2018, 60, 1–8. [Google Scholar]
- Riens, B.; Heldt, H.W. Decrease of nitrate reductase activity in spinach leaves during a light-dark transition. Plant Physiol. 1992, 98, 573–577. [Google Scholar] [CrossRef]
- Lv, T.; Yang, H.; Zhang, R.; Fan, W.; Xu, Y.; Cao, H.; Ning, L.; Zhou, C.; Wang, L. Effects of lignin on root activity and soil nutrients of Malus hupehensis. var. pingyiensis under the use of organic fertilizer. Agric. Sci. 2017, 8, 341–347. [Google Scholar]
- Cheng, C.; Li, D.; Qi, Q.; Sun, X.; Anue, M.R.; David, B.M.; Zhang, Y.; Hao, X.; Zhang, Z.; Lai, Z. The root endophytic fungus Serendipita indica improves resistance of Banana to Fusarium oxysporum f. sp. cubense tropical race 4. Eur. J. Plant Pathol. 2020, 156, 87–100. [Google Scholar] [CrossRef]
- Rai, M.; Acharya, D.; Singh, A.; Varma, A. Positive growth responses of the medicinal plants Spilanthes calva and Withania somnifera to inoculation by Piriformospora indica in a field trial. Mycorrhiza 2001, 11, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kharkwal, A.C.; Abdin, M.Z.; Varma, A. Piriformospora indica improves micropropagation, growth and phytochemical content of Aloe vera L. plants. Symbiosis 2014, 64, 11–23. [Google Scholar] [CrossRef]
- Salah, S.M.; Yajing, G.; Dongdong, C.; Jie, L.; Aamir, N.; Qijuan, H.; Weimin, H.; Mingyu, N.; Jin, H. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 2015, 5, 14278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Gharbi, A.; Hipkin, C.R. Studies on nitrate reductase in British angiosperms: I. A comparison of nitrate reductase activity in ruderal, woodland-edge and woody species. New Phytol. 1984, 97, 629–639. [Google Scholar] [CrossRef]
- Vyas, S.; Nagori, R.; Purohit, S. Root colonization and growth enhancement of micropropagated Feronia limonia (L.) swingle by Piriformospora indica—A cultivable root endophyte. Int. J. Plant Dev. Biol. 2008, 2, 128–132. [Google Scholar]
- Bagde, U.S.; Prasad, R.; Varma, A. Interaction of mycobiont: Piriformospora indica with medicinal plants and plants of economic importance. Afr. J. Biotechnol. 2010, 9, 9214–9226. [Google Scholar]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Meng, L.L.; Liu, R.C.; Yang, L.; Zou, Y.N.; Srivastava, A.K.; Kuča, K.; Hashem, A.; Abd-Allah, E.F.; Giri, B.; Wu, Q.S. The change in fatty acids and sugars reveals the association between trifoliate orange and endophytic fungi. J. Fungi 2021, 7, 716. [Google Scholar] [CrossRef]
- Li, D.; Bodjrenou, D.M.; Zhang, S.; Wang, B.; Pan, H.; Yeh, K.W.; Lai, Z.; Cheng, C. The endophytic fungus Piriformospora indica reprograms banana to cold resistance. Int. J. Mol. Sci. 2021, 22, 4973. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kuo, Y.W.; Lin, K.H.; Huang, W.; Deng, C.; Yeh, K.W.; Chen, S.P. Piriformospora indica colonization increases the growth, development, and herbivory resistance of sweet potato (Ipomoea batatas L.). Plant Cell Rep. 2021, 40, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, M.E.; Abdelsattar, M.; Abdeldaym, E.A.; Atia, M.A.M.; Mahmoud, A.W.M.; Saad, M.M.; Hirt, H. Piriformospora indica alters Na+/K+ homeostasis, antioxidant enzymes and LeNHX1 expression of greenhouse tomato grown under salt stress. Sci. Hortic. 2019, 256, 108532. [Google Scholar] [CrossRef]
- Dong, S.; Tian, Z.; Chen, P.J.; Kumar, R.S.; Shen, C.H.; Cai, D.; Oelmüller, R.; Yeh, K.W. The maturation zone is an important target of Piriformospora indica in Chinese cabbage roots. J. Exp. Bot. 2013, 64, 4529–4540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jogawat, A.; Saha, S.; Bakshi, M.; Dayaman, V.; Kumar, M.; Dua, M.; Varma, A.; Oelmüller, R.; Tuteja, N.; Johri, A.K. Piriformospora indica rescues growth diminution of rice seedlings during high salt stress. Plant Signal. Behav. 2013, 8, e26891. [Google Scholar] [CrossRef] [Green Version]
- Varma, A.; Bakshi, M.; Lou, B.; Hartmann, A.; Oelmueller, R. Piriformospora indica: A novel plant growth-promoting mycorrhizal fungus. Agric. Res. 2012, 1, 117–131. [Google Scholar] [CrossRef]
- Moreira, B.C.; Mendes, F.C.; Mendes, I.R.; Paula, T.A.; Prates Junior, P.; Salomão, L.C.C.; Stürmer, S.L.; Otoni, W.C.; Guarçoni, M.A.; Kasuya, M.C.M. The interaction between arbuscular mycorrhizal fungi and Piriformospora indica improves the growth and nutrient uptake in micropropagation-derived pineapple plantlets. Sci. Hortic. 2015, 197, 183–192. [Google Scholar] [CrossRef]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmüller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar]
- Cheng, C.; Liu, F.; Wang, B.; Qu, P.; Liu, J.; Zhang, Y.; Liu, W.; Tong, Z.; Deng, G. Influences of Serendipita indica and Dictyophorae echinovolvata on the growth and Fusarium wilt disease resistance of banana. Biology 2022, 11, 393. [Google Scholar] [CrossRef]
- Shende, S.; Bhagwat, K.; Wadegaonkar, P.; Rai, M.; Varma, A.; Rai, M.K. Piriformospora indica as a new and emerging mycofertilizer and biotizer: Potentials and prospects in sustainable agriculture. In Handbook of Microbial Biofertilizers, 1st ed.; Food Products Press: Binghamton, NY, USA, 2005; pp. 477–496. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | Benihoppe | Sweet Charlie | ||
|---|---|---|---|---|
| CK | Pi | CK | Pi | |
| Above-ground fresh weight (g) | 2.91 ± 0.50 | 4.29 ± 0.55 * | 4.36 ± 0.74 | 5.92 ± 1.25 * |
| Root fresh weight (g) | 3.11 ± 0.51 | 4.45 ± 0.50 * | 4.08 ± 0.91 | 5.44 ± 0.69 * |
| Above-ground dry weight (g) | 0.79 ± 0.12 | 1.18 ± 0.15 * | 1.08 ± 0.15 | 1.37 ± 0.19 |
| Root dry weight (g) | 0.54 ± 0.10 | 0.83 ± 0.13 * | 0.74 ± 0.13 | 1.03 ± 0.20 * |
| Petiole length (cm) | 7.65 ± 1.51 | 10.63 ± 0.71 * | 9.86 ± 1.94 | 11.33 ± 1.23 |
| Leaf area (cm2) | 22.57 ± 1.94 | 25.97 ± 1.08 * | 24.84 ± 1.60 | 23.04 ± 1.41 |
| Plant height (cm) | 13.63 ± 1.51 | 15.91 ± 0.62 * | 15.05 ± 0.99 | 16.45 ± 1.68 |
| Root length (cm) | 18.08 ± 1.71 | 15.5 ± 1.43 | 17.88 ± 1.95 | 13.48 ± 0.91 * |
| Root number | 6.60 ± 0.93 | 10.80 ± 2.27 * | 10.00 ± 2.10 | 10.20 ± 1.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Tan, M.; Qu, P.; Huo, C.; Liang, W.; Li, R.; Jia, Y.; Fan, X.; Cheng, C. Use of Piriformospora indica to Promote Growth of Strawberry Daughter Plants. Horticulturae 2022, 8, 370. https://doi.org/10.3390/horticulturae8050370
Liu W, Tan M, Qu P, Huo C, Liang W, Li R, Jia Y, Fan X, Cheng C. Use of Piriformospora indica to Promote Growth of Strawberry Daughter Plants. Horticulturae. 2022; 8(5):370. https://doi.org/10.3390/horticulturae8050370
Chicago/Turabian StyleLiu, Wei, Min Tan, Pengyan Qu, Chensi Huo, Wenjie Liang, Runlin Li, Yan Jia, Xinping Fan, and Chunzhen Cheng. 2022. "Use of Piriformospora indica to Promote Growth of Strawberry Daughter Plants" Horticulturae 8, no. 5: 370. https://doi.org/10.3390/horticulturae8050370
APA StyleLiu, W., Tan, M., Qu, P., Huo, C., Liang, W., Li, R., Jia, Y., Fan, X., & Cheng, C. (2022). Use of Piriformospora indica to Promote Growth of Strawberry Daughter Plants. Horticulturae, 8(5), 370. https://doi.org/10.3390/horticulturae8050370