
Changes of Fruit Abscission and Carbohydrates, Hormones, Related Gene Expression in the Fruit and Pedicel of Macadamia under Starvation Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Starvation Stress Simulationby Girdling and Defoliation Treatment and Sample Collection
2.3. Investigation of Fruit Set
2.4. Determination of Carbohydrate Composition
2.5. Determination of Endogenous Hormones
2.6. Gene Expression by qRT-PCR Analysis
2.7. Statistical Analysis
3. Results and Analysis
3.1. Effect of Starvation Stress on Young Fruit Abscission
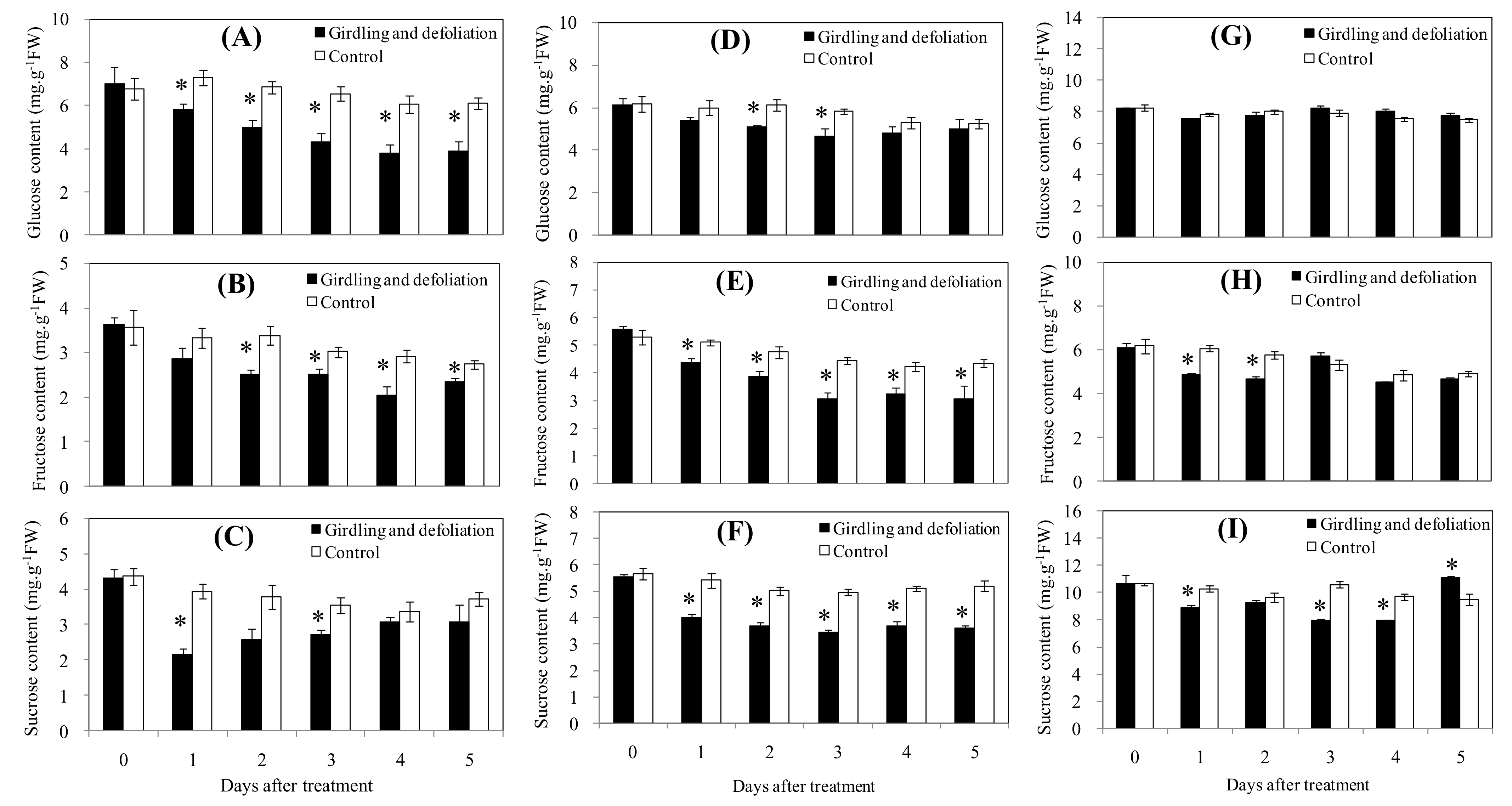
3.2. Effect of Starvation Stress on Carbohydrates in Fruit Tissues and Pedicel
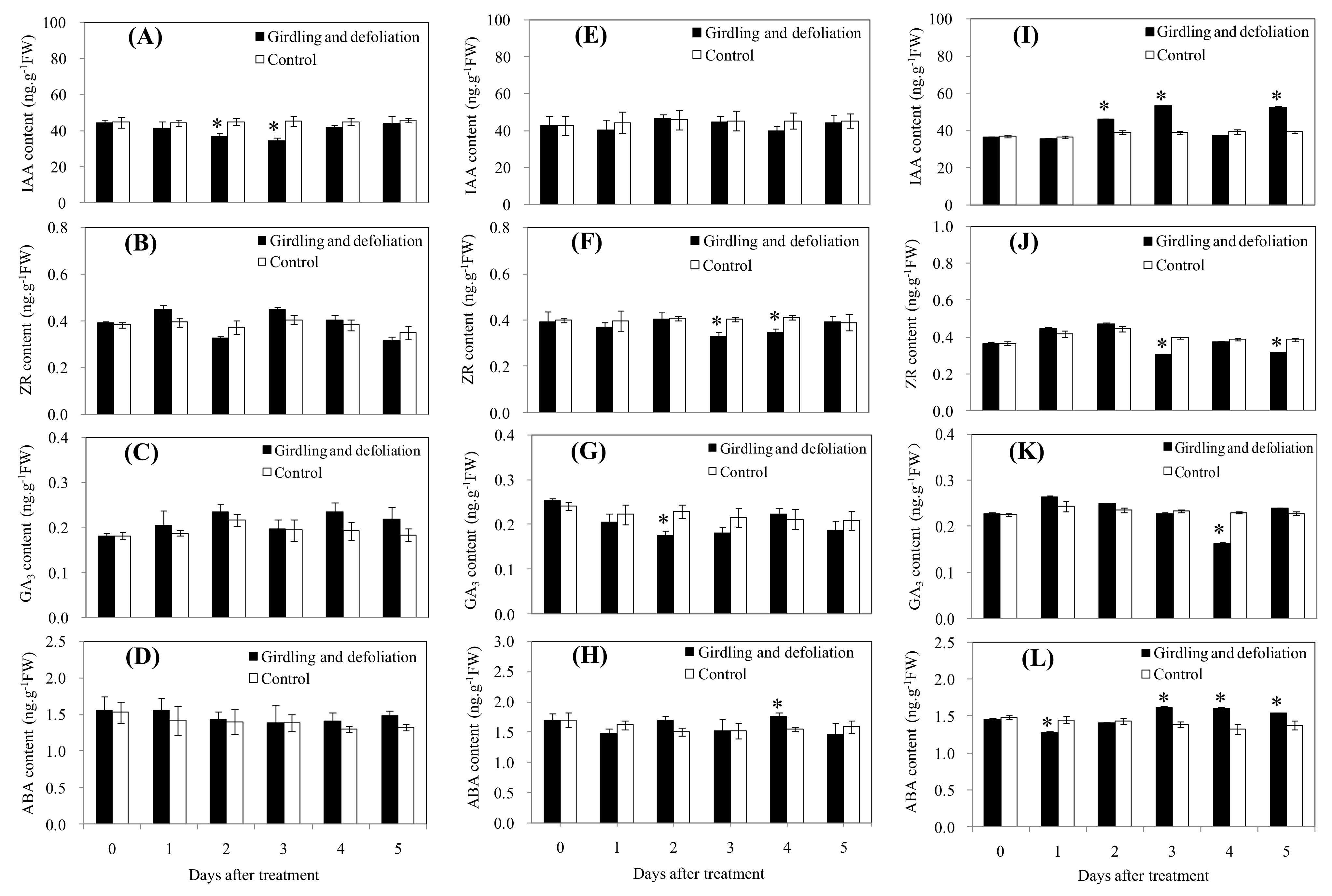
3.3. Effect of Starvation Stress on the Endogenous Hormonesin Fruit Tissues and Pedicel
3.4. Effect of Starvation Stress on the Gene Expression in Fruit Tissues and Pedicel
4. Discussion
4.1. Effects of Sugar on Fruit Abscission
4.2. Effects of Endogenous Hormones on Fruit Abscission
4.3. Crosstalk between Sugar and Hormones under Starvation Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAO | Abscisic aldehyde oxidase |
| ABA | Abscisic acid |
| ABI | Abscisic acid insensitive 5 |
| ACO | 1-Aminocyclopropane-1-carboxylate oxidase |
| ACS | 1-Aminocyclopropane-1-carboxylate synthase |
| AP2/ERF | APETALA2/Ethylene Responsive Factor |
| Aux/IAA | Auxin/indole-3-acetic acid protein |
| bZIP | Basic leucine zipper |
| CTK | Cytokinin |
| CPPU | N-(2-Chloro-4-pyridyl)-N’-phenylurea |
| ELISA | Enzyme-linked immune sorbent assay |
| GA | Gibberellic acid |
| GH3 | Gretchen Hagen 3 |
| HD-ZIP | Homeodomain-leucine zipper |
| HPLC | High-performance liquid chromatography |
| HXK | Hexokinase |
| IAA | Indole-3-acetic acid |
| NCED | 9-cis-epoxycarotenoid dioxygenase |
| NI | Neutral invertase |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| TPP | Trehalose-6-phosphate phosphatase |
| TPS | Trehalose-6-phosphate synthase |
| ZR | Zeatin-riboside |
References
- Ito, P.J. Effect of style removal on fruit set in Macadamia. HortScience 1980, 15, 520–521. [Google Scholar]
- Trueman, S.J.; Turnbull, C.G.N. Fruit set, abscission and dry matter accumulation on girdled branches of Macadamia. Ann. Bot. 1994, 74, 667–674. [Google Scholar] [CrossRef]
- Taylor, J.E.; Whitelaw, C.A. Signals in abscission. New Phytol. 2001, 151, 323–329. [Google Scholar] [CrossRef]
- Lakso, A.N.; Robinson, T.L.; Greene, D.W. Integration of environment, physiology and fruit abscission via carbon balance modeling-implications for understanding growth regulator responses. Acta Hortic. 2006, 727, 321–326. [Google Scholar] [CrossRef]
- Picken, A.J.F. A review of pollination and fruit set in the tomato (Lycopersicon esculentum Mill.). J. Hortic. Sci. 2015, 59, 1–13. [Google Scholar] [CrossRef]
- McAtee, P.; Karim, S.; Schaffer, R.J.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, L.E.; Paul, M.J.; Wingler, A. How do sugars regulate plant growth and development? New insight into the role of trehalose-6-phosphate. Mol. Plant 2013, 6, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Lordan, J.; Reginato, G.H.; Lakso, A.N.; Francescatto, P.; Robinson, T.L. Natural fruitlet abscission as related to apple tree carbon balance estimated with the MaluSim model. Sci. Hortic. 2019, 247, 296–309. [Google Scholar] [CrossRef]
- Zhao, M.; Li, J. Molecular events involved in fruitlet abscission in litchi. Plants 2020, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, D.J.; Tadeo, F.R.; Primo-Millo, E.; Talon, M. Fruit set dependence on carbohydrate availability in citrus trees. Tree Physiol. 2003, 23, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, D.J.; Tadeo, F.R.; Primo-Millo, E.; Talon, M. Carbohydrate and ethylene levels related to fruitlet drop through abscission zone A in citrus. Trees 2006, 20, 348–355. [Google Scholar] [CrossRef]
- Yang, Z.; Li, M.; Zhang, X.; Yu, Y.; Wang, H.; Huang, X. Effects of starvation stress on fruit abscission and sugar metabolism in longan. J. Fruit Sci. 2011, 28, 428–432. [Google Scholar]
- Hagemann, M.H.; Winterhagen, P.; Roemer, M.G.; Hegele, M.; Wünsche, J.N. Proposed physiological mechanism of mango fruitlet abscission. Acta Hortic. 2016, 1119, 73–80. [Google Scholar] [CrossRef]
- Stopar, M.; Resnik, M.; Pongrac, V.Z. Non-structural carbohydrate status and CO2 exchange rate of apple fruitlets at the time of abscission influenced by shade, NAA or BA. Sci. Hortic. 2001, 87, 65–76. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Huang, X.; Li, J.; Wang, H.; Li, J. De novo assembly and characterization of fruit transcriptome in Litchi chinensis Sonn and analysis of differentially regulated genes in fruit in response to shading. BMC Genom. 2013, 14, 552. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Dardick, C.D.; Beers, E.P.; Callanhan, A.M.; Xia, R.; Yuan, R. Transcriptomics of shading-induced and NAA-induced abscission in apple (Malus domestica) reveals a shared pathway involving reduced photosynthesis, alterations in carbohydrate transport and signaling and hormone crosstalk. BMC Plant Biol. 2011, 11, 138–157. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cadenas, A.; Mehouachi, J.; Tadeo, F.R.; Primo-Millo, E.; Talon, M. Hormonal regulation of fruitlet abscission induced by carbohydrate shortage in citrus. Planta 2000, 210, 636–643. [Google Scholar] [CrossRef]
- Yang, Z.; Zhong, X.; Fan, Y.; Wang, H.; Li, J.; Huang, X. Burst of reactive oxygen species in pedicel-mediated fruit abscission after carbohydrate supply was cut off in longan (Dimocarpus longan). Front. Plant Sci. 2015, 6, 360. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zeng, L.; Xiao, Q.; Shi, S. Changes of fruit abscission and carbohydrate, ABA and related genes expression in the pericarp and fruit abscission zone of longan under starvation stress. Acta Hortic. Sin. 2021, 48, 1457–1469. [Google Scholar]
- Zeng, H.; Yang, W.; Lu, C.; Lin, W.; Zou, M.; Zhang, H.; Wan, J.; Huang, X. Effect of CPPU on carbohydrate and endogenous hormone levels in young macadamia fruit. PLoS ONE 2016, 11, e0158705. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.H.; Lu, C.Z.; Chen, W.; Xu, H.Y. Reduction of early fruit abscission by main-branch-girdling in macadamia is related to the favorable status of carbohydrates and endogenous hormones. Hortscience 2022, 57, 40–47. [Google Scholar] [CrossRef]
- Huang, Y.J.; Ma, P.Q.; Wu, W.; Chen, J.Z.; Li, J.; Tang, X.L.; Liu, X.Y. Effects of summer shoot growth on sugar metabolism and abscission of fruitlet in ‘Shatangju’ mandarin. Acta Hortic. Sin. 2013, 40, 1869–1876. [Google Scholar]
- Guo, C.M.; Yang, B.; Mubareke, A.; Che, Y.; Xu, J.; Gong, P.; Xu, Y.; Liao, K. The sucrose metabolism dynamics in different tissues of almond during the physiological fruit drop and its relations with fruit drop. Plant Physiol. J. 2020, 56, 317–326. [Google Scholar]
- Zhou, C.; Lakso, A.N.; Robinson, T.L.; Gan, S. Isolation and characterization of genes associated with shade-induced apple abscission. Mol. Genet. Genom. 2008, 280, 83–92. [Google Scholar] [CrossRef]
- Botton, A.; Eccher, G.; Forcato, C.; Ferrarini, A.; Begheldo, M.; Zermiani, M.; Moscatello, S.; Battistelli, A.; Velasco, R.; Ruperti, B.; et al. Signalling pathways mediating the induction of apple fruitlet abscission. Plant Physiol. 2011, 155, 185–208. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wang, Y.; Huang, X.; Li, J.; Wang, H.; Li, J. An improved fruit transcriptome and the identification of the candidate genes involved in fruit abscission induced by carbohydrate stress in litchi. Front. Plant Sci. 2015, 6, 439. [Google Scholar] [CrossRef] [Green Version]
- Eccher, G.; Botton, A.; Dimauro, M.; Boschetti, A.; Ruperti, B.; Ramina, A. Early induction of apple fruitlet abscission is characterized by an increase of both isoprene emission and abscisic acid content. Plant Physiol. 2013, 161, 112–120. [Google Scholar]
- Han, J.X.; Zheng, H.; Zhang, Q.; Zhong, C. Research advances in the metabolism and regulation of carbohydrate in fruit trees. Plant Sci. J. 2020, 38, 143–149. [Google Scholar]
- Li, C.; Wang, Y.; Ying, P.; Ma, W.; Li, J. Genome-wide digital transcript analysis of putative fruitlet abscission related genes regulated by ethephon in litchi. Front. Plant Sci. 2015, 6, 502. [Google Scholar] [CrossRef] [Green Version]
- Price, J.; Laxmi, A.; StMartin, S.K.; Jang, J.C. Global transcription profiling reveals multiple sugar signal transduction mechanisms in Arabidopsis. Plant Cell 2004, 16, 2128–2150. [Google Scholar] [CrossRef] [Green Version]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Roitsch, T.; Gonzalez, M.C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci. 2004, 9, 606–613. [Google Scholar] [CrossRef]
- Ruan, Y.L.; Jin, Y.; Yang, Y.J.; Li, G.J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef]
- Bihmidine, S.; Hunter, C.T.; Johns, C.E.; Koch, K.E.; Braun, D.M. Regulation of assimilate import into sink organs: Update on molecular drivers of sink strength. Front. Plant Sci. 2013, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.I.; Ryoo, N.; Eom, J.S.; Lee, D.W.; Kim, H.B.; Jeong, S.W.; Lee, Y.H.; Kwon, Y.K.; Cho, M.H.; Bhoo, S.H.; et al. Role of the rice hexokinases OsHXK5 and OsHXK6 as glucose sensors. Plant Physiol. 2009, 149, 745–759. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.H.; Hong, J.W.; Kim, E.C.; Yoo, S.D. Regulatory functions of SnRK1 in stress responsive gene expression and in plant growth and development. Plant Physiol. 2012, 158, 1955–1964. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Offler, C.E.; Ruan, Y.L. Regulation of fruit and seed response to heat and drought by sugars as nutrients and signals. Front. Plant Sci. 2013, 4, 282. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef] [Green Version]
- Yadav, U.P.; Lvakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.M.; Stitt, M.; et al. The sucrose-trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot. 2014, 65, 1051–1068. [Google Scholar] [CrossRef] [Green Version]
- Wilmowicz, E.; Frankowski, K.; Kućko, A.; Świdziński, M.; de Dios, A.J.; Nowakowska, A.; Kopcewicz, J. The influence of abscisic acid on the ethylene biosynthesis pathway in the functioning of the flower abscission zone in Lupinus luteus. J. Plant Growth Regul. 2016, 206, 49–58. [Google Scholar] [CrossRef]
- Wu, J.Y.; Li, C.Q.; Li, J.G. Isolation of ACS1 gene and the relationship between its expression and fruitlet abscission in litchi. J. Fruit Sci. 2017, 34, 817–827. [Google Scholar]
- Iglesias, D.J.; Cercos, M.; Colmenero-Flore, J.M.; Naranjo, M.A.; Rios, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Kuang, J.F.; Wu, J.Y.; Zhong, H.Y.; Li, C.Q.; Chen, J.Y.; Lu, W.J.; Li, J.G. Carbohydrate stress affecting fruitlet abscission and expression of genes related to auxin signal transduction pathway in litchi. Int. J. Mol. Sci. 2012, 13, 16084–16103. [Google Scholar] [CrossRef]
- Xie, R.; Ge, T.; Zhang, J.; Pan, X.; Ma, Y.; Yi, S.; Zheng, Y. The molecular events of IAA inhibiting citrus fruitlet abscission revealed by digital gene expression profiling. Plant Physiol. Biochem. 2018, 130, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; He, Z.; Yang, B.; Li, G.; Yao, L. Effect of girdling on litchi foliar nutrient and development of flower and fruit. Guangdong Agric. Sci. 2018, 45, 34–42. [Google Scholar]
- Shinozaki, Y.; Hao, S.; Kojima, M.; Sakakibara, H.; Ozeki-Iida, Y.; Zheng, Y.; Fei, Z.; Zhong, S.; Giovannoni, J.J.; Rose, J.K.; et al. Ethylene suppresses tomato (Solanum lycopersicum) fruit set through modification of gibberellin metabolism. Plant J. 2015, 83, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Else, M.A.; Stankiewicz-Davies, A.P.; Crisp, C.M.; Atkinson, C.J. The role of polar auxin transport through pedicels of Prunus avium L. in relation to fruit development and retention. J. Exp. Bot. 2004, 55, 2099–2109. [Google Scholar] [CrossRef] [Green Version]
- Meir, S.; Philosoph-Hadas, S.; Sundaresan, S.; Selvaraj, K.S.V.; Burd, S.; Ophir, R.; Kochanek, B.; Reid, M.S.; Jiang, C.Z.; Lers, A. Microarray analysis of the abscission-related transcriptome in the tomato flower abscission zone in response to auxin depletion. PlantPhysiol. 2010, 154, 1929–1956. [Google Scholar] [CrossRef]
- Xie, R.; Pang, S.; Ma, Y.; Deng, L.; He, S.; Yi, S.; Lv, Q.; Zheng, Y. The ARF, AUX/IAA and GH3 gene families in citrus: Genome-wide identification and expression analysis during fruitlet drop from abscission zone A. Mol. Genet. Genom. 2015, 290, 2089–2105. [Google Scholar] [CrossRef]
- Trainotti, L.; Tadiello, A.; Casadoro, G. The involvement of auxin in the ripening of climacteric fruits comes of age: The hormone plays a role of its own and has an intense interplay with ethylene in ripening peaches. J. Exp. Bot. 2007, 58, 3299–3308. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Frasse, P.; Olmos, E.; Zegzouti, H.; Li, Z.G.; Latche, A.; Pech, J.C.; Bouzayen, M. Down-regulation of DR12, an auxin-response-factor homolog, in the tomato results in a pleiotropic phenotype including dark green and blotchy ripening fruit. Plant J. 2002, 32, 603–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, M.M.; González-Carranza, Z.H.; Azam-Ali, S.; Tang, S.; Shahid, A.A.; Roberts, J.A. The manipulation of auxin in the abscission zone cells of Arabidopsis flowers reveals that indoleacetic acid signaling is a prerequisite for organ shedding. Plant Physiol. 2013, 162, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abebie, B.; Lers, A.; Philosoph-Hadas, S.; Goren, R.; Riov, J.; Meir, S. Differential effects of NAA and 2,4-D in reducing floret abscission in Cestrum (Cestrum elegans) cut flowers are associated with their differential activation of AUX/IAA homologous genes. Ann. Bot. 2008, 101, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Meir, S.; Hunter, D.A.; Chen, J.; Halaly, V.; Reid, M.S. Molecular changes occurring during acquisition of abscission competence following auxin depletion in Mirabilis jalapa. Plant Physiol. 2006, 141, 1604–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-aceticacid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangerth, F. Dominance among fruits/sinks and the search for a correlative signal. Physiol. Plant. 1989, 76, 608–614. [Google Scholar] [CrossRef]
- Abel, S.; Nguyen, M.D.; Chow, W.; Theologis, A. ASC4, a primary indoleacetic acid-responsive gene encoding 1-Aminocyclopropane-1-carboxylate synthase in Arabidopsis thaliana. J. Biol. Chem. 1995, 270, 19093–19099. [Google Scholar] [CrossRef] [Green Version]
- Solfanclli, C.; Poggi, A. Sucrose-specific induction of the anthocyaninbiosyntheticpathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.H.; Endo, A.; Zhou, L.; Penney, J.; Chen, H.C.; Arroyo, A.; Leon, P.; Nambara, E.; Asami, T.; Seo, M.; et al. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 2002, 14, 2723–2743. [Google Scholar] [CrossRef]
- Cho, Y.H.; Sheen, J.; Yoo, S.D. Low glucose uncouples hexokinase1-dependent sugar signaling from stress and defense hormone abscisic acid and C2H4 responses in Arabidopsis. Plant Physiol. 2010, 152, 1180–1182. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Ann. Rev. Plant Physiol. 2006, 57, 781–803. [Google Scholar]
- Zhao, W.; Guan, C.; Feng, J.; Liang, Y.; Zhan, N.; Zuo, J.; Ren, B. The Arabidopsis CROWDED NUCLEI genes regulate seed germination by modulating degradation of ABI5 protein. J. Integr. Plant Biol. 2016, 58, 669–678. [Google Scholar] [CrossRef]
- Li, C.; Zhao, M.; Ma, X.; Wen, Z.; Ying, P.; Peng, M.; Ning, X.; Xia, R.; Wu, H.; Li, J. The HD-Zip transcription factor LcHB2 regulates litchi fruit abscission through the activation of two cellulase genes. J. Exp. Bot. 2019, 70, 5189–5203. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.G.; Price, J.; Lin, P.C.; Hong, J.C.; Jang, J.C. The arabidopsis bZIP1 transcription factor is involved in sugar signaling, protein networking and DNA binding. Mol. Plant 2010, 3, 361–373. [Google Scholar]
- Joo, J.; Lee, Y.H.; Song, S.I. Overexpression of thericebasic leucine zipper transcription factorOsbZIP12confers drought tolerance toriceand makes seedlings hypersensitive to ABA. Plant Biotechnol. Rep. 2014, 8, 431–441. [Google Scholar] [CrossRef]
- Heo, S.; Hwang, J.H.; Jun, J.H.; Lee, H.J. Abscission-related genes revealed by RNA-Seq analysis using self-abscising apple (Malus × domestica). J. Pomol. Hortic. Sci. 2016, 91, 271–278. [Google Scholar] [CrossRef]
- Gil-Amado, J.A.; Gomez-Jimenez, M.C. Transcriptome analysis of mature fruit abscission control in olive. Plant Cell Physiol. 2013, 54, 244–269. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer Sequence (5′→3′) | Reverse Primer Sequence (5′→3′) | Length/bp |
|---|---|---|---|
| NADPH5 | CAGTGCCAGAAGTATTCAACCA | CAATGCGACCAAAACCGT | 116 |
| NI | GCTTAGGCTTGGCTATCTTCTT | CCAGAATACTATGACGGGAAGAC | 166 |
| HXK2 | ATCAAATGTTGCGGAATGGG | TTCTGAAGGCGGGAGTAAGC | 195 |
| TPS | TCCCCTTAAAATACCAGCGTG | CGGAGAACCCATCTTTGAGC | 96 |
| TPP | AGATGACCGAACAGATGAAGATG | AATGCGTTGCTTTCTTTTGG | 99 |
| ACO2 | GTGATAGCCCAAACAGACGG | GGATAAACCACCTGGCATTG | 158 |
| ACS | TTTGGAGAAACTGGACATAGCC | CCCTTGAGAATAAGACCTTGGAT | 91 |
| NCED1.1 | CTTCATTCTGTGATTTGGGCTAC | TGGAGGACTGGAGGAGTTTGT | 103 |
| AAO | GTGCTTCAAGACCTTCCGTG | CAGGAGGGAAGAACATAGGAAT | 177 |
| BG | GCCACGTCTCCATTGCTTT | TTCCACCAGGTTTCTATTTCG | 148 |
| GH3.9 | AGACGAAGAAGATGAGGAGGTG | ACTGGGGTGCTTTGTTGTAGA | 164 |
| Aux/IAA22 | TATGGCATCGGTGGGTTGT | TCCTTAGCCTTTTGCATGACTC | 136 |
| HDZIP12 | TCCAGAAGTGAACCCGAACC | TGCCCAGACCACTCAGGAAT | 110 |
| bZIP124 | TTAACGCAGGACTCCGTATCG | ACCCATCGTCAGTGAGCCAT | 132 |
| AP2/ERF | GTGGCTGGGGACATTTGAT | GTAACATAAGCGGCAGGCA | 192 |
| ABI5 | CTACCGTGTATGCCTGTTTCC | AATGGGGAGTTGTTACAGGGT | 164 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Xiang, P. Changes of Fruit Abscission and Carbohydrates, Hormones, Related Gene Expression in the Fruit and Pedicel of Macadamia under Starvation Stress. Horticulturae 2022, 8, 398. https://doi.org/10.3390/horticulturae8050398
Yang W, Xiang P. Changes of Fruit Abscission and Carbohydrates, Hormones, Related Gene Expression in the Fruit and Pedicel of Macadamia under Starvation Stress. Horticulturae. 2022; 8(5):398. https://doi.org/10.3390/horticulturae8050398
Chicago/Turabian StyleYang, Weihai, and Peijin Xiang. 2022. "Changes of Fruit Abscission and Carbohydrates, Hormones, Related Gene Expression in the Fruit and Pedicel of Macadamia under Starvation Stress" Horticulturae 8, no. 5: 398. https://doi.org/10.3390/horticulturae8050398