
Electrolytic Disinfection of Irrigation Water for Intensive Crop Production in Greenhouses as Demonstrated on Tomatoes (Solanum lycopersicum Mill)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Plant Pathogens and Inoculum
2.2. Detection of Plant Pathogens in Plant Material
2.3. Detection of Plant Pathogens in Nutrient Solution
2.4. Electrolytic Disinfection of Fertigation Solution
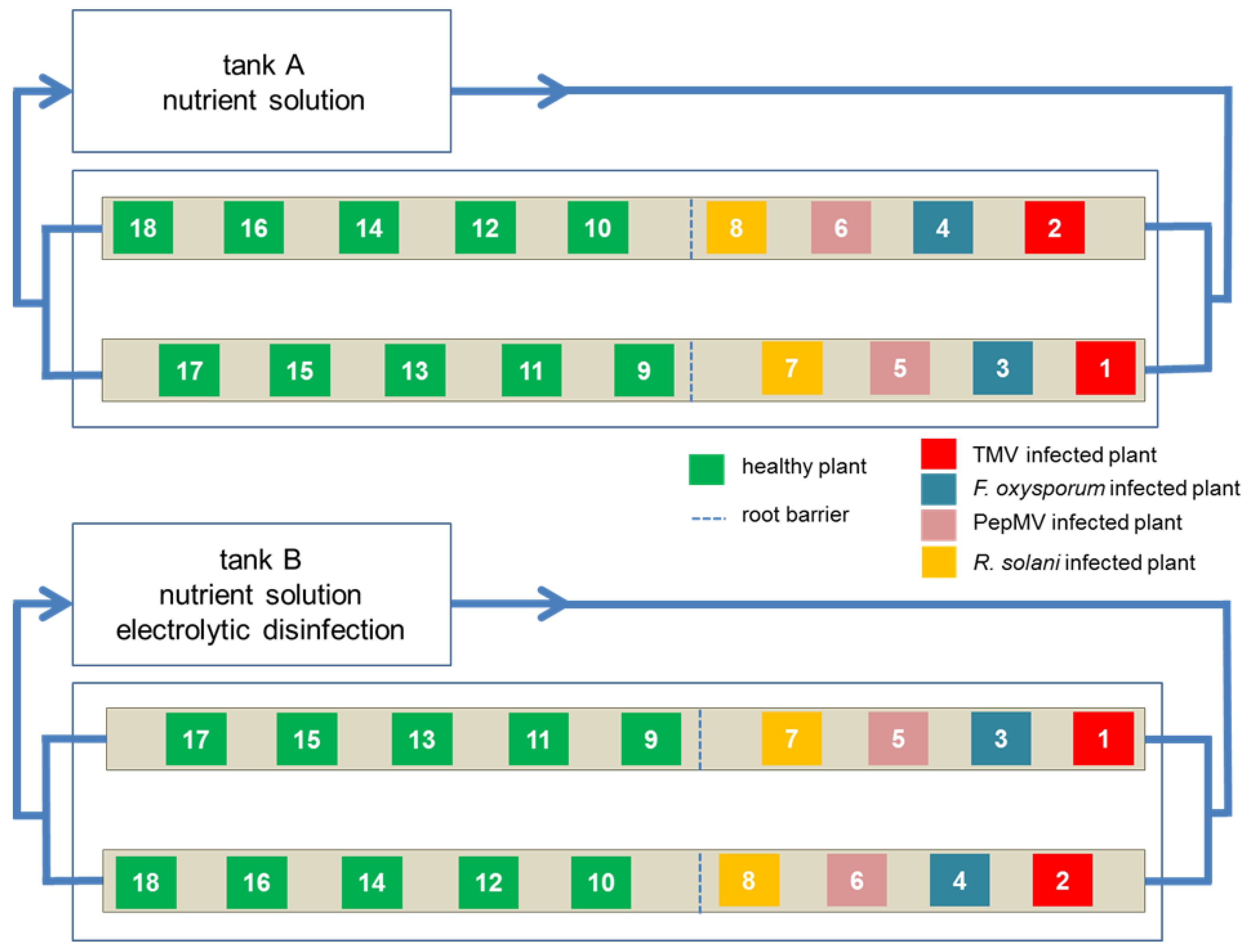
2.5. Experimental Design
2.6. Data Collection and Analysis
3. Results
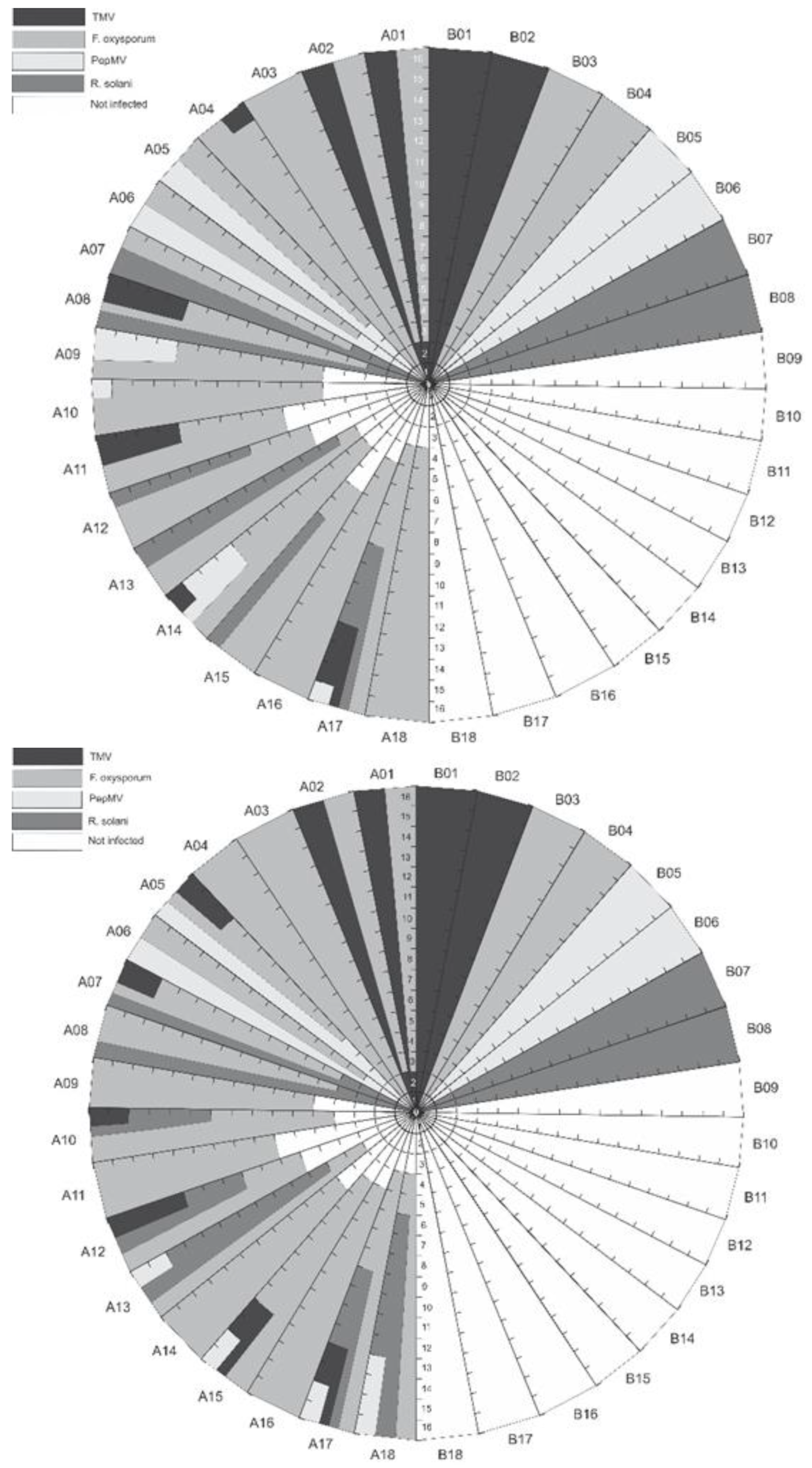
3.1. Detection of Plant Pathogens in Fertigation Solution
3.2. Effect of Fertigation Solution Treatment on Pathogen Dispersal
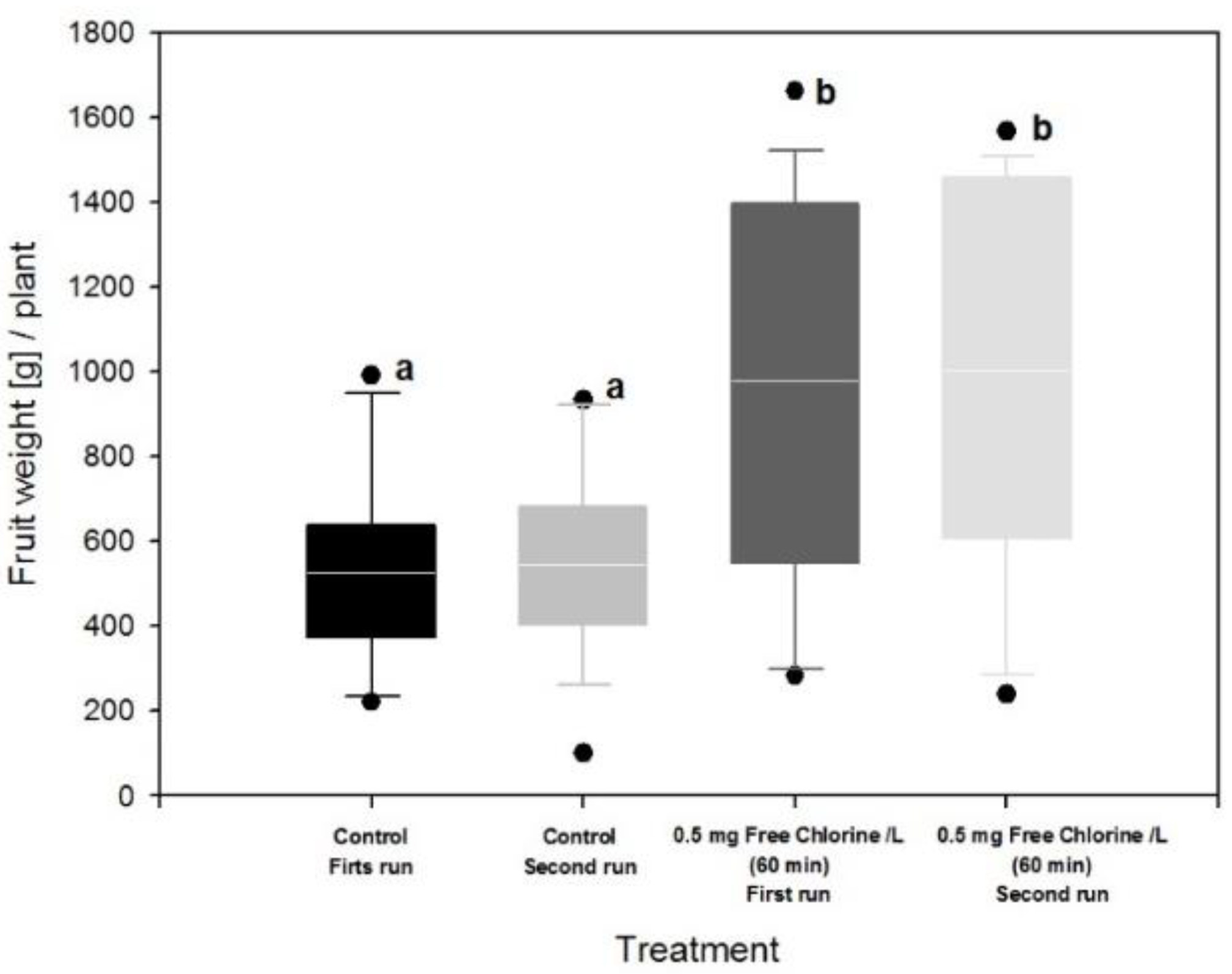
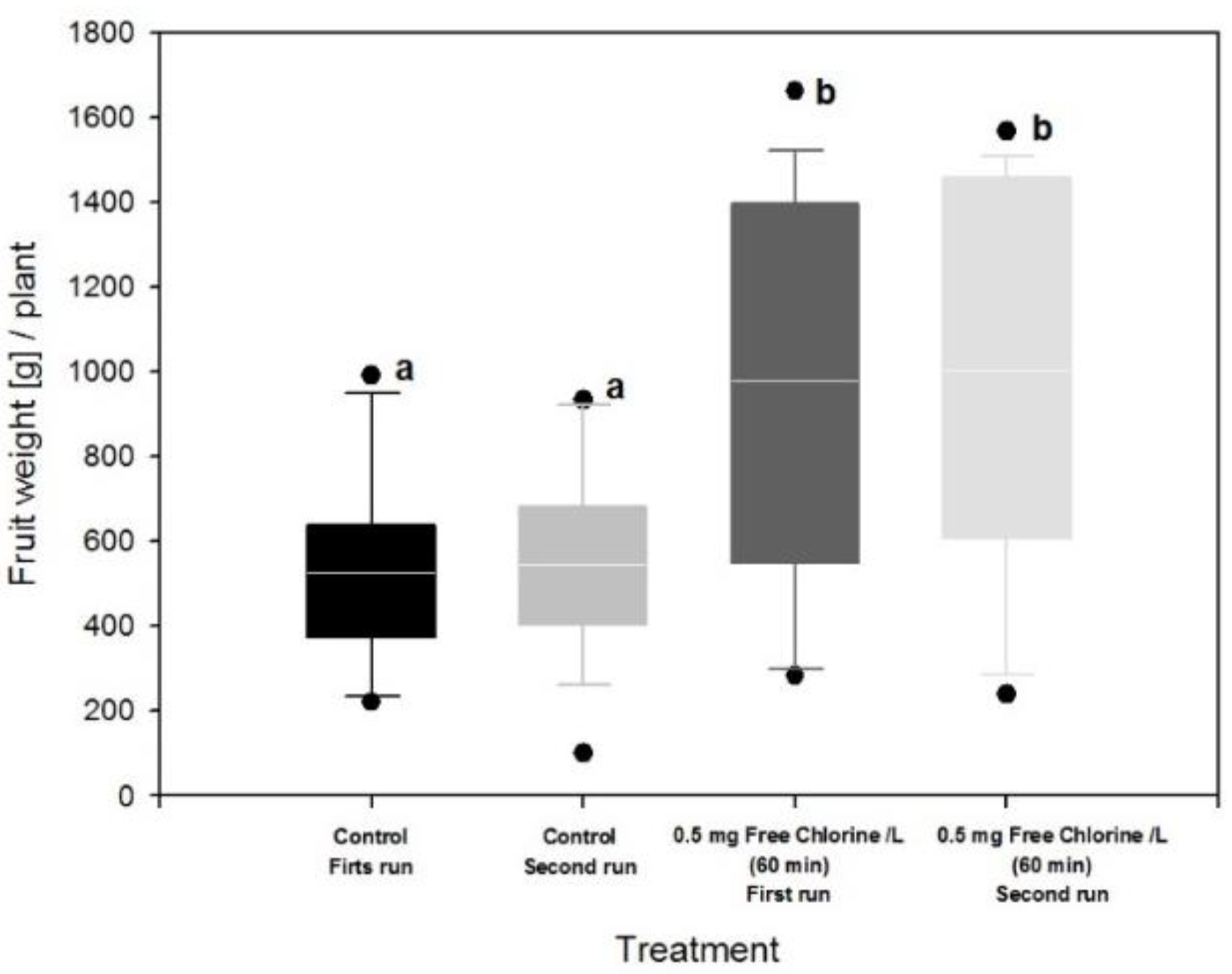
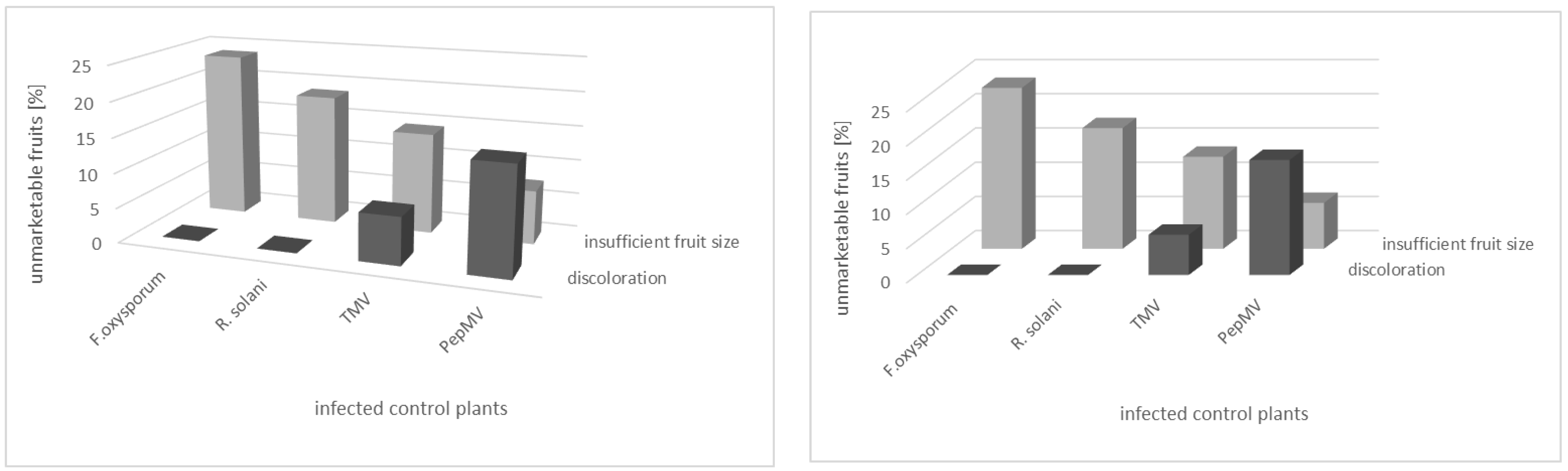
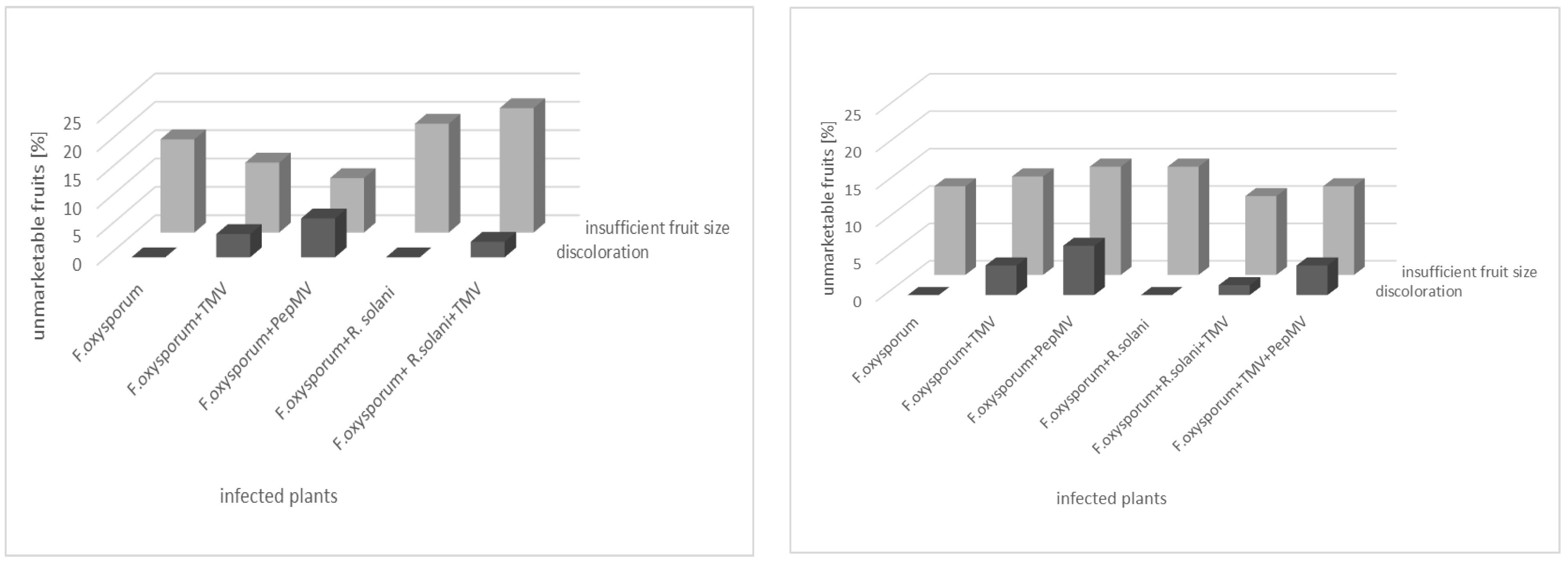
3.3. Effect of Fertigation Solution Treatment on Tomato Plant and Fruit
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture—Systems at Breaking Point; Earthscan: London, UK, 2021; p. 82. [Google Scholar]
- Lévesque, S.; Graham, T.; Bejan, D.; Lawson, J.; Zhang, P.; Dixon, M. An electrochemical advanced oxidation process (EAOP) for the inactivation of Rhizoctonia solani in fertigation solutions. Can. J. Plant Sci. 2020, 100, 415–424. [Google Scholar] [CrossRef] [Green Version]
- FAO. Towards a Water and Food Secure Future; FAO & WWC: Marseille, France; Rome, Italy, 2015; p. 76. [Google Scholar]
- FAO. Water. 2022. Available online: https://www.fao.org/water/en/ (accessed on 5 January 2022).
- Stanghellini, M.; Rasmussen, S.; Kim, D.; Rorabaugh, P. Efficacy of nonionic surfactants in the control of zoospore spread of Pythium aphanidermatum in a recirculating hydroponic system. Plant Dis. 1996, 80, 422–428. [Google Scholar] [CrossRef]
- Schnitzler, W.H. Pest and disease management of soilless culture. Acta Hortic. 2004, 648, 191–203. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Plant pathogens in recycled irrigation water in commercial plant nurseries and greenhouses: Their detection and management. Irrig. Sci. 2011, 29, 267–297. [Google Scholar] [CrossRef]
- Hong, C.; Richardson, P.; Kong, P.; Bush, E. Efficacy of chlorine on multiple species of Phytophthora in recycled nursery irrigation water. Plant Dis. 2003, 87, 1183–1189. [Google Scholar] [CrossRef]
- Hong, C.; Moorman, G. Plant pathogens in irrigation water: Challenges and opportunities. Crit. Rev. Plant Sci. 2005, 24, 189–208. [Google Scholar] [CrossRef]
- Mehle, N.; Ravnikar, M. Plant viruses in aqueous environment–survival, water mediated transmission and detection. Water Res. 2012, 46, 4902–4917. [Google Scholar] [CrossRef]
- FAO. Crops and Livestock Products; The Food and Agriculture Organization FAOstat: Roma, Italy, 2022. [Google Scholar]
- Büttner, C.; Koenig, R. Plant viruses in irrigation water. In Biology, Detection, and Management of Plant Pathogens in Irrigation Wate; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 97–110. [Google Scholar]
- Wick, R.L. Fungal plant pathogens in irrigation water. In Biology, Detection and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 77–82. [Google Scholar]
- Walker, J.C. Fusarium Wilt of Tomato; APS, Ed.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 1971; Volume 6, p. 56. [Google Scholar]
- Reis, A.; Costa, H.; Boiteux, L.S.; Lopes, C.A. First report of Fusarium oxysporum f. sp. lycopersici race 3 on tomato in Brazil. Fitopatol. Bras. 2005, 30, 426–428. [Google Scholar] [CrossRef] [Green Version]
- Đorđević, M.; Vatchev, T.; Girek, Z.; Šević, M.; Zečević, B.; Zdravković, J.; Ivanović, M. Reaction of different tomato cultivars toward race 1 of Fusarium oxysporum f. sp. lycopersici. Genetika 2012, 44, 109–118. [Google Scholar]
- Nelson, P.E. Life cycle and epidemiology of Fusarium oxysporum. In Fungal Wilt Diseases of Plants; Mace, M., Bell, A.A., Beckman, C.H., Eds.; Academic Press: New York, NY, USA, 1981; pp. 51–80. [Google Scholar] [CrossRef]
- Jarvis, W.; Shoemaker, R. Taxonomic status of Fusarium oxysporum causing foot and root rot of tomato. Phytopathology 1978, 68, 1679–1680. [Google Scholar] [CrossRef]
- Kapoor, I. Fungi involved in tomato wilt syndrome in Delhi, Maharashtra and Tamil Nadu. Indian Phytopathol. 1988, 41, 208–213. [Google Scholar]
- Gupta, S.K.; Thind, T. Disease Problems in Vegetable Production; Scientific Publishers Journals Department: Jodhpur, India, 2018; p. 576. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi; Academic Press: London, UK; New York, NY, USA; Toronto, ON, Canada; Sydney, Australia; San Francisco, CA, USA, 1980; p. 859. [Google Scholar]
- Ogoshi, A. Introduction—the genus Rhizoctonia. In Rhizoctonia species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 1–9. [Google Scholar] [CrossRef]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 1991; p. 133. [Google Scholar]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef]
- González García, M. Reseña de “Aspectos de sistemática y biologíca del complejo Rhizoctonia”. Fitosanidad 2008, 12, 147–159. [Google Scholar]
- Jones, R.A.C.; Koenig, R.; Lesemann, D. Pepino mosaic virus, a new potexvirus from pepino (Solanum muricatum). Ann. Appl. Biol. 1980, 94, 61–68. [Google Scholar] [CrossRef]
- Wright, D.; Mumford, R. Pepino mosaic Potexvirus (PepMV): First records in tomato in the United Kingdom. Plant Dis. Not. 1999, 89, 400. [Google Scholar]
- van der Vlugt, R.A.; Stijger, C.I.; Verhoeven, J.T.J.; Lesemann, D.E. First Report of Pepino mosaic virus on Tomato. Plant Dis. 2000, 84, 103. [Google Scholar] [CrossRef]
- Cordoba-Selles, M.d.C.; García-Rández, A.; Alfaro-Fernández, A.; Jordá-Gutiérrez, C. Seed transmission of Pepino mosaic virus and efficacy of tomato seed disinfection treatments. Plant Dis. 2007, 91, 1250–1254. [Google Scholar] [CrossRef] [Green Version]
- Lacasa, A.; Guerrero, M.; Hita, I.; Martínez, M.; Hernández, M. La diseminación del virus del mosaico del pepino dulce (Pepino mosaic Virus) en las labores de entutorado y desbrotado de las plantas de tomate. Bol. San. Veg. Plagas 2001, 21, 489–501. [Google Scholar]
- Spence, N.; Basham, J.; Mumford, R.; Hayman, G.; Edmondson, R.; Jones, D. Effect of Pepino mosaic virus on the yield and quality of glasshouse-grown tomatoes in the UK. Plant Pathol. 2006, 55, 595–606. [Google Scholar] [CrossRef]
- Schwarz, D.; Beuch, U.; Bandte, M.; Fakhro, A.; Büttner, C.; Obermeier, C. Spread and interaction of Pepino mosaic virus (PepMV) and Pythium aphanidermatum in a closed nutrient solution recirculation system: Effects on tomato growth and yield. Plant Pathol. 2010, 59, 443–452. [Google Scholar] [CrossRef]
- Jordá, C.; Pérez, A.L.; Martínez-Culebras, P.; Abad, P.; Lacasa, A.; Guerrero, M. First report of Pepino mosaic virus on tomato in Spain. Plant Dis. 2001, 85, 1292. [Google Scholar] [CrossRef]
- Hanssen, I.; Paeleman, A.; Vandewoestijne, E.; Van Bergen, L.; Bragard, C.; Lievens, B.; Vanachter, A.; Thomma, B. Pepino mosaic virus isolates and differential symptomatology in tomato. Plant Pathol. 2009, 58, 450–460. [Google Scholar] [CrossRef]
- Roggero, P.; Masenga, V.; Lenzi, R.; Coghe, F.; Ena, S.; Winter, S. First report of Pepino mosaic virus in tomato in Italy. Plant Pathol. 2001, 50, 798. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Paeleman, A.; Wittemans, L.; Goen, K.; Lievens, B.; Bragard, C.; Vanachter, A.C.; Thomma, B.P. Genetic characterization of Pepino mosaic virus isolates from Belgian greenhouse tomatoes reveals genetic recombination. Eur. J. Plant Pathol. 2008, 121, 131–146. [Google Scholar] [CrossRef]
- Mayer, A. Concerning the mosaic disease of tobacco. In Phytopathological Classics; Johnson, J., Ed.; APS, The American Phytopathological Society: St. Paul, MN, USA, 1886; Volume 7, pp. 11–24. [Google Scholar]
- Jones, J.; Jones, J.; Stall, R.; Zitter, T. Compendium of Tomato Diseases, 1st ed.; APS, The American Phytopathological Society: St. Paul, MN, USA, 1991; p. 73. [Google Scholar]
- Scholthof, K. Tobacco mosaic virus. In The Plant Health Instructor; APS, The American Phytopathological Society: St. Paul, MN, USA, 2005. [Google Scholar] [CrossRef]
- Hong, C. Component analyses of irrigation water in plant disease epidemiology. In Biology, Detection and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014. [Google Scholar] [CrossRef]
- De Hayr, R.; Bodman, K.; Forsberg, L. Bromine and chlorine disinfestation of nursery water supplies. Comb. Proc. Int. Plant Propag. Soc. 1994, 44, 60–66. [Google Scholar]
- Fisher, P.R.; Huang, J.; Raudales, R.E.; Meador, D.P. Chlorine for irrigation water treatment. In Biology, Detection, and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 235–249. [Google Scholar] [CrossRef]
- Nirenberg, H. Studies on the morphological and biological differentiation in the Fusarium-section Liseola. In Mitteilungen der Biologischen Bundeanstalt für Land-und Forstwirtschaft; P. Parey: Berlin, Germany, 1976; Volume 169, pp. 1–117. [Google Scholar]
- Büttner, C.; Bandte, M.; Pettitt, T.R. Filtration and centrifugation for detection of plant pathogens in irrigation water. In Biology, Detection and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 139–148. [Google Scholar] [CrossRef]
- Bandte, M.; Rodriguez, M.H.; Schuch, I.; Schmidt, U.; Buettner, C. Plant viruses in irrigation water: Reduced dispersal of viruses using sensor-based disinfection. Irrig. Sci. 2016, 34, 221–229. [Google Scholar] [CrossRef]
- Schuch, I.; Dannehl, D.; Bandte, M.; Suhl, J.; Gao, Y.; Schmidt, U. Minimising chlorate in the electrolytic disinfection of irrigation water. Landtechnik 2016, 71, 25–33. [Google Scholar]
- Rodriguez, M.H.; Bandte, M.; Gaskin, T.; Fischer, G.; Büttner, C. Efficacy of electrolytically-derived disinfectant against dispersal of Fusarium oxysporum and Rhizoctonia solani in hydroponic tomatoes. Sci. Hortic. 2018, 234, 116–125. [Google Scholar] [CrossRef]
- Göhler, F.; Molitor, H.D. Erdelose Kulturverfahren im Gartenbau; Eugen Ulmer Stuttgart: Stuttgart, Germany, 2002; p. 268. [Google Scholar]
- OECD. Tomatoes. In International Standards for Fruit and Vegetables; OECD Publishing: Paris, France, 2019. [Google Scholar] [CrossRef]
- Pettitt, T.R. Irrigation water and the health of nursery crops. In Biology, Detection, and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 13–22. [Google Scholar] [CrossRef]
- Moorman, G.W.; Gevens, A.J.; Granke, L.L.; Hausbeck, M.K.; Hendricks, K.; Roberts, P.D.; Pettitt, T.R. Sources and distribution systems of irrigation water and their potential risks for crop health. In Biology, Detection, and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 3–12. [Google Scholar] [CrossRef]
- Moorman, G.W. Irrigation water and the health of greenhouse crops. In Biology, Detection, and Management of Plant Pathogens in Irrigation Water; Hong, C., Moorman, G.W., Wohanka, W., Büttner, C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2014; pp. 23–29. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Fungi and oomycetes in the irrigation water of forest nurseries. Forests 2020, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Younis, B.A.; Mahoney, L.; Schweigkofler, W.; Suslow, K. Inactivation of plant pathogens in irrigation water runoff using a novel UV disinfection system. Eur. J. Plant Pathol. 2019, 153, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Cayanan, D.F.; Zhang, P.; Liu, W.; Dixon, M.; Zheng, Y. Efficacy of chlorine in controlling five common plant pathogens. HortScience 2009, 44, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Machado, P.d.S.; Alfenas, A.C.; Coutinho, M.M.; Silva, C.M.; Mounteer, A.H.; Maffia, L.A.; de Freitas, R.G.; Freitas, C.d.S. Eradication of plant pathogens in forest nursery irrigation water. Plant Dis. 2013, 97, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehle, N.; Gutiérrez-Aguirre, I.; Prezelj, N.; Delić, D.; Vidic, U.; Ravnikar, M. Survival and transmission of potato virus Y, Pepino mosaic virus, and potato spindle tuber viroid in water. Appl. Environ. Microbiol. 2014, 80, 1455–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins Jr, S.; Averre, C. Root diseases of vegetables in hydroponic culture systems in North Carolina greenhouses. Plant Dis. 1983, 67, 968–970. [Google Scholar] [CrossRef]
- Rattink, H. Epidemiology of Fusarium wilt in cyclamen in an ebb and flow system. Neth. J. Plant Pathol. 1990, 96, 171–177. [Google Scholar] [CrossRef]
- Lévesque, S.; Graham, T.; Bejan, D.; Lawson, J.; Zhang, P.; Dixon, M. Inactivation of Rhizoctonia solani in fertigation water using regenerative in situ electrochemical hypochlorination. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Raudales, R.E.; Parke, J.L.; Guy, C.L.; Fisher, P.R. Control of waterborne microbes in irrigation: A review. Agric. Water Manag. 2014, 143, 9–28. [Google Scholar] [CrossRef]
- Haute, S.v.; Sampers, I.; Jacxsens, L.; Uyttendaele, M. Selection criteria for water disinfection techniques in agricultural practices. Crit. Rev. Food Sci. Nutr. 2015, 55, 1529–1551. [Google Scholar] [CrossRef]
- Buck, J.; Van Iersel, M.; Oetting, R.; Hung, Y.-C. Evaluation of acidic electrolyzed water for phytotoxic symptoms on foliage and flowers of bedding plants. Crop Prot. 2003, 22, 73–77. [Google Scholar] [CrossRef]
- Hakme, E.; Herrmann, S.S.; Poulsen, M.E. Chlorate and perchlorate residues in food products on the Danish market. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2022, 39, 551–559. [Google Scholar] [CrossRef]
- Dannehl, D.; Schuch, I.; Gao, Y.; Cordiner, S.; Schmidt, U. Effects of hypochlorite as a disinfectant for hydroponic systems on accumulations of chlorate and phytochemical compounds in tomatoes. Eur. Food Res. Technol. 2016, 242, 345–353. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2020/685 of 20 May 2020 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Perchlorate in Certain Foods (Text with EEA Relevance); European Comission: Brussels, Belgium, 2020; pp. 3–5.
- Commission Regulation (EU) 2020/749 of 4 June 2020 Amending Annex III to Regulation (EC) No 396/2005 of the European Parliament and of the Council as Regards Maximum Residue Levels for Chlorate in or on Certain Products (Text with EEA Relevance); European Comission: Brussels, Belgium, 2020; pp. 7–20.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Yield [Kg] | Yield by Plant [Kg] | ||||
|---|---|---|---|---|---|
| Survey | Tomato Plants | No Sanitation | 0.5 mg Cl/L | No Sanitation | 0.5 mg Cl/L |
| 1 | F. oxysporum, R. solani, TMV and PepMV-infected control plants | 3.34 | 4.17 | 0.42 ± 0.13 b | 0.52 ± 0.16 b |
| Test plants | 6.15 | 13.46 | 0.61 ± 0.28 b | 1.35 ± 0.17 a | |
| 2 | F. oxysporum, R. solani, TMV and PepMV-infected control plants | 3.55 | 4.27 | 0.44 ± 0.18 b | 0.53 ± 0.18 b |
| Test plants | 6.32 | 13.81 | 0.63 ± 0.23 b | 1.38 ± 0.14 a | |
| Mean Fruit/Plant [No.] | Unmarketable Fruit [%] | ||||
|---|---|---|---|---|---|
| Survey | Tomato Plants | No Sanitation | 0.5 mg Cl/L | No Sanitation | 0.5 mg Cl/L |
| 1 | F. oxysporum, R. solani, TMV and PepMV—infected control plants | 9.13 ± 1.55 b | 14.75 ± 1.69 ab | 93.15 | 86.44 |
| Test plants | 10.90 ± 3.04 ab | 27.40 ± 2.22 a | 63.31 | 4.38 | |
| 2 | F. oxysporum, R. solani, TMV and PepMV—infected control plants | 9.50 ± 1.60 b | 14.88 ± 2.02 ab | 92.11 | 84.04 |
| Test plants | 11 ± 2.81 ab | 28.70 ± 2.00 a | 58.19 | 4.88 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, M.H.; Schmidt, U.; Büttner, C.; Bandte, M. Electrolytic Disinfection of Irrigation Water for Intensive Crop Production in Greenhouses as Demonstrated on Tomatoes (Solanum lycopersicum Mill). Horticulturae 2022, 8, 414. https://doi.org/10.3390/horticulturae8050414
Rodriguez MH, Schmidt U, Büttner C, Bandte M. Electrolytic Disinfection of Irrigation Water for Intensive Crop Production in Greenhouses as Demonstrated on Tomatoes (Solanum lycopersicum Mill). Horticulturae. 2022; 8(5):414. https://doi.org/10.3390/horticulturae8050414
Chicago/Turabian StyleRodriguez, Marlon Hans, Uwe Schmidt, Carmen Büttner, and Martina Bandte. 2022. "Electrolytic Disinfection of Irrigation Water for Intensive Crop Production in Greenhouses as Demonstrated on Tomatoes (Solanum lycopersicum Mill)" Horticulturae 8, no. 5: 414. https://doi.org/10.3390/horticulturae8050414
APA StyleRodriguez, M. H., Schmidt, U., Büttner, C., & Bandte, M. (2022). Electrolytic Disinfection of Irrigation Water for Intensive Crop Production in Greenhouses as Demonstrated on Tomatoes (Solanum lycopersicum Mill). Horticulturae, 8(5), 414. https://doi.org/10.3390/horticulturae8050414