
Effect of Temperature and Photoperiod Preconditioning on Flowering and Yield Performance of Three Everbearing Strawberry Cultivars


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Experimental Design and Data Observations
3. Results
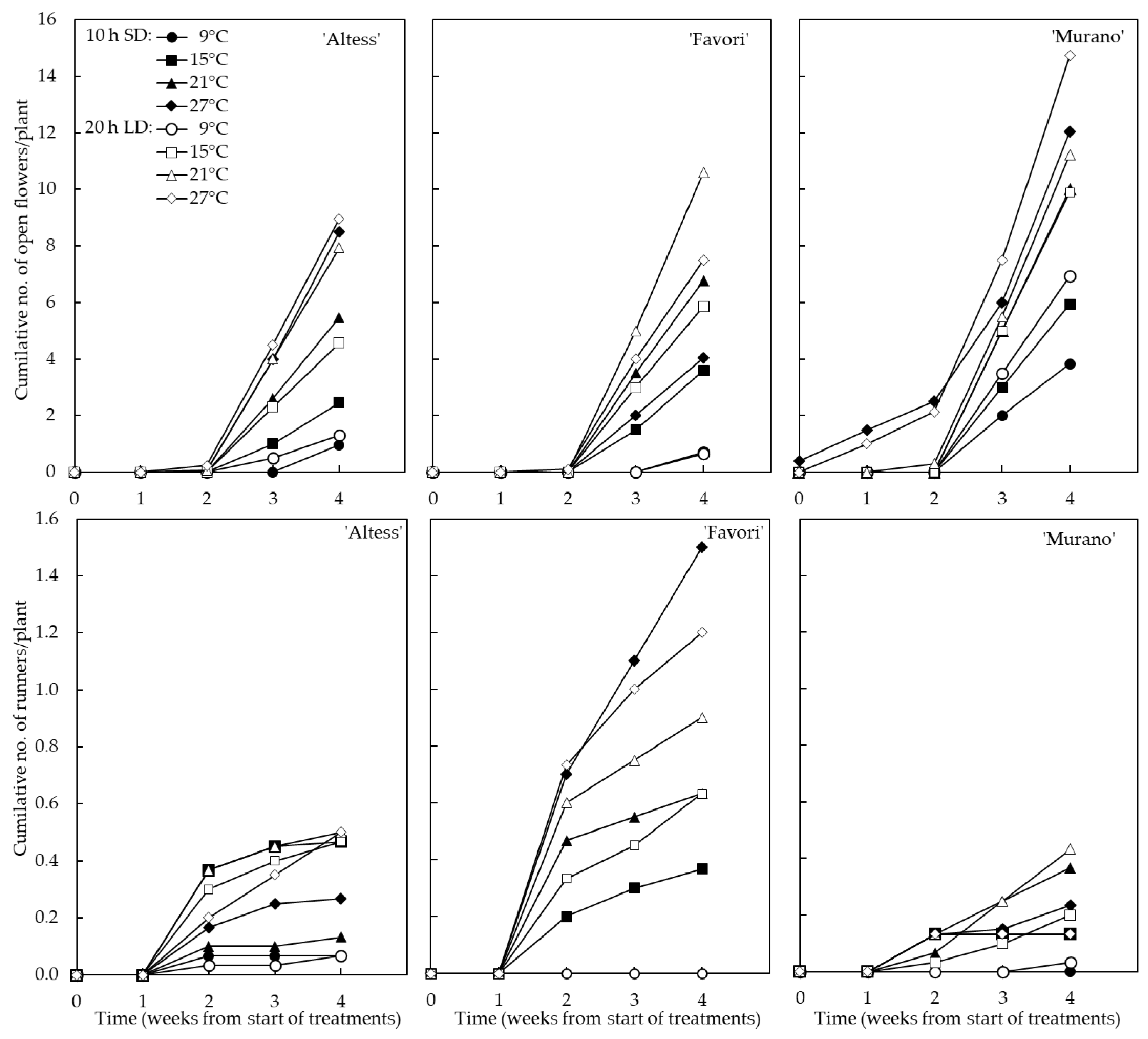
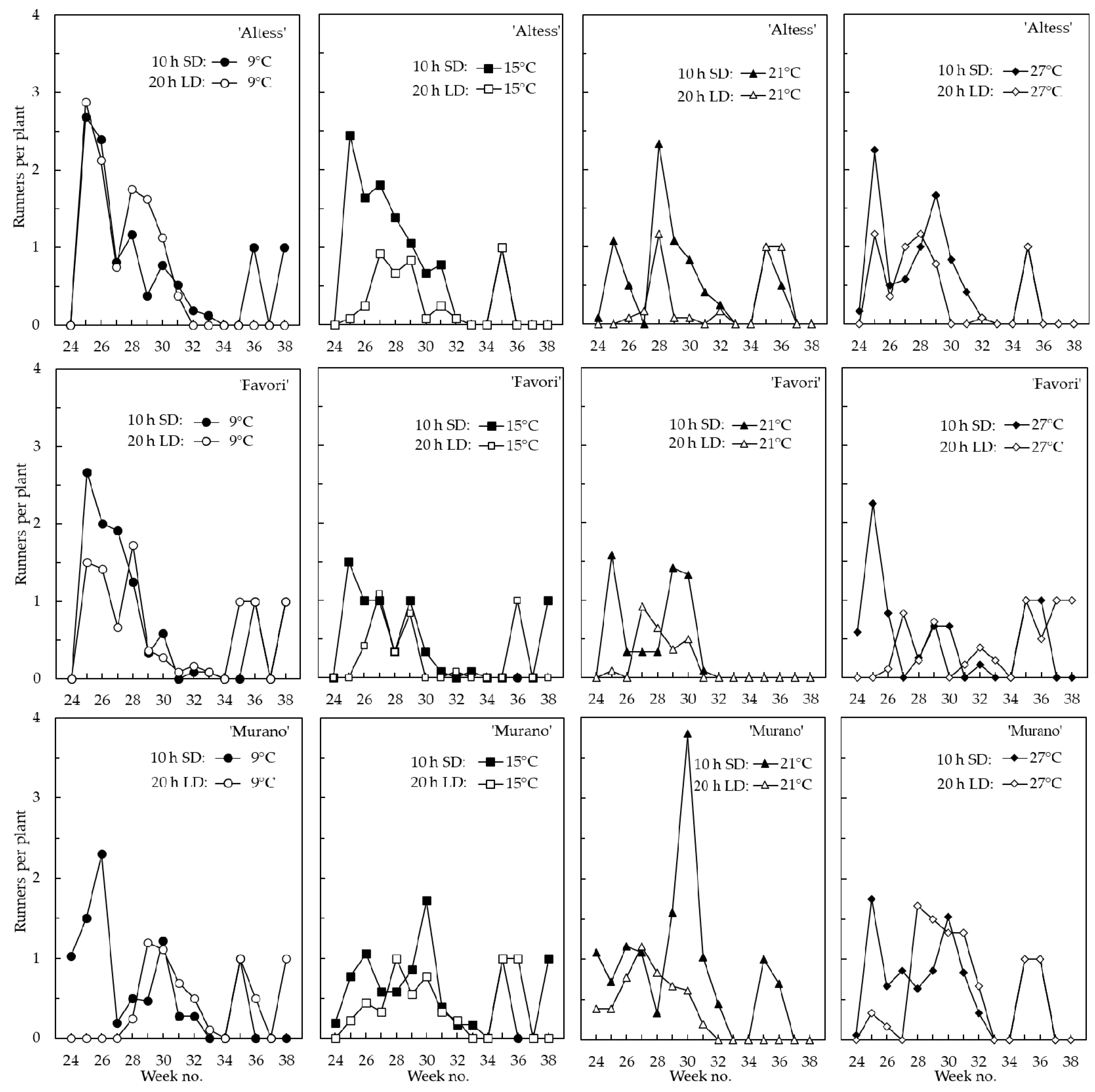
3.1. Flowering and Runnering in the Phytotron
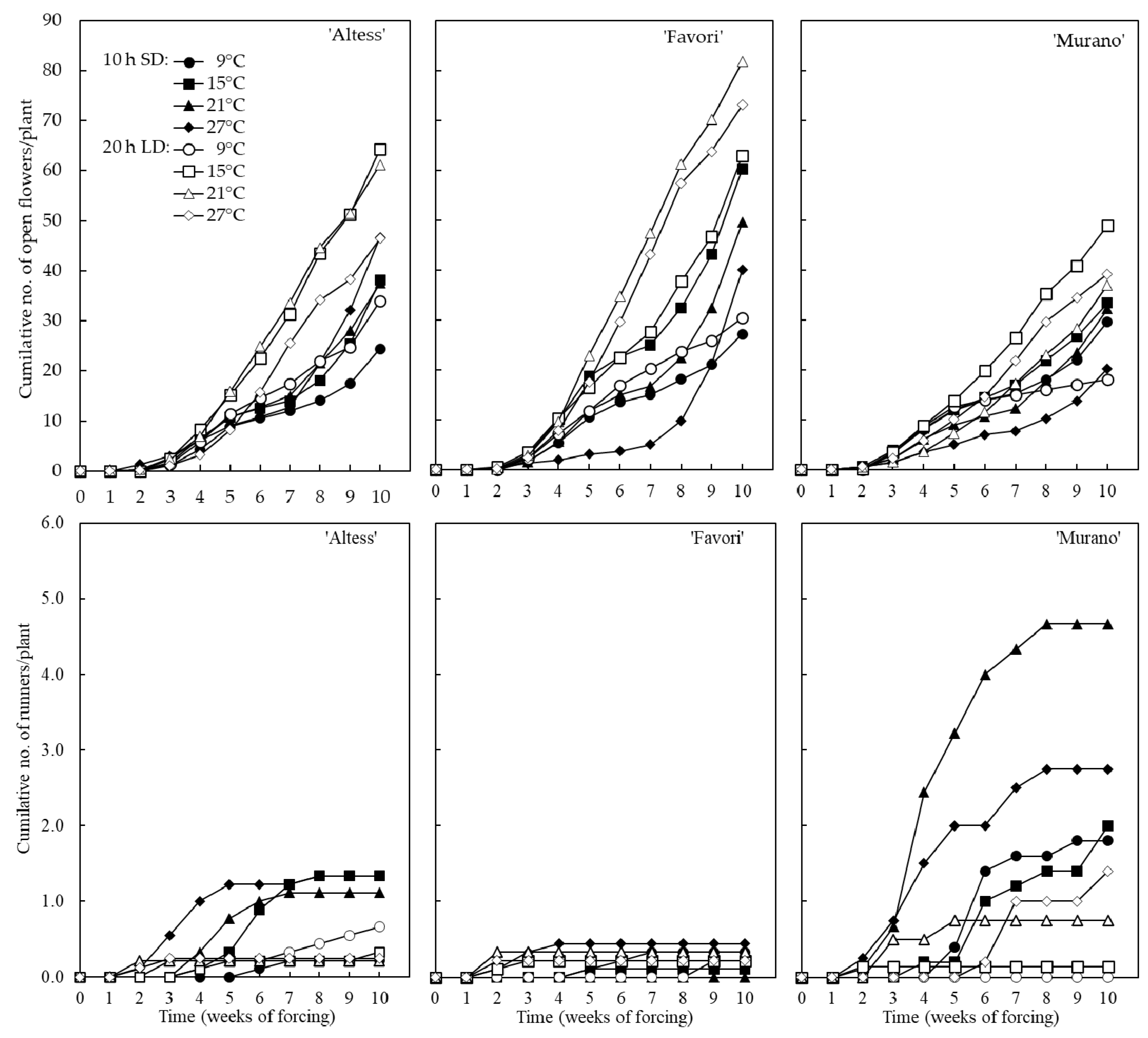
3.2. Flowering Potential of the Preconditioned Plants
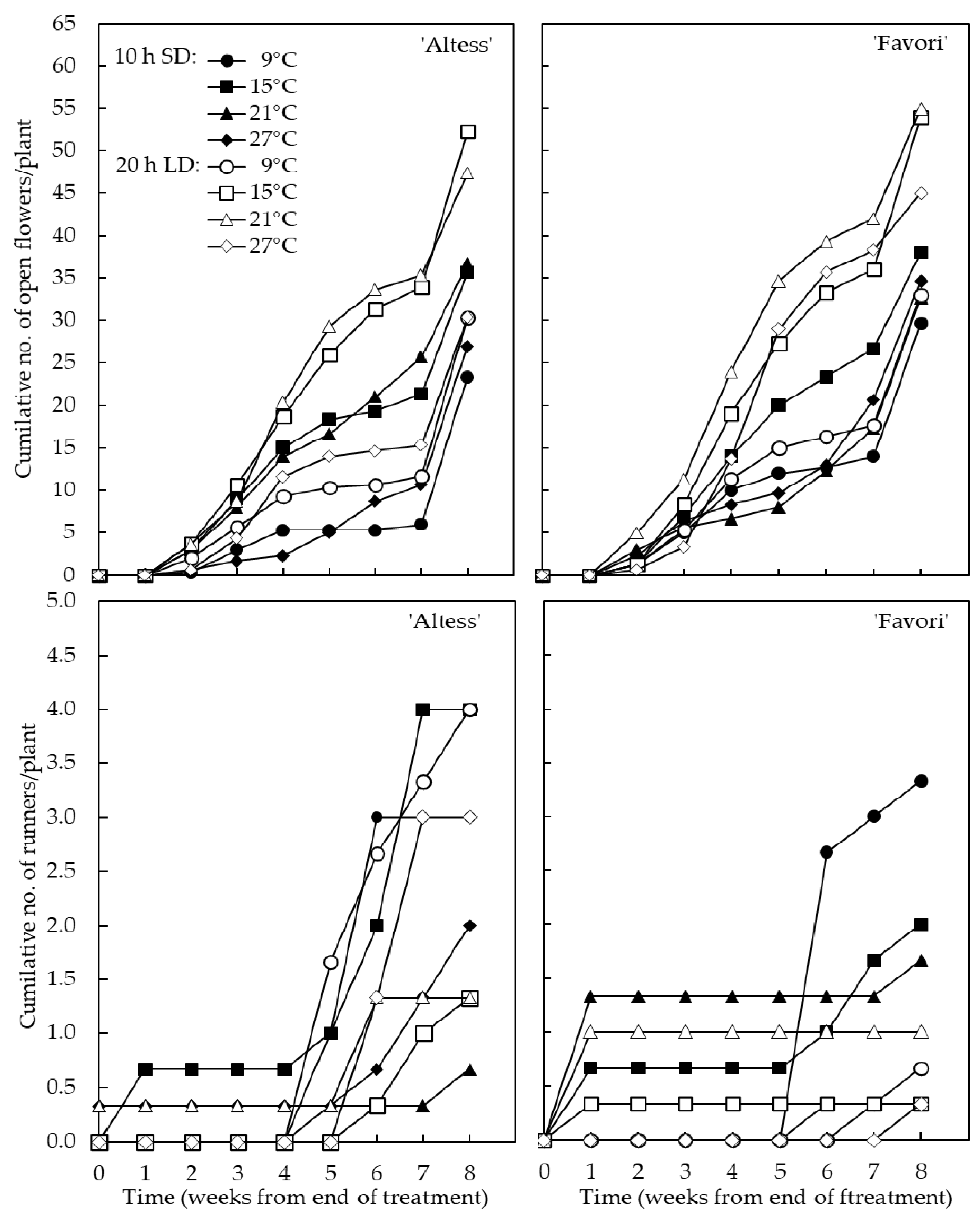
3.3. Plant Flowering Potential in Late Autumn
3.4. Yield Performance of the Preconditioned Plants
4. Discussion
4.1. Flowering and Runnering in the Phytotron
4.2. Flowering Potential of the Preconditioned Plants
4.3. Plant Flowering Potential in Late Autumn
4.4. Yield Performance of the Preconditioned Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flowers Plant−1 | Runners Plant−1 | ||||
|---|---|---|---|---|---|
| Temperature | Photoperiod (h) | ||||
| Cultivar | (°C) | 10 | 20 | 10 | 20 |
| Altess | 9 | 1.0 ± 0.4 | 1.3 ± 0.4 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| 15 | 2.5 ± 0.8 | 4.6 ± 0.4 | 0.5 ± 0.2 | 0.5 ± 0.1 | |
| 21 | 5.5 ± 2.4 | 8.0 ± 1.0 | 0.1 ± 0.0 | 0.5 ± 0.2 | |
| 27 | 8.6 ± 1.3 | 8.9 ± 2.8 | 0.3 ± 0.1 | 0.5 ± 0.1 | |
| Mean | 4.4 | 5.7 | 0.2 | 0.4 | |
| Favori | 9 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.0 ± | 0.0 ± 0.0 |
| 15 | 3.6 ± 0.0 | 5.9 ± 1.2 | 0.4 ± | 0.6 ± 0.1 | |
| 21 | 6.8 ± 0.9 | 10.6 ± 1.4 | 0.6 ± | 0.9 ± 0.3 | |
| 27 | 4.0 ± 0.9 | 7.6 ± 1.8 | 1.5 ± | 1.2 ± 0.2 | |
| Mean | 3.8 | 6.2 | 0.6 | 0.7 | |
| Murano | 9 | 3.8 ± 0.9 | 7.1 ± 1.5 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 15 | 5.9 ± 1.7 | 10.0 ± 2.5 | 0.1 ± 0.0 | 0.2 ± 0.0 | |
| 21 | 10.0 ± 3.2 | 11.7 ± 1.3 | 0.4 ± 0.0 | 0.4 ± 0.1 | |
| 27 | 13.6 ± 5.4 | 16.9 ± 1.4 | 0.2 ± 0.0 | 0.1 ± 0.0 | |
| Mean | 8.3 | 11.4 | 0.2 | 0.2 | |
| Probability level of significance (ANOVA) | |||||
| Source of variation | |||||
| Temperature (A) | <0.001 | <0.001 | |||
| Photoperiod (B) | 0.003 | ns | |||
| A × B | ns | ns | |||
| Cultivar (C) | <0.001 | <0.001 | |||
| C × A | 0.002 | <0.001 | |||
| C × B | ns | ns | |||
| A × B × C | ns | ns | |||
| After Preconditioning | In Late Autumn | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Flowers Plant−1 | Runners Plant−1 | Flowers Plant−1 | Runners Plant−1 | |||||||
| Temperature | Photoperiod (h) | |||||||||
| Cultivar | (°C) | 10 | 20 | 10 | 20 | 10 | 20 | 10 | 20 | |
| Altess | 9 | 42.9 ± 6.9 | 56.9 ± 13.0 | 0.3 ± 0.0 | 0.7 ± 0.3 | 23.3 ± 6.0 | 30.3 ± 4.0 | 3.0 ± 1.0 | 4.0 ± 1.0 | |
| 15 | 63.0 ± 8.3 | 87.8 ± 23.0 | 1.3 ± 0.2 | 0.5 ± 0.0 | 35.7 ± 9.1 | 52.3 ± 14.0 | 4.0 ± 0.0 | 2.0 ± 0.0 | ||
| 21 | 58.8 ± 1.9 | 83.3 ± 23.0 | 1.1 ± 0.6 | 0.7 ± 0.0 | 36.7 ± 7.2 | 47.3 ± 8.5 | 1.0 ± 0.0 | 2.0 ± 0.4 | ||
| 27 | 75.2 ± 14.5 | 67.3 ± 21.2 | 1.3 ± 0.6 | 0.4 ± 0.1 | 27.0 ± 3.5 | 30.3 ± 7.2 | 2.0 ± 0.8 | 3.0 ± 1.7 | ||
| Mean | 60.0 | 73.8 | 1.1 | 0.6 | 30.7 | 40.1 | 2.6 | 2.9 | ||
| Favori | 9 | 49.9 ± 8.2 | 54.9 ± 6.9 | 0.5 ± 0.1 | 0.3 ± 0.0 | 29.7 ± 2.3 | 33.0 ± 7.2 | 3.3 ± 1.1 | 2.0 ± 0.0 | |
| 15 | 91.1 ± 3.7 | 99.9 ± 14.6 | 0.3 ± 0.0 | 0.3 ± 0.0 | 38.0 ± 7.0 | 54.0 ± 4.4 | 2.0 ± 0.1 | 1.0 ± 0.0 | ||
| 21 | 81.7 ± 5.5 | 109.2 ± 27.1 | 0.0 ± 0.0 | 0.5 ± 0.0 | 32.7 ± 4.9 | 55.0 ± 7.9 | 1.7 ± 0.2 | 1.5 ± 0.7 | ||
| 27 | 79.8 ± 16.6 | 92.8 ± 5.4 | 0.4 ± 0.1 | 0.7 ± 0.0 | 34.7 ± 7.2 | 45.0 ± 7.0 | 1.0 ± 0.0 | 1.0 ± 0.0 | ||
| Mean | 75.6 | 89.2 | 0.4 | 0.4 | 33.8 | 46.8 | 2.2 | 1.4 | ||
| Murano | 9 | 43.5 ± 13.7 | 19.0 ± 8.5 | 2.3 ± 1.0 | 0.0 ± 0.0 | - | - | - | - | |
| 15 | 47.6 ± 1.9 | 59.7 ± 22.7 | 2.7 ± 0.5 | 0.3 ± 0.0 | - | - | - | - | ||
| 21 | 47.0 ± 13.1 | 52.7 ± 18.3 | 4.7 ± 2.4 | 3.0 ± 0.0 | - | - | - | - | ||
| 27 | 33.8 ± 6.8 | 44.8 ± 21.9 | 3.8 ± 1.1 | 1.3 ± 0.5 | - | - | - | - | ||
| Mean | 43.8 | 46.3 | 3.5 | 1.5 | - | - | - | - | ||
| Probability level of significance (ANOVA) | ||||||||||
| Source of variation | ||||||||||
| Temperature (A) | 0.008 | ns | 0.004 | 0.041 | ||||||
| Photoperiod (B) | 0.03 | <0.001 | 0.001 | ns | ||||||
| A × B | ns | ns | ns | 0.030 | ||||||
| Cultivar (C) | <0.001 | <0.001 | ns | ns | ||||||
| C × A | ns | 0.04 | ns | ns | ||||||
| C × B | ns | <0.001 | ns | ns | ||||||
| A × B × C | ns | ns | ns | ns | ||||||
References
- Heide, O.M.; Stavang, J.A.; Sønsteby, A. Physiology and genetics of flowering in cultivated and wild strawberries-a review. J. Hortic. Sci. Biotec. 2013, 88, 1–18. [Google Scholar] [CrossRef]
- Rivero, R.; Remberg, S.F.; Heide, O.M.; Sønsteby, A. Environmental Regulation of Dormancy, Flowering and Runnering in Two Genetically Distant Everbearing Strawberry Cultivars. Sci. Hortic. 2021, 290, 110515. [Google Scholar] [CrossRef]
- Lieten, P. The strawberry nursery industry in The Netherlands: An update. Acta Hortic. 2014, 1049, 99–106. [Google Scholar] [CrossRef]
- Sønsteby, A.; Sadojevic, M.; Heide, O.M. Effect of rooting date and environmental conditions during plant raising on fruit yield performance of two everbearing strawberry cultivars in a Nordic climate. Horticulturae 2022, 8, 249. [Google Scholar] [CrossRef]
- Gallace, N.; Lieten, P.; Boonen, M.; Bylemans, D. Reduced winter chill as a means to improve the reproductive potential of late day-neutral strawberry cultivars. Eur. J. Hortic. Sci. 2019, 84, 20–23. [Google Scholar] [CrossRef]
- Melis, P. Manipulation of Everbearers to Steer Harvest Peaks and Yielding. International Soft Fruit Conference’s-Hertogenbosch, Holland. 2020. Available online: http://goodberry-eu.eu/fileadmin/goodberry/appliedpublications/3737 (accessed on 10 November 2021).
- Sønsteby, A.; Woznicki, T.L.; Heide, O.M. Effects of runner removal and partial defoliation on growth and yield performance of ‘Favori’ everbearing strawberry plants. Horticulturae 2021, 7, 215. [Google Scholar]
- Sønsteby, A.; Heide, O.M. Long-day control of flowering in everbearing strawberries. J. Hortic. Sci. Biotec. 2007, 82, 875–884. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O.M. Quantitative long-day flowering response in the perpetual-flowering F1 strawberry cultivar Elan. J. Hortic. Sci. Biotec. 2007, 82, 266–274. [Google Scholar] [CrossRef]
- Samad, S.; Butare, D.; Marttila, S.; Sønsteby, A.; Khalil, S. Effects of Temperature and Photoperiod on the Flower Potential in Everbearing Strawberry as Evaluated by Meristem Dissection. Horticulturae 2021, 7, 484. [Google Scholar] [CrossRef]
- Gallace, N.; Lieten, P. Winter manipulation of photoperiod and chill to enhance the perpetual flowering nature of ‘Soprano’. Acta Hortic. 2021, 1309, 333–340. [Google Scholar] [CrossRef]
- Rivero, R.; Sønsteby, A.; Solhaug, K.A.; Heide, O.M.; Remberg, S.F. Effects of temperature and photoperiod on photosynthesis in everbearing strawberry. Acta Hortic. 2021, 1309, 379–386. [Google Scholar] [CrossRef]

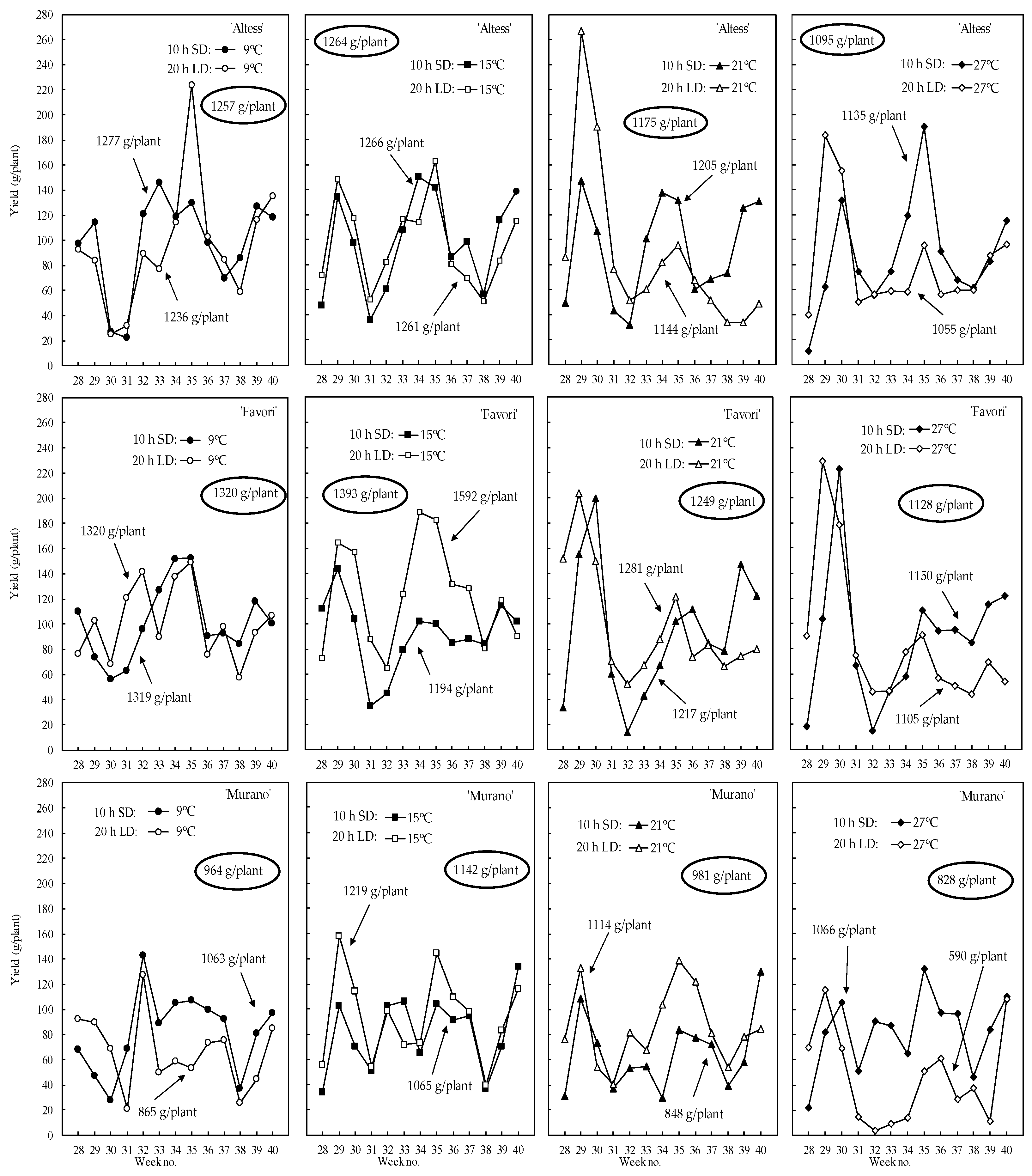
| Yield (g/Plant) | Berry Weight (g) | Berries Plant−1 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Temp. (°C) | Photoperiod (h) | |||||||
| 10 | 20 | 10 | 20 | 10 | 20 | ||||
| Altess | 9 | 1276.5 ± 147.4 | 1235.6 ± 82.6 | 20.2 ± 0.6 | 18.2 ± 1.0 | 63.3 ± 7.5 | 67.9 ± 5.1 | ||
| 15 | 1266.2 ± 211.2 | 1261.3 ± 53.9 | 18.2 ± 1.3 | 16.8 ± 0.6 | 70.2 ± 16.2 | 75.1 ± 5.1 | |||
| 21 | 1204.9 ± 98.1 | 1143.7 ± 331.7 | 18.6 ± 0.4 | 15.9 ± 0.7 | 64.8 ± 5.3 | 71.8 ± 20.1 | |||
| 27 | 1134.7 ± 204.8 | 1054.7 ± 352.1 | 17.6 ± 0.7 | 15.6 ± 0.4 | 64.4 ± 12.3 | 67.6 ± 22.9 | |||
| Mean | 1220.6 | 1173.8 | 18.7 | 16.6 | 65.7 | 70.6 | |||
| Favori | 9 | 1318.5 ± 12.9 | 1320.1 ± 20.5 | 18.0 ± 1.2 | 18.3 ± 0.3 | 73.3 ± 4.6 | 72.3 ± 0.8 | ||
| 15 | 1193.9 ± 99.3 | 1592.4 ± 203.0 | 18.1 ± 0.7 | 17.0 ± 1.1 | 66.0 ± 7.9 | 93.4 ± 6.6 | |||
| 21 | 1216.6 ± 124.7 | 1280.6 ± 263.8 | 17.8 ± 0.9 | 15.6 ± 0.3 | 68.6 ± 8.8 | 82.3 ± 17.8 | |||
| 27 | 1149.6 ± 234.1 | 1105.1 ± 439.2 | 17.1 ± 0.9 | 13.5 ± 0.3 | 67.8 ± 17.4 | 81.5 ± 32.3 | |||
| Mean | 1219.6 | 1324.5 | 17.8 | 16.1 | 68.9 | 82.4 | |||
| Murano | 9 | 1063.0 ± 177.3 | 864.5 ± 634.2 | 19.0 ± 1.9 | 14.9 ± 3.2 | 56.4 ± 12.7 | 54.0 ± 30.8 | ||
| 15 | 1065.0 ± 296.6 | 1219.1 ± 107.4 | 19.0 ± 0.4 | 17.6 ± 0.6 | 56.1 ± 16.9 | 69.2 ± 5.9 | |||
| 21 | 848.2 ± 247.7 | 1114.1 ± 192.5 | 17.0 ± 1.3 | 17.4 ± 1.0 | 50.4 ± 16.3 | 64.0 ± 8.2 | |||
| 27 | 1065.7 ± 180.5 | 589.5 ± 116.4 | 18.7 ± 0.6 | 14.4 ± 1.2 | 57.1 ± 11.0 | 40.8 ± 6.9 | |||
| Mean | 1010.0 | 946.8 | 18.5 | 16.1 | 55.0 | 57.0 | |||
| Probability level of significance (ANOVA) | |||||||||
| Source of variation | |||||||||
| Temperature (A) | <0.001 | ns | ns | ||||||
| Photoperiod (B) | <0.001 | <0.001 | <0.001 | ||||||
| A × B | 0.03 | <0.001 | ns | ||||||
| Cultivar (C) | <0.001 | <0.001 | 0.001 | ||||||
| C × A | ns | ns | 0.001 | ||||||
| C × B | ns | ns | ns | ||||||
| A × B × C | ns | ns | 0.04 | ||||||
| Berries >25 mm (%) | Unsaleable Berries (g/plant) | ||||||||
| Cultivar | Temperature (°C) | Photoperiod (h) | |||||||
| 10 | 20 | 10 | 20 | ||||||
| Altess | 9 | 99.6 ± 0.2 | 98.6 ± 1.0 | 19.4 ± 2.9 | 32.4 ± 3.6 | ||||
| 15 | 99.0 ± 0.4 | 98.6 ± 0.4 | 19.3 ± 3.7 | 20.8 ± 4.8 | |||||
| 21 | 99.2 ± 0.2 | 98.3 ± 0.6 | 12.3 ± 4.8 | 18.0 ± 4.9 | |||||
| 27 | 98.1 ± 0.5 | 98.0 ± 0.4 | 11.6 ± 3.7 | 14.1 ± 5.7 | |||||
| Mean | 99.0 | 98.4 | 15.6 | 21.3 | |||||
| Favori | 9 | 99.0 ± 0.4 | 98.6 ± 0.6 | 17.1 ± 3.5 | 8.8 ± 2.0 | ||||
| 15 | 99.0 ± 0.9 | 98.1 ± 0.8 | 14.3 ± 3.9 | 9.4 ± 2.9 | |||||
| 21 | 98.2 ± 0.9 | 97.0 ± 0.4 | 25.4 ± 8.6 | 8.4 ± 2.7 | |||||
| 27 | 97.6 ± 0.3 | 94.8 ± 0.8 | 16.8 ± 2.1 | 22.4 ± 3.1 | |||||
| Mean | 98.5 | 97.1 | 18.4 | 12.3 | |||||
| Murano | 9 | 98.2 ± 0.6 | 94.4 ± 5.8 | 28.2 ± 9.2 | 21.0 ± 2.7 | ||||
| 15 | 98.8 ± 0.3 | 97.2 ± 0.9 | 25.8 ± 11.1 | 19.9 ± 9.9 | |||||
| 21 | 96.6 ± 1.4 | 96.5 ± 0.7 | 8.9 ± 3.1 | 15.0 ± 2.3 | |||||
| 27 | 97.1 ± 1.8 | 95.4 ± 0.8 | 9.9 ± 5.7 | 2.3 ± 1.1 | |||||
| Mean | 97.7 | 95.9 | 18.2 | 12.6 | |||||
| Probability level of significance (ANOVA) | |||||||||
| Source of variation | |||||||||
| Temperature (A) | ns | ns | |||||||
| Photoperiod (B) | 0.005 | 0.003 | |||||||
| A × B | 0.01 | ns | |||||||
| Cultivar (C) | <0.001 | 0.007 | |||||||
| C × A | ns | ns | |||||||
| C × B | ns | ns | |||||||
| A × B × C | ns | ns | |||||||
| Plant Height (cm) | Crowns Plant−1 | Leaves Plant−1 | Runners Plant−1 * | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Temp. | Photoperiod (h) | |||||||||||||
| (°C) | 10 | 20 | 10 | 20 | 10 | 20 | 10 | 20 | |||||||
| Altess | 9 | 28.0 ± 2.3 | 28.5 ± 0.7 | 4.0 ± 1.0 | 4.1 ± 0.8 | 31.4 ± 6.3 | 37.0 ± 2.8 | 5.6 ± 1.3 | 7.8 ± 1.7 | ||||||
| 15 | 28.0 ± 1.4 | 25.9 ± 2.8 | 4.5 ± 1.2 | 4.4 ± 0.8 | 32.8 ± 8.8 | 35.8 ± 0.5 | 8.4 ± 1.5 | 2.9 ± 0.4 | |||||||
| 21 | 27.0 ± 1.8 | 23.9 ± 2.1 | 3.9 ± 0.1 | 3.0 ± 0.6 | 35.4 ± 4.2 | 24.4 ± 6.2 | 7.1 ± 1.8 | 2.1 ± 0.7 | |||||||
| 27 | 27.1 ± 4.1 | 23.1 ± 3.7 | 4.6 ± 0.1 | 2.9 ± 1.2 | 32.5 ± 1.4 | 26.7 ± 8.1 | 6.6 ± 1.3 | 4.4 ± 1.8 | |||||||
| Mean | 27.5 | 25.4 | 4.3 | 3.6 | 33.2 | 30.9 | 7.0 | 4.3 | |||||||
| Favori | 9 | 29.8 ± 3.4 | 29.8 ± 0.6 | 5.9 ± 0.8 | 3.8 ± 0.4 | 38.9 ± 7.2 | 35.8 ± 1.9 | 6.9 ± 1.8 | 4.6 ± 1.9 | ||||||
| 15 | 33.2 ± 2.7 | 33.2 ± 3.2 | 5.3 ± 1.8 | 6.3 ± 0.7 | 41.6 ± 10.6 | 51.8 ± 6.6 | 4.3 ± 2.2 | 2.3 ± 1.7 | |||||||
| 21 | 32.8 ± 3.3 | 26.0 ± 1.1 | 5.5 ± 0.3 | 5.1 ± 0.5 | 57.8 ± 2.6 | 41.7 ± 6.4 | 5.1 ± 0.4 | 2.5 ± 1.1 | |||||||
| 27 | 31.3 ± 3.6 | 22.9 ± 2.9 | 7.3 ± 0.8 | 4.3 ± 1.7 | 55.3 ± 7.7 | 31.2 ± 9.3 | 4.6 ± 1.5 | 2.6 ± 1.3 | |||||||
| Mean | 31.8 | 28.0 | 6.0 | 4.8 | 48.4 | 40.1 | 5.2 | 3.0 | |||||||
| Murano | 9 | 23.1 ± 2.3 | 19.6 ± 3.8 | 4.8 ± 1.4 | 4.2 ± 2.0 | 30.6 ± 4.5 | 35.4 ± 17.4 | 5.5 ± 0.9 | 5.1 ± 1.1 | ||||||
| 15 | 24.4 ± 2.5 | 23.6 ± 3.2 | 4.8 ± 1.6 | 4.0 ± 1.3 | 37.3 ± 8.6 | 28.3 ± 8.5 | 5.4 ± 0.5 | 3.4 ± 1.1 | |||||||
| 21 | 26.9 ± 2.7 | 21.8 ± 1.9 | 6.3 ± 1.4 | 3.6 ± 0.5 | 44.1 ± 9.4 | 26.1 ± 7.1 | 10.1 ± 3.5 | 4.3 ± 1.5 | |||||||
| 27 | 23.6 ± 2.5 | 17.2 ± 5.8 | 7.3 ± 1.4 | 5.2 ± 2.0 | 43.9 ± 19.3 | 33.3 ± 17.4 | 6.9 ± 2.3 | 6.8 ± 3.6 | |||||||
| Mean | 24.5 | 20.5 | 5.8 | 4.2 | 39.0 | 30.8 | 7.0 | 4.9 | |||||||
| Probability level of significance (ANOVA) | |||||||||||||||
| Source of variation | |||||||||||||||
| Temperature (A) | <0.010 | ns | ns | ns | |||||||||||
| Photoperiod (B) | <0.001 | 0.042 | 0.038 | <0.001 | |||||||||||
| A × B | 0.001 | ns | 0.006 | 0.019 | |||||||||||
| Cultivar(C) | 0.022 | 0.044 | 0.010 | 0.008 | |||||||||||
| C × A | ns | ns | ns | ns | |||||||||||
| C × B | ns | ns | ns | ns | |||||||||||
| A × B × C | ns | ns | ns | ns | |||||||||||
| Plant FW (g) | Infloresc. Plant−1 * | Flowers/Fruits not Harvested | |||||||||||||
| Cultivar | Temp. | Photoperiod (h) | |||||||||||||
| (°C) | 10 | 20 | 10 | 20 | 10 | 20 | |||||||||
| Altess | 9 | 287.1 ± 21.1 | 295.2 ± 54.0 | 12.2 ± 0.1 | 12.2 ± 2.6 | 70.7 ± 11.3 | 78.3 ± 14.5 | ||||||||
| 15 | 267.6 ± 30.0 | 211.3 ± 5.5 | 12.8 ± 1.6 | 14.5 ± 0.7 | 85.1 ± 14.3 | 62.9 ± 6.0 | |||||||||
| 21 | 277.0 ± 23.9 | 146.6 ± 43.1 | 14.3 ± 0.6 | 11.3 ± 2.3 | 81.8 ± 14.6 | 61.3 ± 13.9 | |||||||||
| 27 | 261.2 ± 15.6 | 158.1 ± 56.7 | 13.4 ± 0.8 | 11.3 ± 1.7 | 74.3 ± 12.3 | 61.5 ± 17.0 | |||||||||
| Mean | 271.9 | 202.8 | 13.3 | 12.3 | 78.6 | 66.0 | |||||||||
| Favori | 9 | 297.3 ± 61.4 | 262.3 ± 16.3 | 13.6 ± 3.1 | 11.3 ± 0.7 | 63.5 ± 20.2 | 52.8 ± 13.7 | ||||||||
| 15 | 320.6 ± 66.3 | 341.9 ± 62.2 | 16.3 ± 3.4 | 17.4 ± 2.4 | 88.6 ± 43.8 | 71.8 ± 26.5 | |||||||||
| 21 | 411.9 ± 34.8 | 231.6 ± 35.8 | 19.4 ± 1.3 | 18.6 ± 4.5 | 117.7 ± 6.5 | 76.6 ± 10.4 | |||||||||
| 27 | 385.2 ± 56.0 | 179.7 ± 42.2 | 18.3 ± 3.4 | 13.7 ± 1.5 | 112.3 ± 31.7 | 66.1 ± 5.5 | |||||||||
| Mean | 353.8 | 253.9 | 16.9 | 15.2 | 95.5 | 66.8 | |||||||||
| Murano | 9 | 185.0 ± 14.8 | 157.3 ± 94.2 | 12.6 ± 1.9 | 9.5 ± 3.5 | 76.3 ± 17.0 | 66.0 ± 24.1 | ||||||||
| 15 | 228.6 ± 41.5 | 157.8 ± 57.5 | 14.6 ± 3.9 | 11.8 ± 1.7 | 114.2 ± 15.9 | 67.7 ± 14.9 | |||||||||
| 21 | 242.8 ± 46.6 | 161.9 ± 35.4 | 17.6 ± 1.7 | 12.7 ± 2.7 | 128.2 ± 18.1 | 59.3 ± 10.4 | |||||||||
| 27 | 211.3 ± 30.1 | 163.5 ± 68.3 | 14.6 ± 6.2 | 15.2 ± 6.3 | 98.7 ± 41.0 | 112.5 ± 53.5 | |||||||||
| Mean | 216.9 | 160.1 | 14.8 | 12.3 | 104.3 | 76.4 | |||||||||
| Probability level of significance (ANOVA) | |||||||||||||||
| Source of variation | |||||||||||||||
| Temperature (A) | ns | 0.019 | 0.042 | ||||||||||||
| Photoperiod (B) | <0.001 | ns | 0.022 | ||||||||||||
| A × B | 0.002 | ns | ns | ||||||||||||
| Cultivar(C) | <0.001 | 0.044 | ns | ||||||||||||
| C × A | ns | ns | ns | ||||||||||||
| C × B | ns | ns | ns | ||||||||||||
| A × B × C | ns | ns | ns | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivero, R.; Remberg, S.F.; Heide, O.M.; Sønsteby, A. Effect of Temperature and Photoperiod Preconditioning on Flowering and Yield Performance of Three Everbearing Strawberry Cultivars. Horticulturae 2022, 8, 504. https://doi.org/10.3390/horticulturae8060504
Rivero R, Remberg SF, Heide OM, Sønsteby A. Effect of Temperature and Photoperiod Preconditioning on Flowering and Yield Performance of Three Everbearing Strawberry Cultivars. Horticulturae. 2022; 8(6):504. https://doi.org/10.3390/horticulturae8060504
Chicago/Turabian StyleRivero, Rodmar, Siv Fagertun Remberg, Ola M. Heide, and Anita Sønsteby. 2022. "Effect of Temperature and Photoperiod Preconditioning on Flowering and Yield Performance of Three Everbearing Strawberry Cultivars" Horticulturae 8, no. 6: 504. https://doi.org/10.3390/horticulturae8060504
APA StyleRivero, R., Remberg, S. F., Heide, O. M., & Sønsteby, A. (2022). Effect of Temperature and Photoperiod Preconditioning on Flowering and Yield Performance of Three Everbearing Strawberry Cultivars. Horticulturae, 8(6), 504. https://doi.org/10.3390/horticulturae8060504