
Successional Effects of No-Till Cover Cropping with Black Oat (Avena strigosa) vs. Soil Solarization on Soil Health in a Tropical Oxisol



Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Trials
2.2. Soil Sample Analysis
2.3. Nematode Assays
2.4. Soil Water and Other Physical Properties and Temperature Monitoring
2.5. Corn Growth and Yield
2.6. Statistical Analysis
3. Results
3.1. Pre-Plant Conditions
3.2. Effects of No-Till Cropping and Black Oat Cover Cropping on Soil Properties
3.3. Effects on Nematode Community as Soil Health Indicators
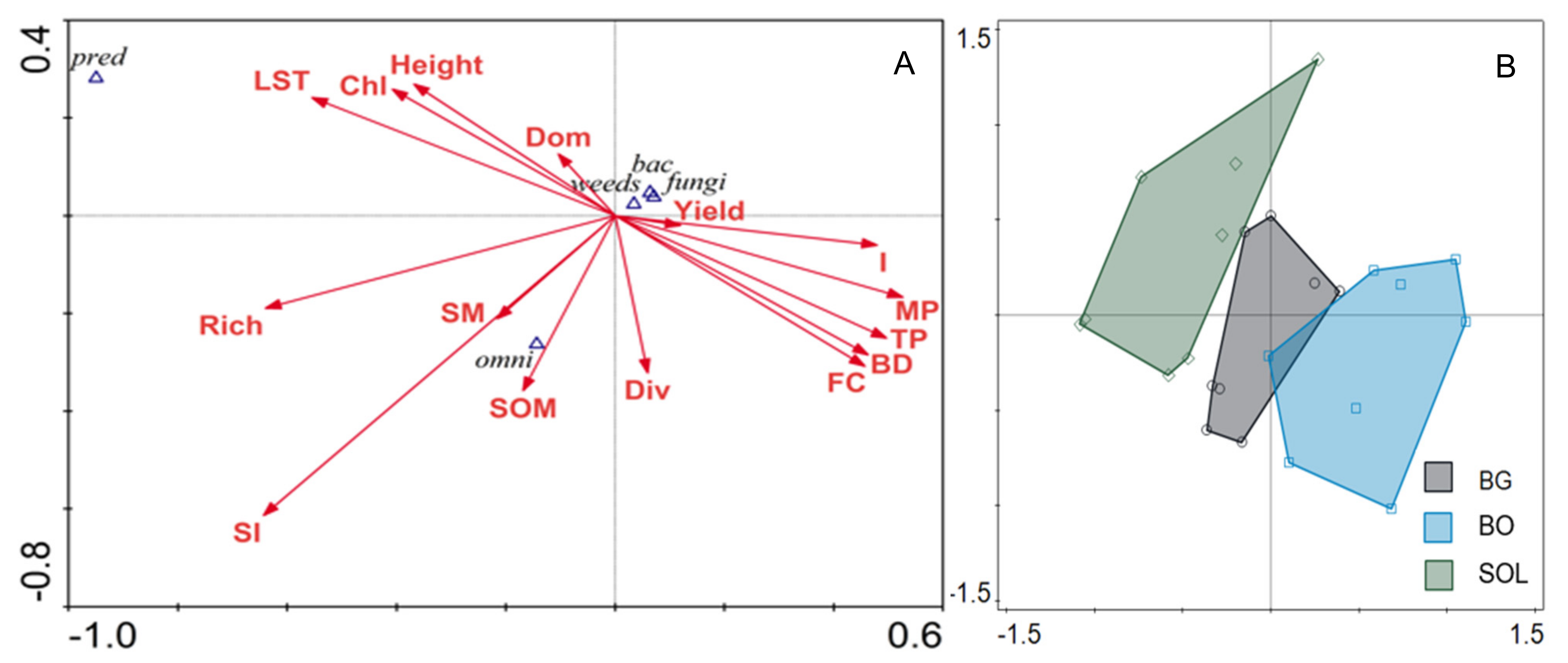
3.4. Relationships between Soil Properties and Free-Living Nematode Abundance
4. Discussion
4.1. Effects of No-Till Black Oat and Soil Solarization on Different Soil Properties
4.1.1. Water-Holding Capacity
4.1.2. Infiltration and Macroporosity
4.1.3. Bulk Density
4.2. Effects of No-Till Black Oat Cropping and Solarization on Nematode Communities
4.2.1. Soil Food Web Structure
4.2.2. Plant-Parasitic Nematodes
4.3. Relationship between Soil Food Web Structure and Soil Water Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Triplett, G.B.; Dick, W. Continuous Application of No-Tillage to Ohio Soils. Agron. J. 2008, 100, S153–S165. [Google Scholar] [CrossRef]
- Lal, R. The Plow and Agricultural Sustainability. J. Sustain. Agric. 2009, 33, 66–84. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Shaxson, F.; Pretty, J. The Spread of Conservation Agriculture: Justification, Sustainability and Uptake. Int. J. Agric. Sustain. 2009, 7, 292–320. [Google Scholar] [CrossRef]
- Holland, J.M. The Environmental Consequences of Adopting Conservation Tillage in Europe: Reviewing the Evidence. Agric. Ecosyst. Environ. 2004, 103, 1–25. [Google Scholar] [CrossRef]
- Greenland, D.J.; Rimmer, D.; Payne, D. Determination of the Structural Stability Class of English and Welsh Soils, Using a Water Coherence Test. J. Soil Sci. 1975, 26, 294–303. [Google Scholar] [CrossRef]
- Evans, R. Some Soil Factors Influencing Accelerated Water Erosion of Arable Land. Prog. Phys. Geogr. 1996, 20, 205–215. [Google Scholar] [CrossRef]
- Miguel Reichert, J.; Trevisan da Rosa, V.; Saldanha Vogelmann, E.; Peres da Rosa, D.; Horn, R.; José Reinert, D.; Sattler, A.; Eloir Denardin, J. Conceptual Framework for Capacity and Intensity Physical Soil Properties Affected by Short and Long-Term (14 Years) Continuous No-Tillage and Controlled Traffic. Soil Tillage Res. 2016, 158, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Brady, N.C.; Weil, R.R. Elements of the Nature and Properties of Soils, 3rd ed.; Pearson Prentice Hall: New York, NY, USA, 2010; ISBN 9780135014332. [Google Scholar]
- Zotarelli, L.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Six, J. Impact of Tillage and Crop Rotation on Light Fraction and Intra-Aggregate Soil Organic Matter in Two Oxisols. Soil Tillage Res. 2007, 95, 196–206. [Google Scholar] [CrossRef]
- Marquez, J.M.K. Evaluating Effects of No-Till Cover Cropping Systems on Indigenous Entomopathogenic Nematodes and Fungi. Ph.D. Thesis, University of Hawaiʻi at Manoa, Honolulu, HI, USA, 2017. [Google Scholar]
- Gaskell, M.; Smith, R.; Jackson, L.E.; Hartz, T.K. Soil Nitrogen Fertility Management. In Cover Cropping for Vegetable Production. A Grower’s Handbook; Smith, R., Bugg, R.L., Gaskell, M., Daugovish, O., Horn, M.V., Eds.; The Regents of the University of California Agriculture and Natural Resources: Richmond, CA, USA, 2011; pp. 37–40. [Google Scholar]
- Bacq-Labreuil, A.; Crawford, J.; Mooney, S.J.; Neal, A.L.; Ritz, K. Cover Crop Species Have Contrasting Influence upon Soil Structural Genesis and Microbial Community Phenotype. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Price, A.J.; Wayne Reeves, D.; Patterson, M.G. Evaluation of Weed Control Provided by Three Winter Cereals in Conservation-Tillage Soybean. Renew. Agric. Food Syst. 2006, 21, 159–164. [Google Scholar] [CrossRef]
- Bauer, P.J.; Reeves, D.W. A Comparison of Winter Cereal Species and Planting Dates as Residue Cover for Cotton Grown with Conservation Tillage. Crop Sci. 1999, 39, 1824–1830. [Google Scholar] [CrossRef]
- Marahatta, S.P.; Wang, K.-H.; Sipes, B.S.; Hooks, C.R.R. Effects of the Integration of Sunn Hemp and Soil Solarization on Plant-Parasitic and Free-Living Nematodes. J. Nematol. 2012, 44, 72. [Google Scholar]
- Katan, J.; Gamliel, A. Soil Solarisation: Theory and Practice; American Phytopathological Society: St. Paul, MN, USA, 2012. [Google Scholar]
- Wang, K.-H.; Mcsorley, R.; Kokalis-Burelle, N. Effects of Cover Cropping, Solarization, and Soil Fumigation on Nematode Communities Nematode Soil Ecology View Project. Plant Soil 2006, 286, 229–243. [Google Scholar] [CrossRef]
- Quintanilla-Tornel, M.A.; Wang, K.H.; Tavares, J.; Hooks, C.R.R. Effects of Mulching on above and below Ground Pests and Beneficials in a Green Onion Agroecosystem. Agric. Ecosyst. Environ. 2016, 224, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Mcsorley, R.; Wang, K.; Frederick, J.J. Integrated Effects of Solarization, Sunn Hemp Cover Crop, and Amendment on Nematodes, Weeds, and Pepper Yields. Nematropica 2008, 38, 115–125. [Google Scholar]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; Deyn, G.D.; Goede, R.D.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil Quality—A Critical Review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Moebius-Clune, B.N.; Moebius-Clune, D.J.; Gugino, B.K.; Idowu, O.J.; Schindelbeck, R.R.; Ristow, A.J.; van Es, H.M.; Thies, J.E.; Shayler, H.A.; McBride, M.B.; et al. Comprehensive Assessment of Soil Health—The Cornell Framework, 3.2 ed.; Cornell University: Geneva, NY, USA, 2016; ISBN 0967650763. [Google Scholar]
- Bongers, T.; Bongers, M. Functional Diversity of Nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera-an Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Forge, T.A.; Simard, S.W. Trophic Structure of Nematode Communities, Microbial Biomass, and Nitrogen Mineralization in Soils of Forests and Clearcuts in the Southern Interior of British Columbia. Can. J. Soil Sci. 2000, 80, 401–410. [Google Scholar] [CrossRef]
- Ferris, H.; Griffiths, B.S.; Porazinska, D.L.; Powers, T.O.; Wang, K.H.; Tenuta, M. Reflections on Plant and Soil Nematode Ecology: Past, Present and Future. J. Nematol. 2012, 44, 115–126. [Google Scholar]
- Wang, K.-H.; Sipes, B.S.; Hooks, C.R.R. Sunn Hemp Cover Cropping and Solarization as Alternatives to Soil Fumigants for Pineapple Production. Acta Hortic. 2011, 902, 221–232. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. A Framework for Soil Food Web Diagnostics: Extension of the Nematode Faunal Analysis Concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Ikawa, H.; Sato, H.H.; Chang, A.K.S.; Nakamura, S.; Robello, E.; Periaswamy, S.P.; Rep, B.S.P.T. Soils of the Hawaii Agricultural Experiment Station, University of Hawaii: Soil Survey, Laboratory Data, and Soil Descriptions; HITHAR Research Extension Series; RES- 022; University of Hawaii: Honolulu, HI, USA, 1985; pp. 1–76. [Google Scholar]
- Byrd, D.W.; Barker, K.R.; Ferris, H.; Nusbaum, C.J.; Griffin, W.E.; Small, R.H.; Stone, C.A. Two Semi-Automatic Elutriators for Extracting Nematodes and Certain Fungi from Soil. J. Nematol. 1976, 8, 206–212. [Google Scholar]
- Jenkins, W.R. A Rapid Centrifugal-Flotation Technique for Separating Nematodes from Soil. Plant Dis. Report. 1964, 48, 692. [Google Scholar]
- Okada, H.; Kadota, I. Host Status of 10 Fungal Isolates for Two Nematode Species, Filenchus misellus and Aphelenchus avenae. Soil Biol. Biochem. 2003, 35, 1601–1607. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Freckman, D.W.; Ettema, C.H. Assessing Nematode Communities in Agroecosystems of Varying Human Intervention. Agric. Ecosyst. Environ. 1993, 45, 239–261. [Google Scholar] [CrossRef]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Source Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Bouwer, H. Intake Rate: Cylinder Infiltrometer. Methods Soil Anal. Part 1 Phys. Mineral. Methods 2018, 5, 825–844. [Google Scholar] [CrossRef]
- Gantzer, C.J.; Blake, G.R. Physical Characteristics of Le Sueur Clay Loam Soil Following No-till and Conventional Tillage 1. Agron. J. 1978, 70, 853–857. [Google Scholar] [CrossRef]
- Vomocil, J.A. Porosity. Methods Soil Anal. Part 1 Phys. Mineral. Prop. Incl. Stat. Meas. Sampl. 2015, 9, 299–314. [Google Scholar] [CrossRef]
- Green, R.E.; Ahuja, L.R.; Chong, S.-K.; Lau, L.S. Water Conduction in Hawaii Oxic Soils; Water Resources Research Center, University of Hawaii at Manoa: Honolulu, HI, USA, 1982. [Google Scholar]
- Peters, D.B. Water Availability. Methods Soil Anal. Part 1 Phys. Mineral. Prop. Incl. Stat. Meas. Sampl. 2015, 279–285. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.; et al. Package ‘Vegan’, Community Ecology Package, Version 2. Available online: http://cran.r-project.org/package=vegan (accessed on 13 February 2022).
- Ter Braak, C.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 5.0)—Research@WUR. Available online: https://research.wur.nl/en/publications/canoco-reference-manual-and-canodraw-for-windows-users-guide-soft (accessed on 13 February 2022).
- Calegari, A.; Hargrove, W.L.; Rheinheimer, D.D.S.; Ralisch, R.; Tessier, D.; Tourdonnet, S.D.; Guimaraes, M. Impact of Long-Term No-Tillage and Cropping System Management on Soil Organic Carbon in an Oxisol: A Model for Sustainability. Agron. J. 2008, 100, 1013–1019. [Google Scholar] [CrossRef]
- Hunt, P.; Karlen, D.L.; Matheny, T.A.; Quisenberry, V.L. Changes in Carbon Content of a Norfolk Loamy Sand after 14 Years of Conservation of Conventional Tillage. J. Soil Water Conserv. 1996, 51, 255–258. [Google Scholar]
- Beare, M.H.; Cabrera, M.L.; Hendrix, P.F.; Coleman, D.C. Aggregate-Protected and Unprotected Organic Matter Pools in Conventional- and No-Tillage Soils. Soil Sci. Soc. Am. J. 1994, 58, 787–795. [Google Scholar] [CrossRef]
- Azooz, R.H.; Arshad, M.A. Tillage Effects on Thermal Conductivity of Two Soils in Northern British Columbia. Soil Sci. Soc. Am. J. 1995, 59, 1413–1423. [Google Scholar] [CrossRef]
- Hill, R.L. Long-Term Conventional and No-Tillage Effects on Selected Soil Physical Properties. Soil Sci. Soc. Am. J. 1990, 54, 161–166. [Google Scholar] [CrossRef]
- Fabrizzi, K.P.; García, F.O.; Costa, J.L.; Picone, L.I. Soil Water Dynamics, Physical Properties and Corn and Wheat Responses to Minimum and No-Tillage Systems in the Southern Pampas of Argentina. Soil Tillage Res. 2005, 81, 57–69. [Google Scholar] [CrossRef]
- Silva, J.H.S.; Deenik, J.L.; Yost, R.S.; Bruland, G.L.; Crow, S.E. Improving Clay Content Measurement in Oxidic and Volcanic Ash Soils of Hawaii by Increasing Dispersant Concentration and Ultrasonic Energy Levels. Geoderma 2014, 237–238, 211–223. [Google Scholar] [CrossRef]
- Hill, R.L.; Horton, R.; Cruse, R.M. Tillage Effects on Soil Water Retention and Pore Size Distribution of Two Mollisols. Soil Sci. Soc. Am. J. 1985, 49, 1264–1270. [Google Scholar] [CrossRef]
- Guzha, A.C. Effects of Tillage on Soil Microrelief, Surface Depression Storage and Soil Water Storage. Soil Tillage Res. 2004, 76, 105–114. [Google Scholar] [CrossRef]
- Reichert, J.M.; Akiyoshi, L.E.; Suzuki, S.; Reinert, J.; Horn, R.; Hå, I. Reference Bulk Density and Critical Degree-of-Compactness for No-till Crop Production in Subtropical Highly Weathered Soils. Soil Tillage Res. 2009, 102, 242–254. [Google Scholar] [CrossRef]
- Tormena, C.A.; Pires Da Silva, A.; Libardi, P.L. Soil Physical Quality of a Brazilian Oxisol under Two Tillage Systems Using the Least Limiting Water Range Approach. Soil Tillage Res. 1999, 52, 223–232. [Google Scholar] [CrossRef]
- Alvarez, R.; Steinbach, H.S. A Review of the Effects of Tillage Systems on Some Soil Physical Properties, Water Content, Nitrate Availability and Crops Yield in the Argentine Pampas. Soil Tillage Res. 2009, 104, 1–15. [Google Scholar] [CrossRef]
- Dam, R.F.; Mehdi, B.B.; Burgess, E.; Madramootoo, C.A.; Mehuys, G.R.; Callum, I.R. Soil Bulk Density and Crop Yield under Eleven Consecutive Years of Corn with Different Tillage and Residue Practices in a Sandy Loam Soil in Central Canada. Soil Tillage Res. 2005, 84, 41–53. [Google Scholar] [CrossRef]
- Suzuki, L.; Reichert, J.M.; Reinert, D.J. Degree of Compactness, Soil Physical Properties and Yield of Soybean in Six Soils under No-Tillage. Soil Res. 2013, 51, 311–321. [Google Scholar] [CrossRef]
- Vyn, T.J.; Raimbault, B.A. Long-Term Effect of Five Tillage Systems on Corn Response and Soil Structure. Agron. J. 1993, 85, 1074–1079. [Google Scholar] [CrossRef]
- Wander, M.M.; Bidart, M.G.; Aref, S. Tillage Impacts on Depth Distribution of Total and Particulate Organic Matter in Three Illinois Soils; Tillage Impacts on Depth Distribution of Total and Particulate Organic Matter in Three Illinois Soils. Soil Sci. Soc. Am. J. 1998, 62, 1704–1711. [Google Scholar] [CrossRef]
- Horn, R. Time Dependence of Soil Mechanical Properties and Pore Functions for Arable Soils; Time Dependence of Soil Mechanical Properties and Pore Functions for Arable Soils. Soil Sci. Soc. Am. J. 2004, 68, 1131–1137. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Water Infiltration and Soil Structure Related to Organic Matter and Its Stratification with Depth. Soil Tillage Res. 2002, 66, 197–205. [Google Scholar] [CrossRef]
- Lal, R.; Mahboubi, A.A.; Fausey, N.R. Long-Term Tillage and Rotation Effects on Properties of a Central Ohio Soil. Soil Sci. Soc. Am. J. 1994, 58, 517–522. [Google Scholar] [CrossRef]
- Hendrix, P.F.; Parmelee, R.W.; Crossley, D.A.J.; Coleman, D.C.; Odum, E.P.; Groffman, P.M. Detritus Food Webs in Conventional and No-Tillage Agroecosystems. Bioscience 1986, 36, 374–380. [Google Scholar] [CrossRef]
- Ferris, H.; Matute, M.M. Structural and Functional Succession in the Nematode Fauna of a Soil Food Web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Q.; Zhu, A.; Liang, W.; Zhang, J.; Steinberger, Y. Effects of Tillage and Residue Management on Soil Nematode Communities in North China. Ecol. Indic. 2012, 13, 75–81. [Google Scholar] [CrossRef]
- Lima, E.A.; Mattos, J.K.; Moita, A.W.; Gomes Carneiro, R.; Carneiro, R.M.D.G. Host Status of Different Crops for Meloidogyne Ethiopica Control. Trop. Plant Pathol. 2009, 34, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Lamondia, J.A.; Elmer, W.H.; Mervosh, T.L.; Cowles, R.S. Integrated Management of Strawberry Pests by Rotation and Intercropping. Crop Prot. 2002, 21, 837–846. [Google Scholar] [CrossRef]
- Webb, J.; Loveland, P.J.; Chambers, B.J.; Mitchell, R.; Garwood, T. The Impact of Modern Farming Practices on Soil Fertility and Quality in England and Wales. J. Agric. Sci. 2001, 137, 127–138. [Google Scholar] [CrossRef]
- Pittelkow, C.M.; Linquist, B.A.; Lundy, M.E.; Liang, X.; van Groenigen, K.J.; Lee, J.; van Gestel, N.; Six, J.; Venterea, R.T.; van Kessel, C. When Does No-till Yield More? A Global Meta-Analysis. Field Crops Res. 2015, 183, 156–168. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | Trial I | Trial II | ||||
|---|---|---|---|---|---|---|
| Properties y | BG | BO | SOL | BG | BO | SOL |
| Db (g/cm3) | 1.01 ± 0.02 z A | 1.06 ± 0.01 A | 0.95 ± 0.01 B | 1.08 ± 0.01 B | 1.16 ± 0.02 A | 1.05 ± 0.02 B |
| I (cm/hour) | 44.8 ± 5.43 B | 24.6 ± 5.82 B | 150.5 ± 21.84 A | 33.2 ± 9.3 A | 18.8 ± 6.8 A | 27.9 ± 7.5 A |
| TP (%) | 64.6 ± 0.9 B | 62.9 ± 0.5 B | 66.7 ± 0.4 A | 62.1 ± 0.4 A | 59.4 ± 0.6 B | 63.1 ± 0.6 A |
| MP (%) | 32.1 ± 2.0 A | 24.3 ± 1.9 B | 36.1 ± 1.4 A | 27.4 ± 1.0 B | 22.0 ± 0.8 C | 30.5 ± 1.4 A |
| SOM (%) | 1.08 ± 0.01 B | 1.45 ± 0.05 A | 0.98 ± 0.02 C | 1.11 ± 0.03 B | 1.46 ± 0.08 A | 0.94 ± 0.02 B |
| SM (θv) | 33.0 ± 0.6 B | 39.6 ± 0.5 A | 28.5 ± 0.5 C | 32.4 ± 0.4 B | 36.9 ± 0.3 A | 32.4 ± 0.2 B |
| FC (θv) | 32.5 ± 1.4 B | 38.5 ± 1.5 A | 30.6 ± 1.1 B | 34.8 ± 0.6 B | 37.4 ± 0.5 A | 32.6 ± 0.9 C |
| Temperature (°C) | BG | BO | SOL |
|---|---|---|---|
| -------------------------------------hours----------------------------------- | |||
| 15–20 | 143.8 z ±6.8 AB | 77.8 ± 8.3 B | 183.3 ± 32.6 A |
| 30–35 | 91.5 ± 12.1 A | 16.8 ± 5.1 B | 110.5 ± 14.1 A |
| 35–37 | 2.3 ± 1.3 B | 0.0 ± 0.0 B | 14.3 ± 2.8 A |
| ≥37 | 0.0 ± 0.0 B | 0.0 ± 0.0 B | 3.5 ± 2.2 A |
| -------------------------------------°C----------------------------------- | |||
| Max | 35.6 ± 0.5 A | 32.0 ± 0.5 B | 37.3 ± 0.4 A |
| Min | 17.0 ± 0.2 A | 17.8 ± 0.3 A | 16.6 ± 0.4 A |
| Nematode Parameters | Trial I | Trial II | ||||
|---|---|---|---|---|---|---|
| BG | BO | SOL | BG | BO | SOL | |
| Abundance | ----------------------------------------------- Number of nematodes/250 cm3 soil ----------------------------------- | |||||
| Helicotylenchus | 3 ± 3 z AB | 21 ± 10 A | 0 ± 0 B | 13 ± 8 AB | 16 ± 7 A | 1 ± 1 B |
| Meloidogyne | 10 ± 8 A | 3 ± 2 A | 137 ± 455 A | 133 ± 33 A | 76 ± 45 B | 336 ± 263 A |
| Paratrichodorus | 4 ± 2 A | 1 ± 1 A | 0 ± 0 A | 15 ± 6 A | 2 ± 1 A | 22 ± 18 A |
| Pratylenchus | 0 ± 0 A | 0 ± 0 A | 0 ± 0 A | 166 ± 52 A | 13 ± 7 B | 10 ± 6 B |
| Rotylenchulus | 152 ± 38 A | 288 ± 50 A | 48 ± 29 B | 611 ± 141 A | 464 ± 78 A | 627 ± 177 A |
| Bacterivore | 373 ± 105 A | 178 ± 29 B | 151 ± 51 C | 659 ± 167 A | 542 ± 108 A | 293 ± 33 B |
| Fungivore | 156 ± 35 A | 201 ± 26 A | 17 ± 12 B | 307 ± 46 A | 255 ± 51 A | 131 ± 30 B |
| Herbivore | 170 ± 45 A | 313 ± 51 A | 185 ± 136 B | 938 ± 144 A | 571 ± 86 A | 997 ± 297 A |
| Omnivore | 10 ± 5 A | 8 ± 4 A | 1 ± 1 A | 31 ± 7 AB | 49 ± 12 A | 17 ± 8 B |
| Predator | 2 ± 2 A | 1 ± 1 A | 0 ± 0 A | 3 ± 3 A | 4 ± 2 A | 1 ± 1 A |
| Indices y | ||||||
| % Bacterivore | 44.3 ± 4.5 A | 25.7 ± 3.4 B | 39.8 ± 10.1 B | 32.6 ± 4.3 A | 35.3 ± 4.0 AB | 27.4 ± 5.2 B |
| % Fungivore | 22.3 ± 3.3 B | 29.1 ± 3.2 A | 3.6 ± 1.9 C | 15.9 ± 1.6 A | 17.0 ± 2.4 A | 10.7 ± 2.5 B |
| % Herbivore | 28.0 ± 5.4 AB | 42.6 ± 3.8 A | 22.8 ± 8.3 B | 48.6 ± 4.9 AB | 42.4 ± 5.6 B | 60.1 ± 6.6 A |
| % Omnivore | 1.3 ± 0.3 A | 1.0 ± 0.5 A | 0.1 ± 0.1 A | 1.7 ± 0.4 B | 3.3 ± 0.6 A | 1.1 ± 0.5 B |
| % Predator | 0.3 ± 0.3 A | 0.1 ± 0.1 A | 0.0 ± 0.0 A | 0.2 ± 0.1 A | 0.3 ± 0.1 A | 0.0 ± 0.0 A |
| Diversity | 8.1 ± 1.3 A | 5.1 ± 0.6 AB | 4.4 ± 1.9 B | 7.0 ± 1.3 A | 6.8 ± 0.9 A | 11.7 ± 6.8 A |
| Dominance | 16.5 ± 3.2 A | 23.0 ± 2.8 A | 18.4 ± 5.4 A | 20.7 ± 3.9 B | 19.9 ± 3.8 B | 32.9 ± 7.6 A |
| Richness | 13 ± 2 A | 12 ± 1 A | 4 ± 1 B | 17 ± 1 A | 17 ± 1 A | 10 ± 1 B |
| CI | 31.8 ± 3.8 B | 55.8 ± 6.4 A | 20.8 ± 11.1 B | 36.4 ± 6.8 A | 36.9 ± 8.0 A | 58.3 ± 10.7 A |
| EI | 57.2 ± 2.8 A | 52.9 ± 1.6 A | 9.0 ± 5.8 B | 56.7 ± 3.5 A | 59.6 ± 6.5 A | 34.2 ± 5.5 B |
| F/F + B | 33.3 ± 4.1 B | 53.4 ± 5.0 A | 4.0 ± 2.0 C | 34.7 ± 4.1 A | 31.8 ± 3.6 A | 27.8 ± 4.8 A |
| MI | 1.9 ± 0.0 B | 2.0 ± 0.0 A | 1.3 ± 0.3 C | 1.9 ± 0.0 A | 2.0 ± 0.1 A | 2.0 ± 0.0 A |
| SI | 11.9 ± 5.2 A | 11.6 ± 3.8 A | 2.6 ± 1.9 B | 20.4 ± 2.7 B | 38.8 ± 4.6 A | 13.1 ± 4.5 B |
| Growth Parameters y | Trial I † | Trial II | ||||
|---|---|---|---|---|---|---|
| BG | BO | SOL | BG | BO | SOL | |
| Yield (kg/ha) | 3733 ± 143 z A | 3873 ± 712 A | 3923 ± 884 A | 6663 ± 420 A | 5808 ± 180 A | 6737 ± 536 A |
| Chlorophyll (SPAD) | 32.7 ± 1.9 B | 38.2 ± 1.4 A | 37.2 ± 1.4 A | 41.6 ± 1.7 B | 45.1 ± 1.4 A | 43.4 ± 1.5 AB |
| Height (cm) | 100.4 ± 15.9 A | 97.2 ± 15.9 A | 100.6 ± 15.1 A | 143.0 ± 22.1 A | 133.0 ± 21.7 B | 148.8 ± 23.4 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marquez, J.; Paudel, R.; Sipes, B.S.; Wang, K.-H. Successional Effects of No-Till Cover Cropping with Black Oat (Avena strigosa) vs. Soil Solarization on Soil Health in a Tropical Oxisol. Horticulturae 2022, 8, 527. https://doi.org/10.3390/horticulturae8060527
Marquez J, Paudel R, Sipes BS, Wang K-H. Successional Effects of No-Till Cover Cropping with Black Oat (Avena strigosa) vs. Soil Solarization on Soil Health in a Tropical Oxisol. Horticulturae. 2022; 8(6):527. https://doi.org/10.3390/horticulturae8060527
Chicago/Turabian StyleMarquez, Josiah, Roshan Paudel, Brent S. Sipes, and Koon-Hui Wang. 2022. "Successional Effects of No-Till Cover Cropping with Black Oat (Avena strigosa) vs. Soil Solarization on Soil Health in a Tropical Oxisol" Horticulturae 8, no. 6: 527. https://doi.org/10.3390/horticulturae8060527
APA StyleMarquez, J., Paudel, R., Sipes, B. S., & Wang, K. -H. (2022). Successional Effects of No-Till Cover Cropping with Black Oat (Avena strigosa) vs. Soil Solarization on Soil Health in a Tropical Oxisol. Horticulturae, 8(6), 527. https://doi.org/10.3390/horticulturae8060527