
Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Endophyte Isolation and Preparation of Inocula
2.2. Fruit
2.3. Pathogen Inoculum
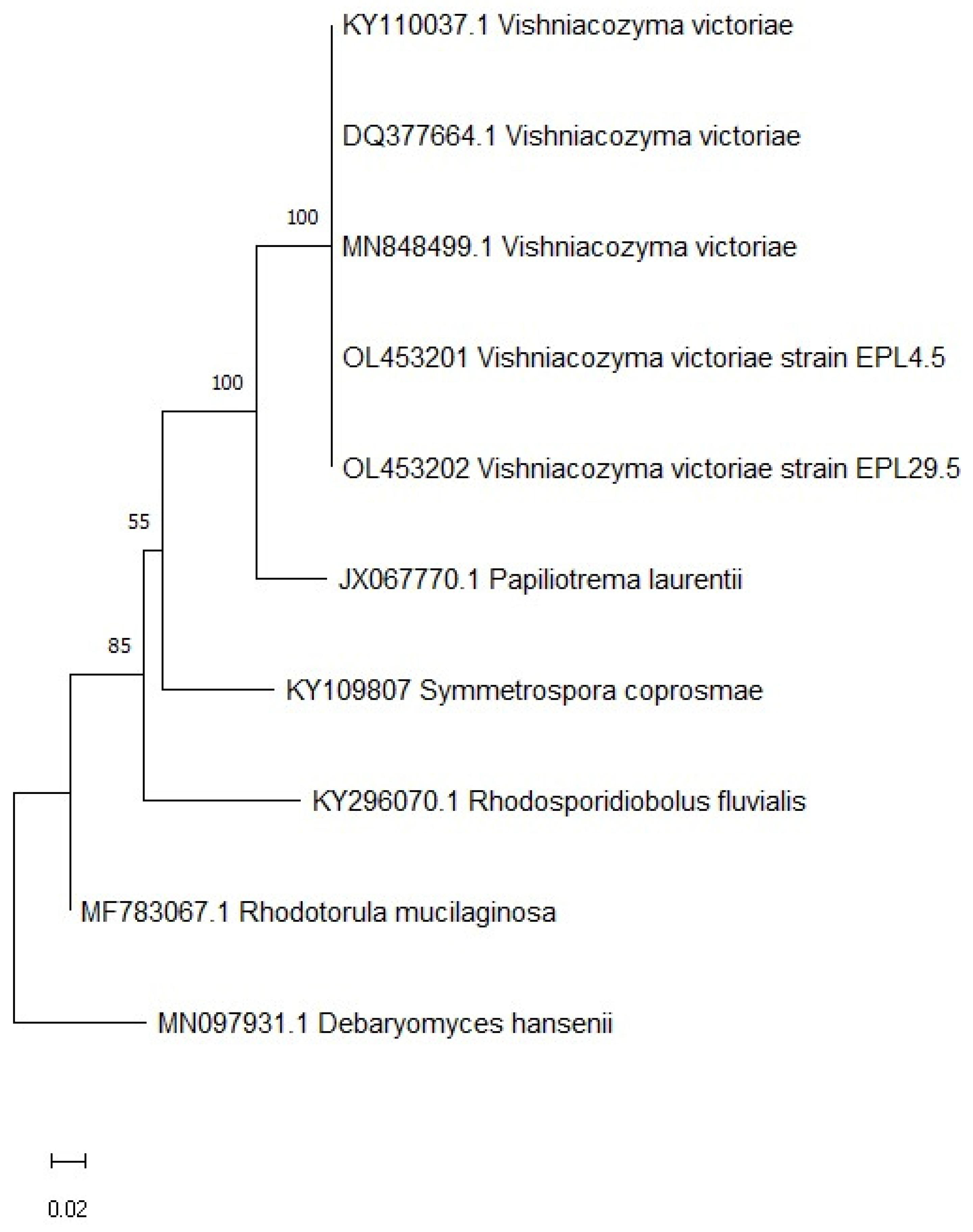
2.4. Selection and Identification of Yeasts as a Potential Biocontrol of P. vagabunda
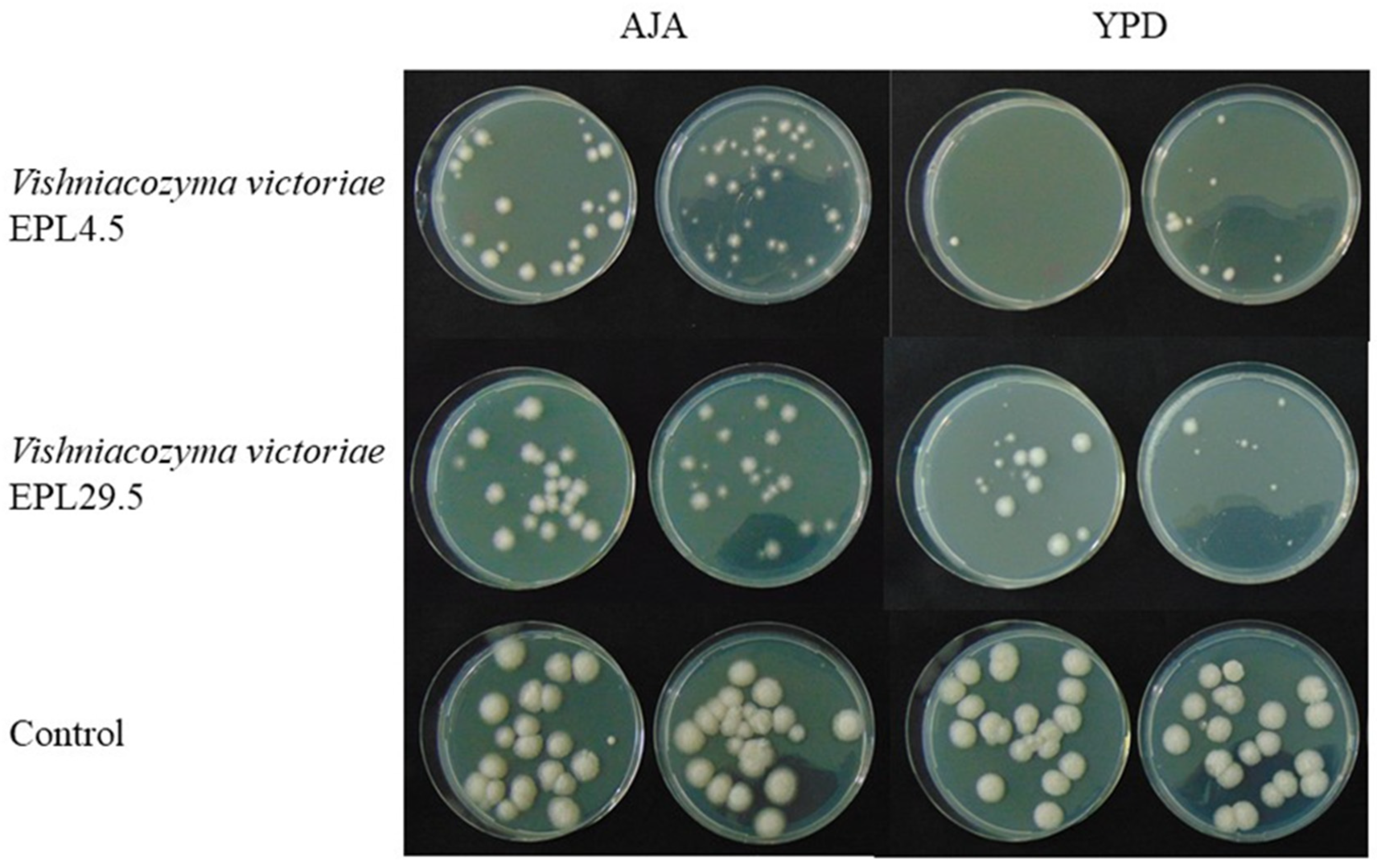
2.5. Antagonistic Activity of Yeast on the Fruit
2.6. Biofilm Formation by Yeast
2.7. Production of Volatile Antifungal Compounds
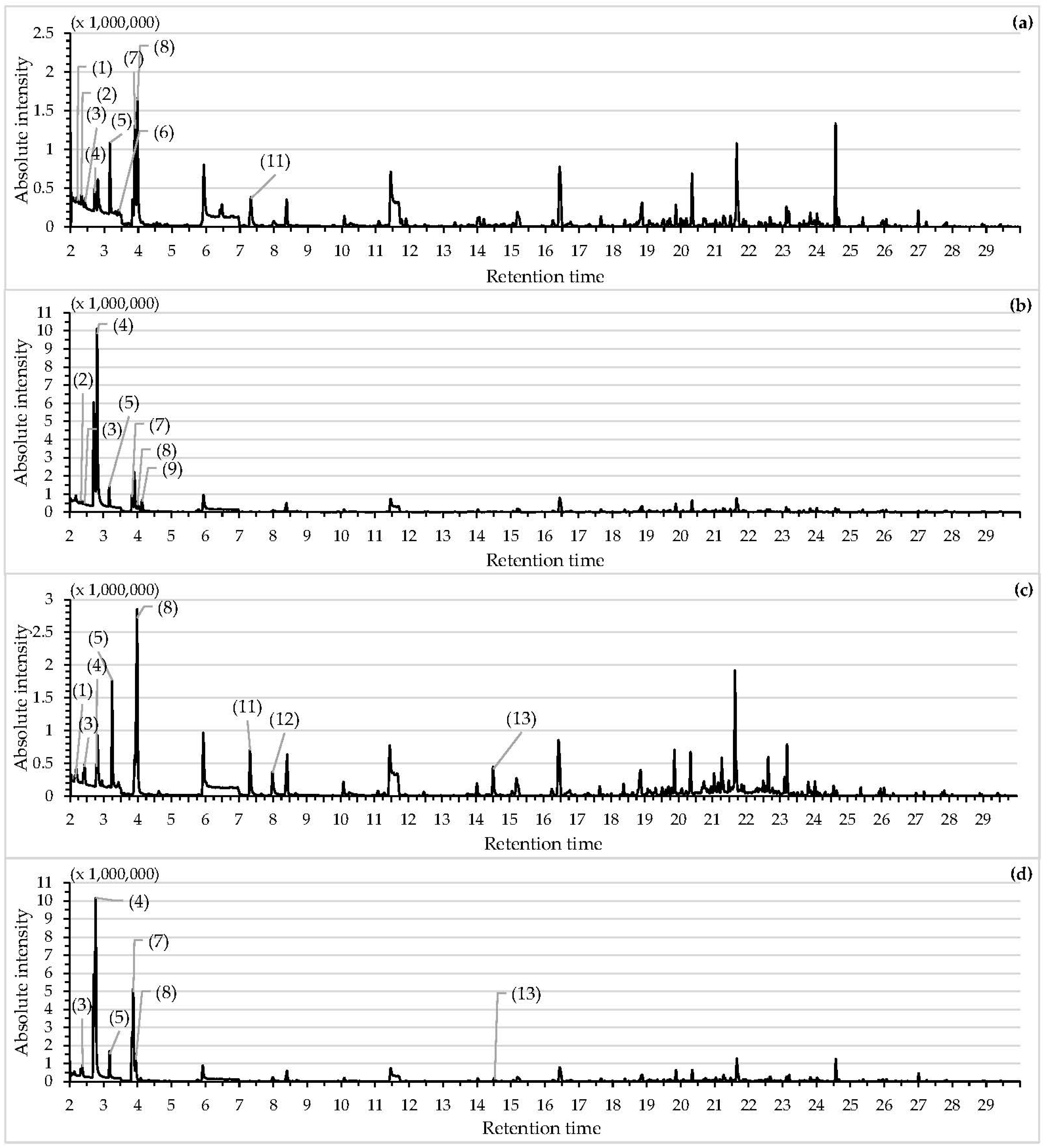
2.8. Chemical Characterization of Volatile Organic Compounds
2.9. Statistical Analysis
3. Results
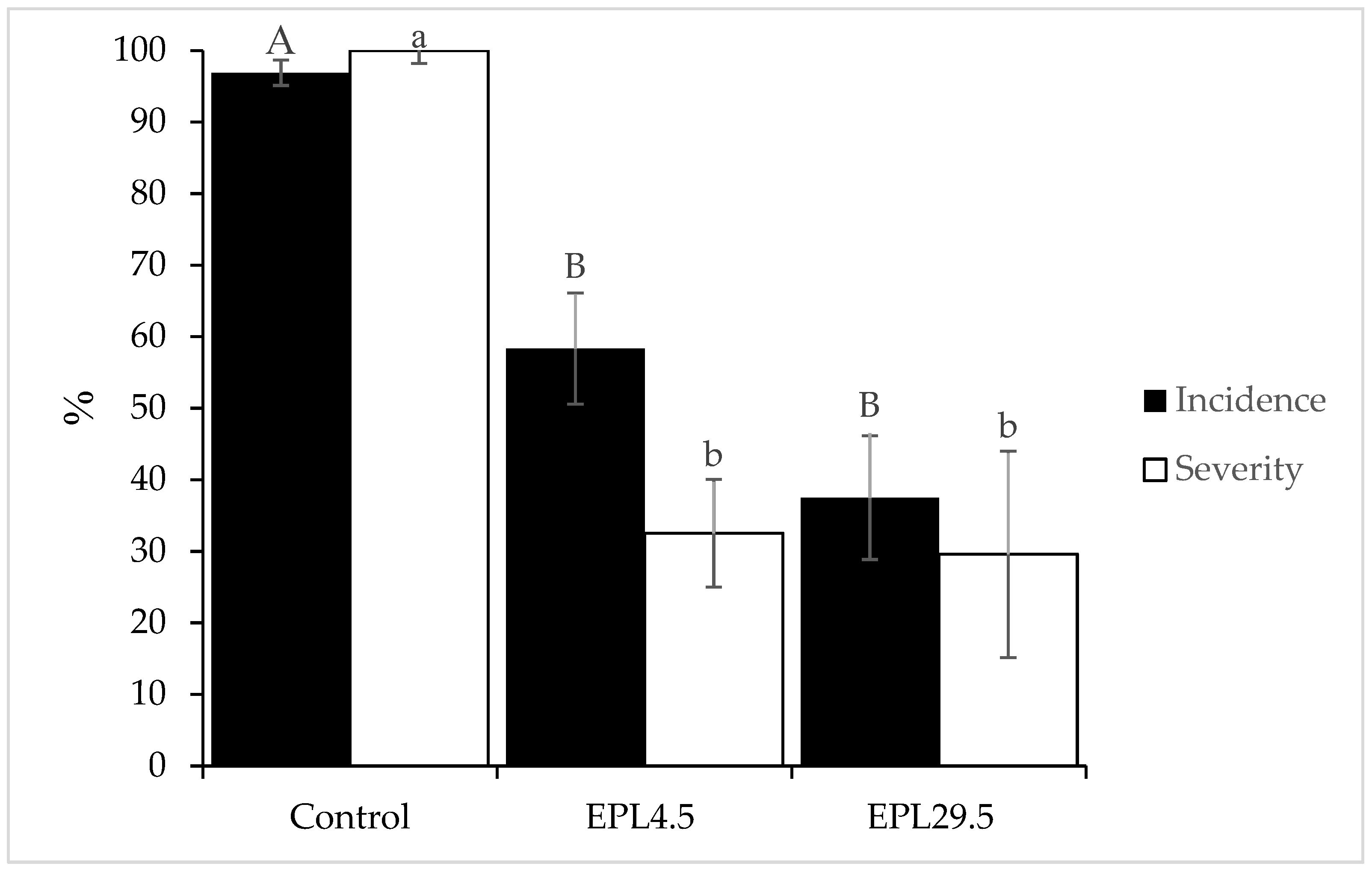
3.1. Antagonistic Activity of Yeast on Fruit
3.2. Biofilm Formation
3.3. Production of Volatile Antifungal Compounds
3.4. Chemical Characterization of Volatile Organic Compounds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soto-Alvear, S.; Lolas, M.; Rosales, I.M.; Chávez, E.R.; Latorre, B.A. Characterization of the Bull’s Eye Rot of Apple in Chile. Plant Dis. 2013, 97, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameldi, I.; Neri, F.; Menghini, M.; Pirondi, A.; Nanni, I.M.; Collina, M.; Mari, M. Characterization of Neofabraea Vagabunda Isolates Causing Apple Bull’s Eye Rot in Italy (Emilia-Romagna Region). Plant Pathol. 2017, 66, 1432–1444. [Google Scholar] [CrossRef]
- Henriquez, J.L.; Sugar, D.; Spotts, R.A. Etiology of Bull’s Eye Rot of Pear Caused by Neofabraea Spp. in Oregon, Washington, and California. Plant Dis. 2004, 88, 1134–1138. [Google Scholar] [CrossRef] [Green Version]
- Wood, P.N.; Fisher, B.M. The Effect of Fungicides on Spore Germination, Mycelial Growth and Lesion Development of Phlyctema Vagabunda (Syn: Neofabraea Alba) (Bull’s Eye Rot of Apples). N. Z. Plant Prot. 2017, 70, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, J.L. First Report of Apple Rot Caused by Neofabraea Alba in Chile. Plant Dis. 2005, 89, 1360. [Google Scholar] [CrossRef] [PubMed]
- Den Breeyen, A.; Rochefort, J.; Russouw, A.; Meitz-Hopkins, J.; Lennox, C.L. Preharvest Detection and Postharvest Incidence of Phlyctema Vagabunda on ‘Cripps Pink’ Apples in South Africa. Plant Dis. 2020, 104, 841–846. [Google Scholar] [CrossRef]
- Pešicová, K.; Kolařík, M.; Hortová, B.; Novotný, D. Diversity and Identification of Neofabraea Species Causing Bull’s Eye Rot in the Czech Republic. Eur. J. Plant Pathol. 2017, 147, 683–693. [Google Scholar] [CrossRef]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Control of Neofabraea Alba by Plant Volatile Compounds and Hot Water. Postharvest Biol. Technol. 2009, 51, 425–430. [Google Scholar] [CrossRef]
- Lolas, M.; Cáceres, M.; Reyes, J.A.; Díaz, G.A. Bull’s Eye Rot Development in Storage Is Related to the Timing of Apple Fruit Infection by Neofabraea Vagabunda in the Orchard in Chile. Acta Hortic. 2021, 1325, 73–76. [Google Scholar] [CrossRef]
- Lolas, M.; Díaz, G.; Mendez, R.; Cáceres, M.; Neubauer, L. Evaluation of the Efficacy of Fungicide Fludioxonil in the Postharvest Control of Bull’s Eye Rot (Neofabraea Alba) in Chile. Acta Hortic. 2016, 1144, 461–464. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of Biocontrol Products for Postharvest Diseases of Fruit: The Importance of Elucidating the Mechanisms of Action of Yeast Antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Wisniewski, M.; Wilson, C.; Droby, S.; Chalutz, E.; El-Ghaouth, A.; Stevens, C. Postharvest Biocontrol: New Concepts and Applications. In Biological Control: A Global Perspective; Vincent, C., Goettel, M.S., Lazarovits, G., Eds.; CABI: Wallingford, UK, 2007; pp. 262–273. ISBN 978-1-84593-265-7. [Google Scholar]
- Abdelhai, M.H.; Awad, F.N.; Yang, Q.; Mahunu, G.K.; Godana, E.A.; Zhang, H. Enhancement the Biocontrol Efficacy of Sporidiobolus Pararoseus Y16 against Apple Blue Mold Decay by Glycine Betaine and Its Mechanism. Biol. Control 2019, 139, 104079. [Google Scholar] [CrossRef]
- He, F.; Zhao, L.; Zheng, X.; Abdelhai, M.H.; Boateng, N.S.; Zhang, X.; Zhang, H. Investigating the Effect of Methyl Jasmonate on the Biocontrol Activity of Meyerozyma Guilliermondii against Blue Mold Decay of Apples and the Possible Mechanisms Involved. Physiol. Mol. Plant Pathol. 2020, 109, 101454. [Google Scholar] [CrossRef]
- Li, R.; Zhang, H.; Liu, W.; Zheng, X. Biocontrol of Postharvest Gray and Blue Mold Decay of Apples with Rhodotorula Mucilaginosa and Possible Mechanisms of Action. Int. J. Food Microbiol. 2011, 146, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Manso, T.; Nunes, C. Metschnikowia Andauensis as a New Biocontrol Agent of Fruit Postharvest Diseases. Postharvest Biol. Technol. 2011, 61, 64–71. [Google Scholar] [CrossRef]
- Mari, M.; Martini, C.; Spadoni, A.; Rouissi, W.; Bertolini, P. Biocontrol of Apple Postharvest Decay by Aureobasidium Pullulans. Postharvest Biol. Technol. 2012, 73, 56–62. [Google Scholar] [CrossRef]
- Navarta, L.G.; Calvo, J.; Posetto, P.; Cerutti, S.; Raba, J.; Benuzzi, D.; Sanz, M.I. Postharvest Control of Gray Mold in Apples with Lyophilized Formulations of Cryptococcus Laurentii: The Effect of Cold Stress in the Survival and Effectiveness of the Yeast. Food Bioprocess Technol. 2014, 7, 2962–2968. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Villalobos, M.C.; Calle, A.; Serradilla, M.J.; Córdoba, M.G.; Hernández, A. Yeasts Isolated from Figs (Ficus Carica L.) as Biocontrol Agents of Postharvest Fruit Diseases. Food Microbiol. 2016, 57, 45–53. [Google Scholar] [CrossRef]
- Türkel, S.; Korukluoğlu, M.; Yavuz, M. Biocontrol Activity of the Local Strain of Metschnikowia Pulcherrima on Different Postharvest Pathogens. Biotechnol. Res. Int. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Efficacy of the Antagonist Aureobasidium Pullulans PL5 against Postharvest Pathogens of Peach, Apple and Plum and Its Modes of Action. Biol. Control 2010, 54, 172–180. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Potential Biocontrol Activity of a Strain of Pichia Guilliermondii against Grey Mold of Apples and Its Possible Modes of Action. Biol. Control 2011, 57, 193–201. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Ma, L.; Dong, Y.; Jiang, S.; Xu, B.; Zheng, X. Biocontrol of Major Postharvest Pathogens on Apple Using Rhodotorula Glutinis and Its Effects on Postharvest Quality Parameters. Biol. Control 2009, 48, 79–83. [Google Scholar] [CrossRef]
- Wilson, C.L.; Wisniewski, M.E. Biological Control of Postharvest Diseases of Fruits and Vegetables: An Emerging Technology*. Annu. Rev. Phytopathol. 1989, 27, 425–441. [Google Scholar] [CrossRef]
- Ocampo-Suarez, I.B.; López, Z.; Calderón-Santoyo, M.; Ragazzo-Sánchez, J.A.; Knauth, P. Are Biological Control Agents, Isolated from Tropical Fruits, Harmless to Potential Consumers? Food Chem. Toxicol. 2017, 109, 1055–1062. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol Ability and Action Mechanism of Food-Isolated Yeast Strains against Botrytis Cinerea Causing Post-Harvest Bunch Rot of Table Grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Abo Elyousr, K.A.M.; Ismail, I.M. Biocontrol of Monilinia Fructigena, Causal Agent of Brown Rot of Apple Fruit, by Using Endophytic Yeasts. Biol. Control 2020, 144, 104239. [Google Scholar] [CrossRef]
- Chanchaichaovivat, A.; Panijpan, B.; Ruenwongsa, P. Putative Modes of Action of Pichia Guilliermondii Strain R13 in Controlling Chilli Anthracnose after Harvest. Biol. Control 2008, 47, 207–215. [Google Scholar] [CrossRef]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Álvarez, N.A.; Ragazzo-Sánchez, J.A. Action Mechanisms of the Yeast Meyerozyma Caribbica for the Control of the Phytopathogen Colletotrichum Gloeosporioides in Mangoes. Biol. Control 2013, 65, 293–301. [Google Scholar] [CrossRef]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Rodriguez Assaf, L.A.; Toro, M.E.; Castellanos de Figueroa, L.I.; Vazquez, F. Antifungal Modes of Action of Saccharomyces and Other Biocontrol Yeasts against Fungi Isolated from Sour and Grey Rots. Int. J. Food Microbiol. 2015, 204, 91–100. [Google Scholar] [CrossRef]
- Chan, Z.; Tian, S. Induction of H2O2-Metabolizing Enzymes and Total Protein Synthesis by Antagonistic Yeast and Salicylic Acid in Harvested Sweet Cherry Fruit. Postharvest Biol. Technol. 2006, 39, 314–320. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Ragazzo-Sánchez, J.A.; Barros-Castillo, J.C.; Sandoval-Contreras, T.; Calderón-Santoyo, M. Sodium Alginate Coatings Added with Meyerozyma Caribbica: Postharvest Biocontrol of Colletotrichum Gloeosporioides in Avocado (Persea Americana Mill. Cv. Hass). Postharvest Biol. Technol. 2020, 163, 111123. [Google Scholar] [CrossRef]
- Liu, Y.; Yao, S.; Deng, L.; Ming, J.; Zeng, K. Different Mechanisms of Action of Isolated Epiphytic Yeasts against Penicillium Digitatum and Penicillium Italicum on Citrus Fruit. Postharvest Biol. Technol. 2019, 152, 100–110. [Google Scholar] [CrossRef]
- Tian, Y.; Li, W.; Jiang, Z.; Jing, M.; Shao, Y. The Preservation Effect of Metschnikowia Pulcherrima Yeast on Anthracnose of Postharvest Mango Fruits and the Possible Mechanism. Food Sci. Biotechnol. 2018, 27, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, L.; Li, S.; Gao, X.; Wu, N.; Zhao, Y.; Sun, W. Control of Postharvest Grey Spot Rot of Loquat Fruit with Metschnikowia Pulcherrima E1 and Potential Mechanisms of Action. Biol. Control 2021, 152, 104406. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Xie, J.; Deng, L.; Yao, S.; Zeng, K. Biocontrol Efficacy of Pichia Membranaefaciens and Kloeckera Apiculata against Monilinia Fructicola and Their Ability to Induce Phenylpropanoid Pathway in Plum Fruit. Biol. Control 2019, 129, 83–91. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, W.; Zeng, J.; Shao, Y. Mechanisms of Action of the Yeast Debaryomyces Nepalensis for Control of the Pathogen Colletotrichum Gloeosporioides in Mango Fruit. Biol. Control 2018, 123, 111–119. [Google Scholar] [CrossRef]
- Glushakova, A.M.; Kachalkin, A.V. Endophytic Yeasts in Malus Domestica and Pyrus Communis Fruits under Anthropogenic Impact. Microbiology 2017, 86, 128–135. [Google Scholar] [CrossRef]
- Cao, D.; Li, X.; Cao, J.; Wang, W. PCR Detection of the Three Neofabraea Pathogenic Species Responsible for Apple Bull’s Eye Rot. Adv. Microbiol. 2013, 03, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Vero, S.; Mondino, P.; Burgueño, J.; Soubes, M.; Wisniewski, M. Characterization of Biocontrol Activity of Two Yeast Strains from Uruguay against Blue Mold of Apple. Postharvest Biol. Technol. 2002, 26, 91–98. [Google Scholar] [CrossRef]
- Růžička, F.; Holá, V.; Votava, M.; Tejkalová, R. Importance of Biofilm InCandida Parapsilosis and Evaluation of Its Susceptibility to Antifungal Agents by Colorimetric Method. Folia Microbiol. 2007, 52, 209–214. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Foggia, M.; Baraldi, E. Aureobasidium Pullulans Volatile Organic Compounds as Alternative Postharvest Method to Control Brown Rot of Stone Fruits. Food Microbiol. 2020, 87, 103395. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of Volatile Organic Compounds by Aureobasidium Pullulans as a Potential Mechanism of Action against Postharvest Fruit Pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Jaibangyang, S.; Nasanit, R.; Limtong, S. Biological Control of Aflatoxin-Producing Aspergillus Flavus by Volatile Organic Compound-Producing Antagonistic Yeasts. BioControl 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of Yeast Volatile Organic Compounds on Ochratoxin A-Producing Aspergillus Carbonarius and A. Ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Cheng, X.; Zhou, H.; Si, L. Hanseniaspora Uvarum Prolongs Shelf Life of Strawberry via Volatile Production. Food Microbiol. 2017, 63, 205–212. [Google Scholar] [CrossRef]
- Lutz, M.C.; Lopes, C.A.; Sosa, M.C.; Sangorrín, M.P. Semi-Commercial Testing of Regional Yeasts Selected from North Patagonia Argentina for the Biocontrol of Pear Postharvest Decays. Biol. Control 2020, 150, 104246. [Google Scholar] [CrossRef]
- Villalba, M.L.; Lutz, M.C.; Lopez, S.; Pildain, M.B.; Sangorrín, M.P. Patagonian Antagonist Yeasts for Food Biopreservation. In Biology and Biotechnology of Patagonian Microorganisms; Olivera, N.L., Libkind, D., Donati, E., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 301–323. ISBN 978-3-319-42799-7. [Google Scholar]
- Sui, Y.; Wisniewski, M.; Droby, S.; Liu, J. Responses of Yeast Biocontrol Agents to Environmental Stress. Appl. Environ. Microbiol. 2015, 81, 2968–2975. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia Laxa by Aureobasidium Pullulans Strains: Insights on Competition for Nutrients and Space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.; Zhou, Y.; Yao, S.; Deng, L.; Zeng, K. Isolation, Identification and in Vitro Screening of Chongqing Orangery Yeasts for the Biocontrol of Penicillium Digitatum on Citrus Fruit. Biol. Control 2017, 110, 18–24. [Google Scholar] [CrossRef]
- Pesce, V.M.; Nally, M.C.; Carrizo, G.P.; Rojo, C.; Pérez, B.A.; Toro, M.E.; Castellanos de Figueroa, L.I.; Vazquez, F. Antifungal Activity of Native Yeasts from Different Microenvironments against Colletotrichum Gloeosporioides on Ripe Olive Fruits. Biol. Control 2018, 120, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Lutz, M.C.; Lopes, C.A.; Rodriguez, M.E.; Sosa, M.C.; Sangorrín, M.P. Efficacy and Putative Mode of Action of Native and Commercial Antagonistic Yeasts against Postharvest Pathogens of Pear. Int. J. Food Microbiol. 2013, 164, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.N.; Kupper, K.C. Biofilm Production by Aureobasidium Pullulans Improves Biocontrol against Sour Rot in Citrus. Food Microbiol. 2018, 69, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Martini, C.; Mari, M. Biological Control of Postharvest Diseases by Microbial Antagonists: How Many Mechanisms of Action? Eur. J. Plant Pathol. 2016, 145, 711–717. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile Organic Compounds Produced by Antarctic Strains of Candida Sake Play a Role in the Control of Postharvest Pathogens of Apples. Biol. Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Konsue, W.; Dethoup, T.; Limtong, S. Biological Control of Fruit Rot and Anthracnose of Postharvest Mango by Antagonistic Yeasts from Economic Crops Leaves. Microorganisms 2020, 8, 317. [Google Scholar] [CrossRef] [Green Version]
- Medina-Córdova, N.; López-Aguilar, R.; Ascencio, F.; Castellanos, T.; Campa-Córdova, A.I.; Angulo, C. Biocontrol Activity of the Marine Yeast Debaryomyces Hansenii against Phytopathogenic Fungi and Its Ability to Inhibit Mycotoxins Production in Maize Grain (Zea Mays L.). Biol. Control 2016, 97, 70–79. [Google Scholar] [CrossRef]
- Fialho, M.B.; Toffano, L.; Pedroso, M.P.; Augusto, F.; Pascholati, S.F. Volatile Organic Compounds Produced by Saccharomyces Cerevisiae Inhibit the in Vitro Development of Guignardia Citricarpa, the Causal Agent of Citrus Black Spot. World J. Microbiol. Biotechnol. 2010, 26, 925–932. [Google Scholar] [CrossRef]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of Postharvest Botrytis Fruit Rot of Strawberry by Volatile Organic Compounds of Candida Intermedia. Phytopathology 2011, 101, 859–869. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strain | Biofilm Formation | |
|---|---|---|
| Absorbance (A620) | ||
| EPL4.5 | 0.023 ± 0.003 | − |
| EPL29.5 | 0.061 ± 0.006 | + |
| Yeast Strain | Colony Forming Unit Inhibition (%) | Mycelial Growth Inhibition (%) | ||
|---|---|---|---|---|
| YPD | AJA | YPD | AJA | |
| EPL4.5 | 54.84 ± 27.4 | 10.38 ± 21.9 | 69.14 ± 3.6 * | 42.1 ± 7.4 |
| EPL29.5 | 50.63 ± 7.6 | 19.44 ± 7.3 | 66.10 ± 12.1 | 43.62 ± 2.4 |
| Peak Number | RT(min) | Possible Compound | Molecular Formula | m/z | Relative Abundance (%) | Reference | |||
|---|---|---|---|---|---|---|---|---|---|
| EPL4.5 | EPL29.5 | ||||||||
| AJA | YPD | AJA | YPD | ||||||
| 1 | 2.14 | Hexane # | C6H14 | 41 | 0.49 | N.D. | 2.15 | N.D. | |
| 2 | 2.32 | Isobutyl chloride | C4H9Cl | 43 | 0.65 | 1.83 | N.D. | N.D. | |
| 3 | 2.41 | 1-Propanol, 2-methyl | C4H10O | 43 | 0.46 | 0.53 | 2.19 | 1.97 | [43,44] |
| 4 | 2.77 | 1-Butanol | C4H10O | 56 | 3.98 | 46.34 | 5.25 | 38.7 | |
| 5 | 3.18 | Silanediol, dimethy l # | C2H8O2Si | 77 | 3.97 | 2.35 | 7.82 | 2.69 | |
| 6 | 3.43 | 2,5-Dimethylfuran | C6H8O | 45 | 1.08 | N.D. | N.D. | N.D. | |
| 7 | 3.87 | 1-Butanol, 3-methyl | C5H12O | 56 | 1.94 | 8.18 | N.D. | 17.63 | [32,35,43] |
| 8 | 3.98 | 1-Butanol, 2-methyl-, (S)- | C5H12O | 41 | 19.06 | 1.33 | 13.27 | 2.97 | [32,43,44] |
| 9 | 4.13 | Disulfide, dimethyl | C2H6S2 | 94 | N.D. | 2.47 | N.D. | N.D. | [45] |
| 10 | 5.95 | Cyclotrisiloxane, hexamethyl- # | C6H18O3Si3 | 207 | 5.55 | 3.25 | N.D. | N.D. | |
| 11 | 7.32 | 1-Hexanol | C6H14O | 56 | 3.39 | N.D. | 3.27 | N.D. | [35,46] |
| 12 | 7.99 | 2-Heptanone | C7H14O | 43 | N.D. | N.D. | 1.65 | N.D. | [45] |
| 13 | 14.51 | 2-Nonanone | C9H18O | 43 | N.D. | N.D. | 2.04 | 0.49 | [35] |
| 14 | 16.45 | Cyclopentasiloxane, decamethyl # | C10H30O5Si5 | 73 | N.D. | N.D. | 3.99 | 1.34 | |
| 15 | 20.35 | Cyclohexasiloxane, dodecamethyl # | C12H36O6Si6 | 88 | N.D. | N.D. | 2.94 | N.D. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepúlveda, X.; Silva, D.; Ceballos, R.; Vero, S.; López, M.D.; Vargas, M. Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples. Horticulturae 2022, 8, 535. https://doi.org/10.3390/horticulturae8060535
Sepúlveda X, Silva D, Ceballos R, Vero S, López MD, Vargas M. Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples. Horticulturae. 2022; 8(6):535. https://doi.org/10.3390/horticulturae8060535
Chicago/Turabian StyleSepúlveda, Ximena, Diego Silva, Ricardo Ceballos, Silvana Vero, María Dolores López, and Marisol Vargas. 2022. "Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples" Horticulturae 8, no. 6: 535. https://doi.org/10.3390/horticulturae8060535
APA StyleSepúlveda, X., Silva, D., Ceballos, R., Vero, S., López, M. D., & Vargas, M. (2022). Endophytic Yeasts for the Biocontrol of Phlyctema vagabunda in Apples. Horticulturae, 8(6), 535. https://doi.org/10.3390/horticulturae8060535