Abstract
Jujube is a characteristic light-demanding fruit tree, but fruit quality is frequently degraded because of the weak light in protected cultivation facilities in south China. In this study, a black shade net was used to set three light intensities artificially: no shading (0%, control, NS), moderate shading (45%, MS), and severe shading (76%, SS) to investigate the effect of low light on sugar metabolism of jujube. Compared with NS, chlorophyll and carotenoid contents in shaded leaves were increased, while specific leaf weight was decreased. A decrease in sucrose content, rather than fructose and glucose, was the main factor affecting sugar content in shaded leaves and fruits. Low expression of ZjSS2 and ZjSS3 inhibited sugar synthesis in shaded leaves, thereby reducing sucrose content by 23.7% in SS leaf than that in NS. Compared with NS, sucrose content in SS fruit was decreased by 58.1%, which was mainly caused by the down-regulated expression of ZjSS2 and ZjSPS1. In conclusion, sugar accumulation in leaf and fruit with shading treatment was suppressed, which was associated with differential transcription levels of related metabolic genes. The study provides new insights into sugar metabolism in leaf and fruit of phototropic plants under low light environments, such as jujube.
1. Introduction
Light is one of the most important environmental factors affecting plant growth, nutrient absorption, yield, and quality of fruit [1]. Low light intensity is a crucial element restricting fruit production in some regions, being influenced by global climate change, geographical location, and cultivation facility [2,3]. Insufficient light could decrease fruit weight and yield [4], reduce fruit quality [5,6,7], and even promote abscission [8,9]. Nevertheless, suitable shading treatment improved the peel color of mango [10] and promoted dry matter accumulation of grape [11], providing an alternative strategy to achieve higher yield and promote improved fruit quality under heat stress [12]. In summary, low light intensity affects various fruit species differently.
Sugar plays a crucial role in the intrinsic quality of fruits [13] and is comprehensively regulated by multiple genes encoding enzymes. Sucrose synthase (SS, SUS) regulates fructose synthesis into sucrose, along with glucose synthesis into sucrose under the action of hexokinase (HK), uridine diphosphate glucose pyrophosphorylase (UGP), and sucrose phosphosynthase (SPS), while invertase (INV) catalyzes sucrose degradation into fructose and glucose [14,15]. Although the proportions and concentrations of sugar are primarily set by genetic factors under natural conditions, the growing environment, and especially light intensity, is also centrally important for these traits [16]. Sugar content was decreased or increased in fruit grown under low light conditions, thus altering sweetness [17,18,19,20,21]. However, studies on the molecular mechanisms affecting sugar content in leaf and fruit under shading conditions are rare, especially in phototropic plants.
Jujube (Ziziphus jujuba Mill.) is native to China, and its cultivation history can be traced back some 7000 years. Jujube is the most valuable species of the family Rhamnaceae in terms of its economic, ecological, and societal importance [22,23]. The cloudy, rainy, weak-light climate of southern production areas is quite different from the sunny conditions ideally needed for jujube [24]. Protected cultivation could alleviate fruit cracking and yield reduction of jujube caused by continuous summer rainfall in southern production areas and should be promoted vigorously, but also further reduce light intensity. Hence, low light is the main limitation restricting the growth and industrial development of jujube in south China. In our previous study, shading significantly decreased the weight, size, and soluble sugar content of jujube fruit but increased titratable acid content, thus degrading fruit quality [25], but the underlying mechanisms remained unknown. Therefore, the objective of the present work was to explore sugar accumulation patterns and dynamic changes in the transcription of related genes in jujube grown under low light by artificial shading, providing a theoretical basis for scientific cultivation and fruit quality improvement of jujube in southern areas with high humidity and little sunshine.
2. Materials and Methods
2.1. Materials
Jujube cultivar ‘Luojiangtiaoyuanzao’ was collected in Qiquan Town, Chongzhou City, Sichuan Province, China (103°07′ E, 30°30′ N). The site experiences a subtropical humid monsoon climate, with an annual average temperature of 15.9 °C, annual rainfall of 1012 mm, and annual sunshine duration of 1162 h. The average solar illumination during the experiment was about 28,000 lx. The terrain of the site is flat and the soil is mainly paddy soil.
The experiment commenced on 29 June 2018, about 45 d after anthesis. Three treatments were applied, namely no shading (shading rate 0%, NS, control), moderate shading (shading rate 46%, MS), and severe shading (shading rate 75%, SS). The shading rates were achieved using black sunshade nets of different densities. The illuminance was measured at different times during the day with an illuminance meter, and the shading rates of MS and SS were calculated relative to NS as control. The sunshade nets were located 0.5 m above the jujube tree, and the canopy was completely covered, but the trunk was not covered to facilitate gas exchange. Each treatment was performed on 10 trees and repeated three times for a total of 90 trees. 30 Leaves at node 4–7 of the fruit-bearing shoot and 30 fruits of each treatment were collected approximately every 15 d after shading, i.e., 0 d (before shading), then at 16 d, 31 d, 46 d, 59 d (mature stage). Samples were chopped and frozen with liquid nitrogen, then stored at −80 °C. Three analytical replicates were performed for each analysis.
2.2. Determination of Photosynthetic Pigments and Specific Leaf Weight
The photosynthetic pigments content was determined by acetone extraction [2]. Fresh leaves (0.5 g) were ground with 5 mL acetone and extracted in the dark for 24 h at 4 °C. The absorbance at different wavelengths (663 nm, 645 nm, and 480 nm) was measured using a spectrophotometer (SUMMIT, China) to calculate chlorophyll a (chl a), chlorophyll b (chl b), total chlorophyll (total chl), and carotenoid content.
The specific leaf weight was expressed as the ratio of total leaf dry weight to total leaf area (mg/cm2) [26]. The leaf area was determined by the small square method. Each group of leaves was dried at 80 °C to constant weight, and then dry weight was measured using an analytical balance.
2.3. Determination of Content and Composition of Soluble Sugar
The contents and compositions of sugar were determined according to Gao et al. [27], using high-performance liquid chromatography (HPLC). The fresh leaf or fruit (0.5 g) was ground into a powder with liquid nitrogen, extracted in 5 mL ethanol–water (90:10, V:V), then heated in a water bath at 80 °C for 30 min. The homogenate was centrifuged at 6000 rpm for 10 min at room temperature. Leaf extract was used directly for determination, while the fruit was extracted twice, and the supernatants were combined and adjusted to 25 mL. The extracts were filtered through a 0.22 μm membrane. The concentrations in the filtrate were determined using an NH2 column (250 mm × 4.6 mm, 5 μm) and a refractive index detector. The mobile phase comprised 75% acetonitrile and 25% distilled water. The flow rate was set to 1 mL/min and the injection volume was 10 μL. The column temperature was set at 40 °C.
2.4. RNA Extraction, cDNA Synthesis, and Genes Expression Analysis
Total RNA was extracted by the CTAB method. RNA quality and quantity were evaluated by 1% agarose gel electrophoresis and ultrafine UV spectrophotometer (Nanodrop one; Thermo Scientific, MA, USA). cDNA synthesis was performed according to the instructions of ReverTra Ace qPCR RT Master Mix with a gDNA Remover kit (Toyobo, Osaka, Japan). The sequences of genes were identified in the jujube genome. UBQ was used as the reference gene for normalization. Primer sequences of related genes were designed using Primer 5.0 and are listed in Supplementary Table S1.
Amplification was performed on CFX 96 (Bio-RAD, CA, USA) real-time quantitative PCR (qRT-PCR) using an SYBR® Premix Ex Taq™ II kit (TaKaRa, Osaka, Japan). The 10 µL reaction system contained SYBR Premix Ex Taq II (2×) (5 µL), 10 µM primers (0.4 µL each), ddH2O (3.2 µL), and diluted cDNA (1 µL). The conditions for qRT-PCR amplifications were as follows: 95 °C for 30 s, followed by 40 cycles of 5 s at 95 °C, 30 s at Tm, and 30 s at 72 °C. At the end of each experiment, a melt-curve analysis was carried out with the default parameters. Gene expression levels were calculated using the formula 2−∆∆Ct.
2.5. Statistical Analysis
Statistical analysis was conducted using SPSS 20.0 statistical software (IBM, NYS, USA). Data were analyzed by one-way analysis of variance, with significant differences (Duncan’s multiple range test) assessed at the 5% confidence level. Pearson’s correlation analysis was carried out between the content of soluble sugar and organic acid vs. gene expression.
3. Results
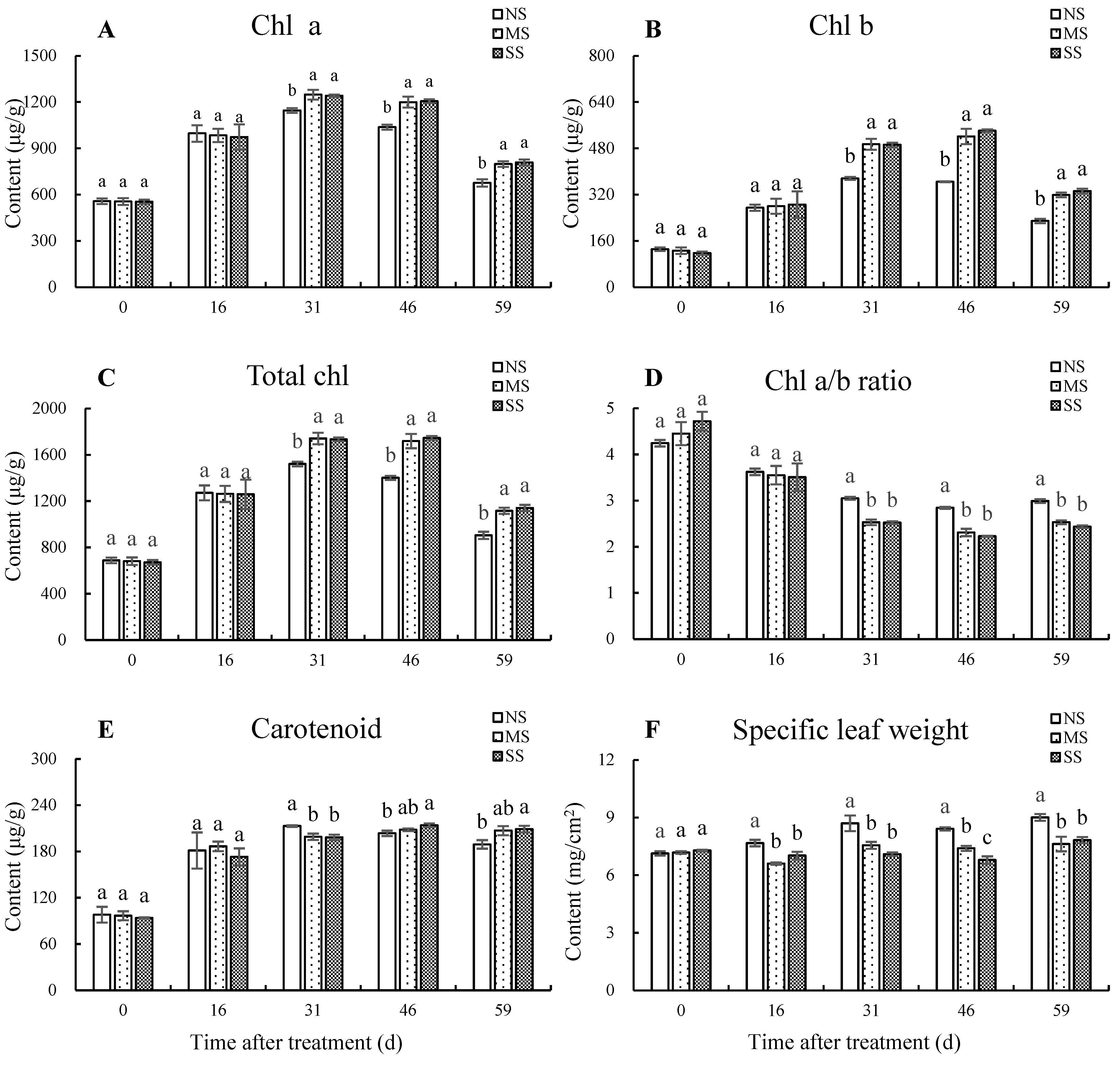
3.1. Dynamic Changes of Photosynthetic Pigments and Specific Leaf Weight
The chlorophyll contents increased first and then decreased, while carotenoid content gradually increased during leaf development (Figure 1). The contents of chl a, chl b, and total chl were significantly higher with MS and SS treatment for 31 d, 46 d, and 59 d than with NS. Compared with NS after shading for 59 d, chl a content in leaves under MS and SS treatments were significantly increased by 18.1% and 19.8%, chl b content by 39.2% and 45.2%, and total chl content by 23.5% and 26.2%, respectively. The carotenoid contents in SS leaves were increased by 5.1% (p < 0.05) and 10.5% (p < 0.05) at 46 d and 59 d, respectively, in comparison with NS. Shading accelerated the decline of chl a/b ratio for 31 d, 46 d, and 59 d. Compared with NS, chl a/b ratio in MS and SS leaves significantly decreased by 15.26% and 18.53% after shading for 59 d, respectively. In addition, the specific leaf weight under MS was significantly reduced by 14.0%, 13.2%, 12.1%, and 15.36% at 16 d, 31 d, 46 d, and 59 d, respectively, and under SS was significantly decreased by 8.5%, 18.5%, 19.3%, and 13.2%, respectively, compared with leaves with NS.
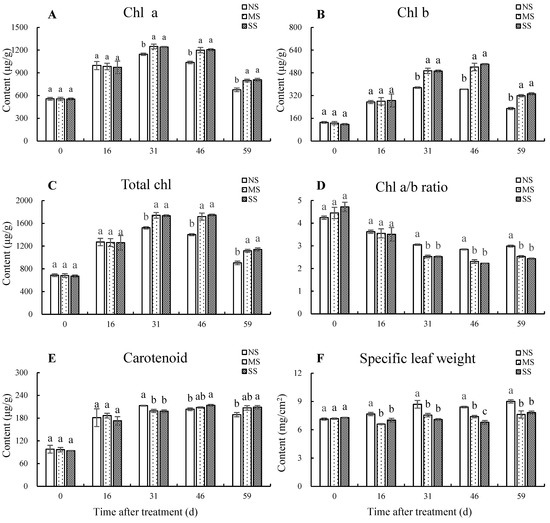
Figure 1.
Dynamic changes in photosynthetic pigments and specific leaf weight. (A) Chl a content. (B) Chl b content. (C) Total chl content (D) Chl a/b ratio. (E) Carotenoid content. (F) Specific leaf weight. Total chl content = chl a content + chl b content, Chl a/b ratio = chl a content/chl b content. Values are means of three replicates ± SD and different letters indicate significant differences (Duncan′s multiple range tests, p < 0.05) among three treatments at the same developmental stage.
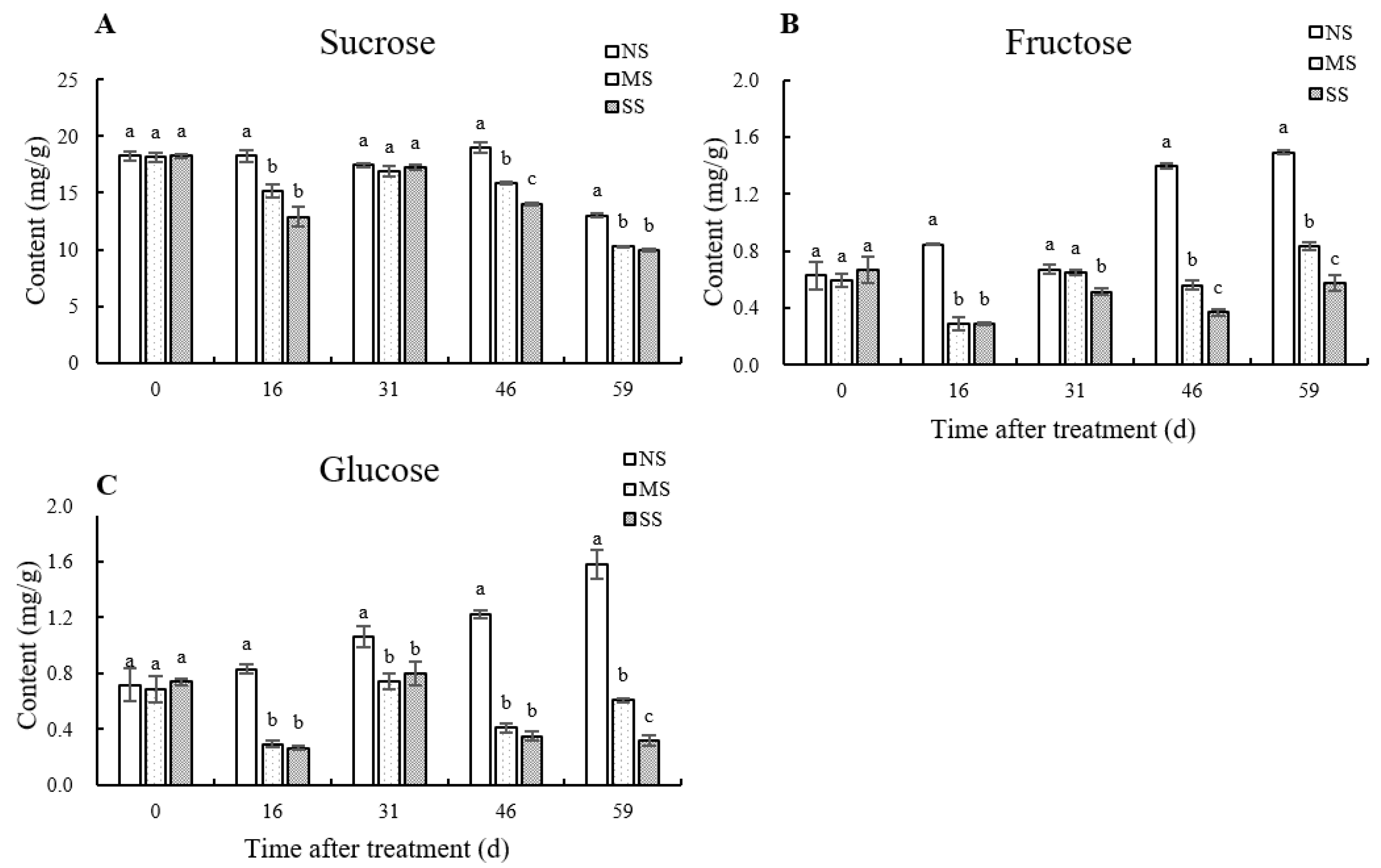
3.2. Dynamic Changes of Soluble Sugar Contents in Leaves
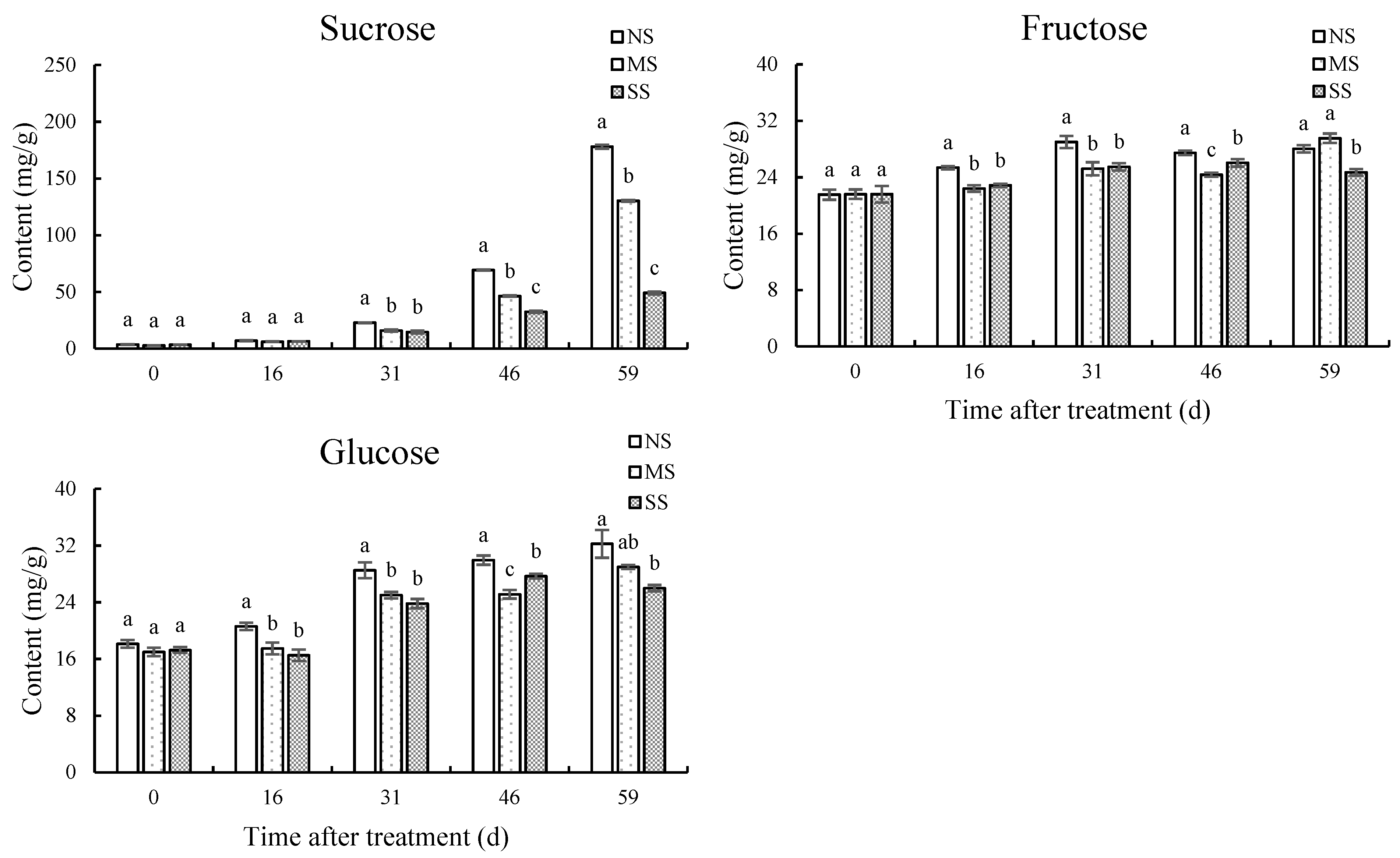
The contents of sucrose, fructose, and glucose in jujube leaves were significantly reduced by shading (Figure 2). Compared with NS, sucrose, fructose, and glucose contents in MS leaves at 59 d were significantly reduced by 20.7%, 44.1%, and 61.5%, and in SS leaves by 23.7%, 61.4%, and 79.9%, respectively. After shading for 16 d, 46 d, and 59 d, the sucrose contents of MS leaves were significantly reduced by 16.83%, 16.41%, and 20.73%, and those of SS by 29.37%, 26.04%, and 23.74%, respectively, in comparison with NS.
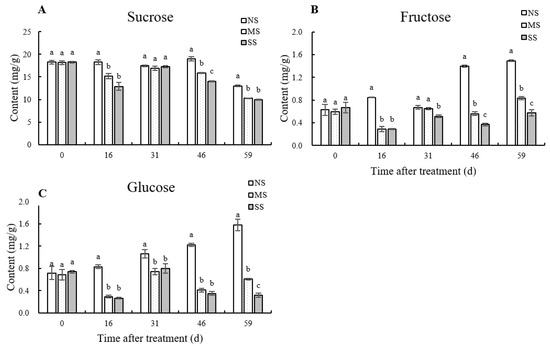
Figure 2.
Dynamic changes in soluble sugar content in leaves. (A) Sucrose content. (B) Fructose content. (C) Glucose content. Values are means of three replicates ± SD and different letters indicate significant differences (Duncan′s multiple range tests, p < 0.05) among three treatments at the same developmental stage.
3.3. Dynamic Changes of Soluble Sugar Contents in Fruits
The contents of sucrose, fructose, and glucose increased gradually during fruit development, while shading treatments significantly inhibited sugar accumulation (Figure 3). Compared with NS fruits, after shading for 31 d, 46 d, and 59 d, the sucrose contents of MS fruits were significantly reduced by 29.9%, 33.2%, and 26.8%, respectively. In SS fruits, the sucrose content was significantly reduced by 35.6%, 53.1%, and 72.4% at 31 d, 46 d, and 59 d, respectively, in comparison with NS. Compared with NS, the fructose and glucose contents in SS fruit at 59 d were reduced by 11.9% (p < 0.05) and 19.4% (p < 0.05), respectively, while there was no significant difference in MS.
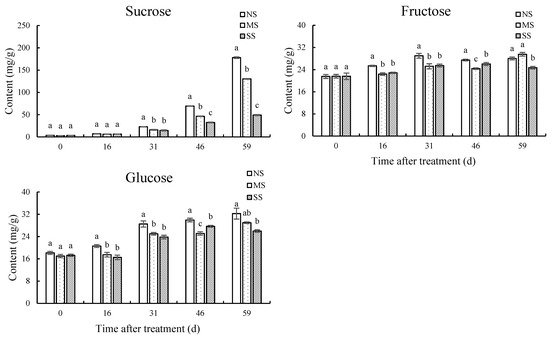
Figure 3.
Dynamic changes in soluble sugar contents in fruits. (A) Sucrose content. (B) Fructose content. (C) Glucose content. Values are means of three replicates ± SD and different letters indicate significant differences (Duncan′s multiple range tests, p < 0.05) among three treatments at the same developmental stage.
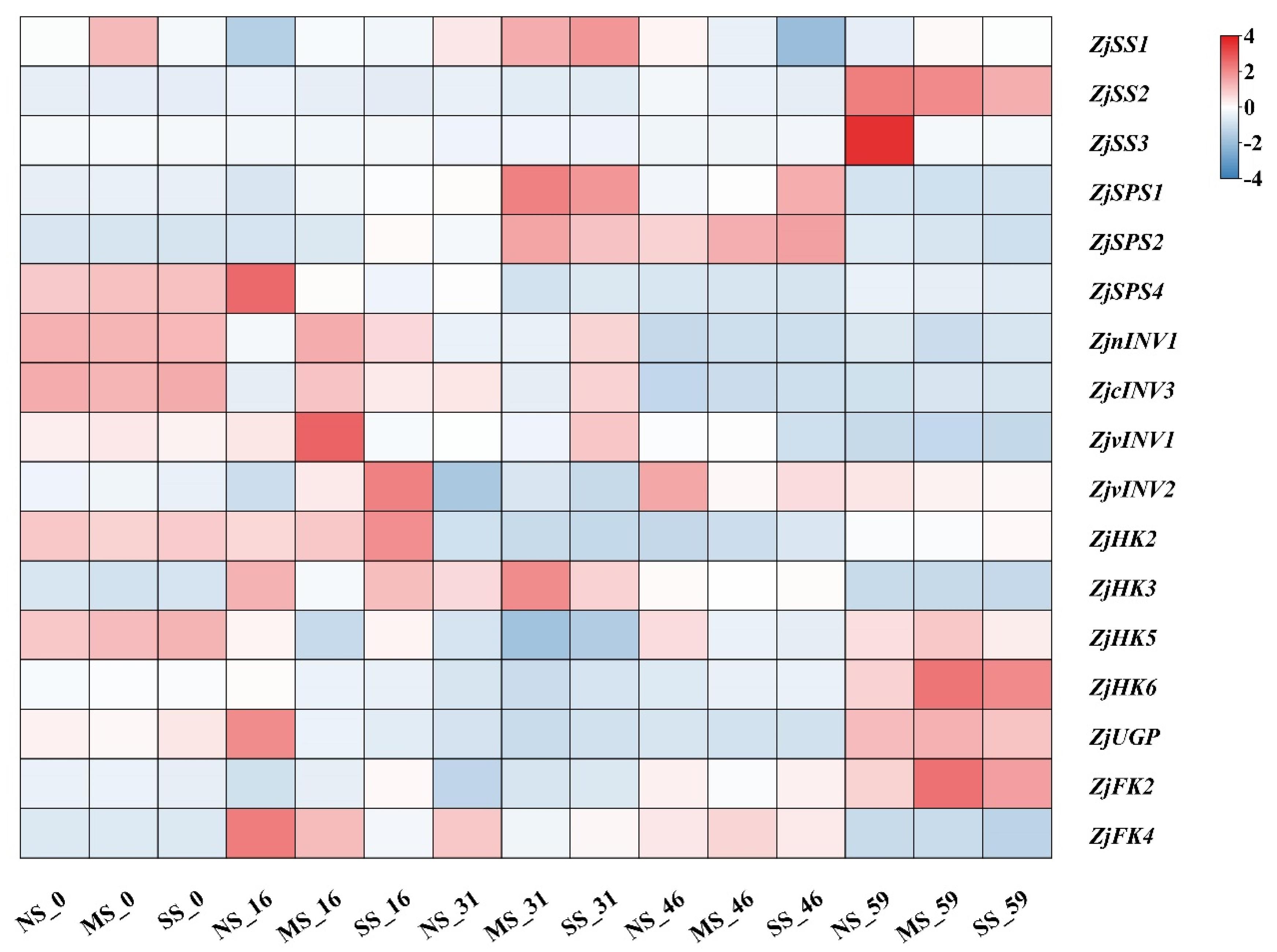
3.4. Expression of Genes Involved in Sugar Metabolism in Leaves
There were various different expression patterns of genes related to sucrose and hexose metabolism in jujube leaves under shading conditions (Figure 4). Compared with NS, the expression of ZjSPS1 and ZjSPS2 in MS leaves was up-regulated at 16 d, 31 d, and 46 d but down-regulated at 59 d. The expression of ZjnINV1, ZjcINV3, and ZjvINV2 were up-regulated after shading. The expression levels of ZjSS2 and ZjSS3 in shaded leaves were low at early development and increased sharply at 59 d, while shading inhibited the up-regulation, especially in fruits under SS conditions. Compared with NS, the expression levels of ZjHK2, ZjHK3, and ZjHK6 were up-regulated in shaded leaves, while ZjHK5 was down-regulated. ZjFK2 expression in shaded leaves was higher than that in NS, while ZjFK4 expression was not significantly different. Compared with NS, ZjUGP expression in shaded leaves was significantly down-regulated at 16 d, 31 d, and 46 d.
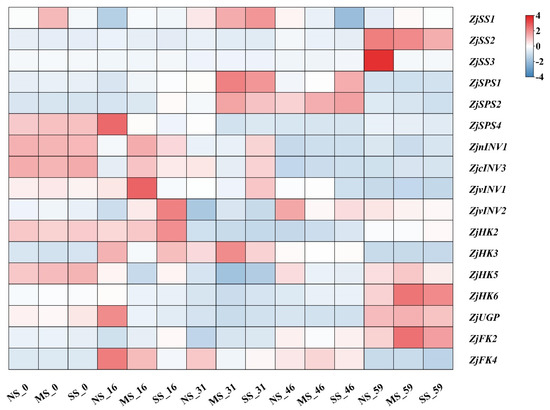
Figure 4.
Heatmap analyses of gene expression involved in sugar metabolism in leaves. Sucrose synthase (SS), sucrose phosphosynthase (SPS), invertase (INV), fructokinase (FK), pyrophosphorylase (UGP), hexokinase (HK). Colors from blue to red indicate that the levels of gene expression are low to high, and a redder color indicates that the expression levels of a gene are high. The genes expression in the fruit of NS-0 were all standardized to one. The expression levels of genes were compared by log2 fold change and averaged, and TBtools were used for heatmap analyses. The same is below.
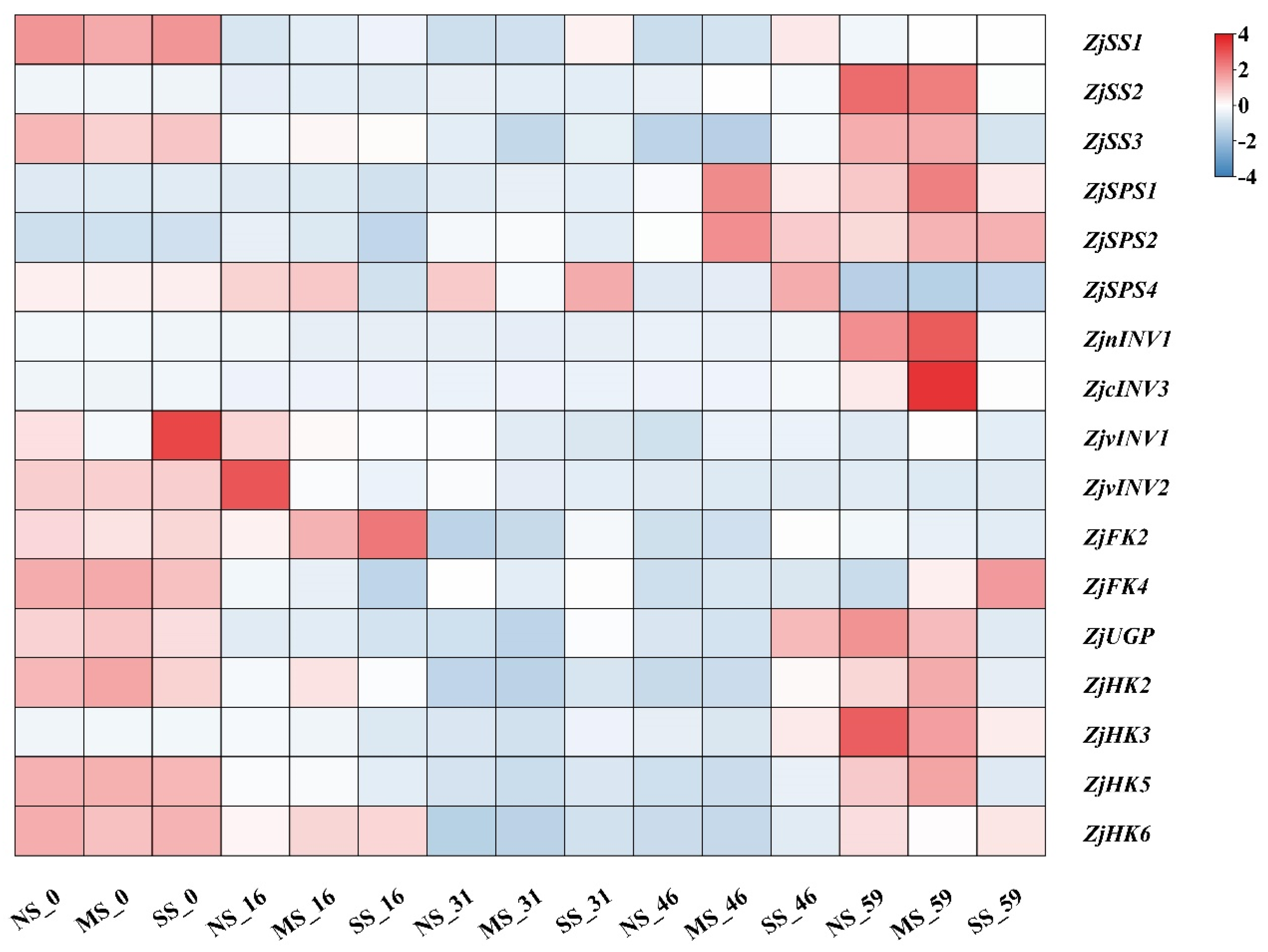
3.5. Expression of Genes Involved in Sugar Metabolism in Fruits
Genes involved in sugar metabolism were differentially expressed during fruit development and with shading (Figure 5). The expression level of ZjSS2 was lower in the early development stage and sharply up-regulated at the mature stage, and ZjSS3 expression decreased first and then increased during the development. ZjSS2 and ZjSS3 expression of fruits in SS were significantly lower than fruits in NS. The ZjSPS1 and ZjSPS2 expression of fruits in MS were down-regulated and then up-regulated, and in SS was down-regulated in comparison with NS. Compared with NS, the expression of ZjnINV1 and ZjcINV3 in fruits of MS at 59 d were up-regulated but were down-regulated in fruits grown under SS. The ZjvINV2 expression at the early stage was higher than in the later stage, and the expression in shaded fruits was decreased compared with NS. The expression levels of ZjHK2, ZjHK6, ZjFK4, and ZjUGP decreased at first and then increased during fruit development under shading conditions. ZjHK3 expression in three treatments peaked at 59 d and was down-regulated in shaded fruits. The tendency of ZjHK6 expression was decreased-increased during fruit development. The ZjFK2 expression was higher at the early stage than in the later stage, and the expression in SS fruits was significantly up-regulated at 16 d, 31 d, and 46 d compared with NS. ZjUGP expression of shaded fruits at 59 d was down-regulated compared with NS.
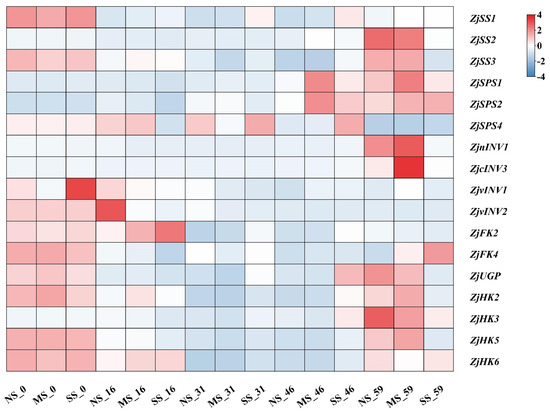
Figure 5.
Heatmap analyses of gene expression involved in sugar metabolism in fruits. Sucrose synthase (SS), sucrose phosphosynthase (SPS), invertase (INV), fructokinase (FK), pyrophosphorylase (UGP), hexokinase (HK).
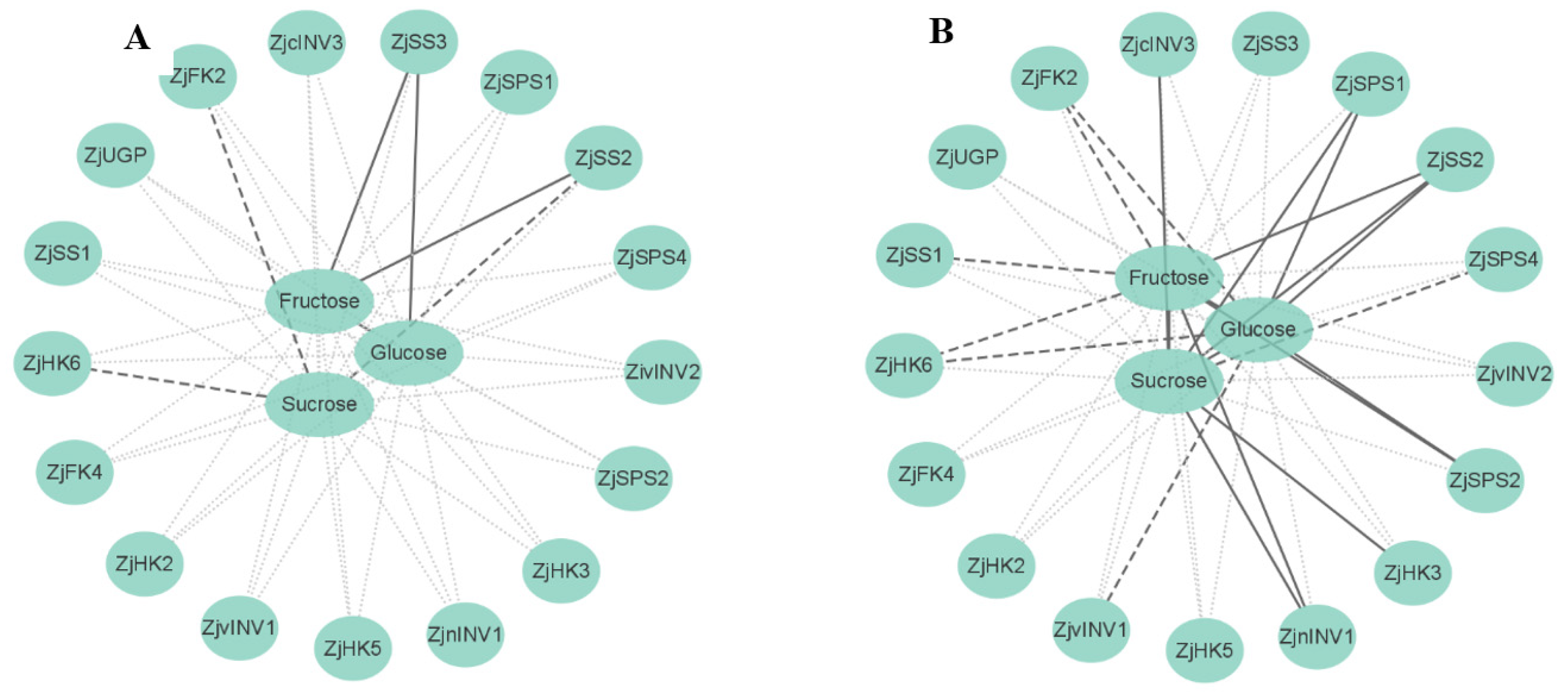
3.6. Correlation Analysis of Metabolite Contents and Gene Expression
The correlations between component contents and related gene expression were diverse under shading conditions (Figure 6). The expression levels of ZjSS2, ZjHK6, and ZjFK2 in leaves were negatively correlated with sucrose. Fructose content in leaves was positively correlated with the expression levels of ZjSS2 and ZjSS3, and glucose content was positively correlated with ZjSS3 expression.
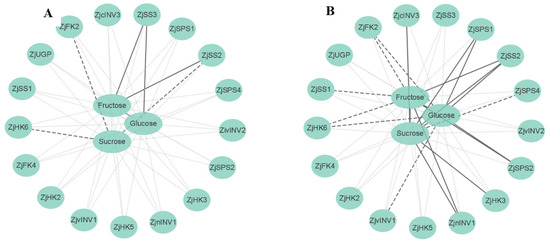
Figure 6.
Correlations between metabolite contents and gene expression. (A) Correlation analysis of sugar metabolism in leaves. (B) Correlation analysis of sugar metabolism in fruits. Sucrose synthase (SS), sucrose phosphosynthase (SPS), invertase (INV), fructokinase (FK), pyrophosphorylase (UGP), hexokinase (HK). Gray dashed lines between indices represent no correlations, whereas black dashed and solid black lines represent negative and positive correlations at significant levels (p < 0.05), respectively. Correlations analysis was drawn using Cytoscape 3.6.1.
The sucrose, fructose, and glucose contents in fruits were positively correlated with each other. Sucrose content was positively correlated with ZjSS2, ZjSPS1, ZjnINV1, ZjcINV3, and ZjHK3 expression but negatively correlated with ZjSPS4 expression. Fructose content was positively correlated with the expression of ZjSS2, ZjSPS2, and ZjnINV1, while negatively correlated with ZjSS1, ZjHK6, and ZjFK2. The glucose content was positively correlated with ZjSS2, ZjSPS1, and ZjSPS2 expression, while negatively correlated with ZjvINV1, ZjHK6, and ZjFK2.
4. Discussion
4.1. Shading Increased Photosynthetic Pigment Contents of Leaves but Decreased Specific Leaf Weight
Under shading conditions, plants might maintain the original photosynthetic rate by changing photosynthetic pigment contents and chl a/b ratio or adapt to low light by reducing the photosynthetic rate [3,28]. In this study, compared with no shading, the chlorophyll and carotenoid contents in shaded leaves increased significantly at the later stage, while the chl a/b ratio decreased, which was consistent with the reports of strawberry and rice [2,29]. The results indicated that shaded jujube leaves obtained higher light energy by increasing photosynthetic pigment contents and meanwhile improved photosynthetic phosphorylation activity by reducing the chl a/b ratio [30,31]. However, the decrease in specific leaf weight in jujube leaves indicated that the enhanced capacity for light capture did not compensate for the reduction in leaf photosynthesis caused by shading, which was similar to results from studies of black currant, grape, and cigar tobacco leaves [32,33,34].
4.2. Shading Inhibited Sugar Accumulation in Leaves
Sugars are the primary products of photosynthetic carbon assimilation in leaves [14]. The growth-promoting effects of sucrose in leaf blade and petiole of Arabidopsis thaliana were highly dependent on light conditions [35]. In this study, the sucrose, fructose, and glucose contents of leaves were negatively correlated with the extent of shading, which was consistent with the changes in specific leaf weight. The decrease in sucrose content after shading was more obvious than for fructose and glucose, which was consistent with reports for nectarine and wax apple [36,37]. Results of strawberry [2], maize [34,38], and soybean [39] showed that, under low light conditions, the decrease in the photosynthetic rate of leaves led to a weakening of the carbon assimilation capacity, thus seriously affecting sugar synthesis [40]. Therefore, the suppression of sucrose synthesis caused by the decreased photosynthetic rate was the main reason underlying the inhibition of the sugar accumulation in jujube leaves under shade conditions; the reduced carbohydrate accumulation then repressed vegetative growth [37].
The activities of SPS and SS-s (synthesis direction) in melon leaves are reduced significantly under low light stress, resulting in decreased sucrose content [41]. Similarly, SuS showed reduced specific activity in nectarine leaves under low light, resulting in lower sucrose concentration [37]. In the present study, we suggested that the lower expression of ZjSS2 and ZjSS3 in shaded leaves jointly inhibited sucrose synthesis and thus reduced sugar accumulation, which was similar to the results of previous studies in cotton [42]. At the same time, the up-regulation of ZjINV expression under shading accelerated sucrose hydrolysis, while the up-regulation of ZjHK6 and ZjFK2 promoted irreversible phosphorylation of hexose, thus inhibiting the accumulation of fructose and glucose. Similar results were seen in grapevine, where lower starch and soluble sugars in shaded leaves than in exposed leaves were related to higher expression of hexose transporters and invertases [43].
4.3. Shading Inhibited Sugar Accumulation in Fruits
Sugar is transported through the phloem from leaf (source) to fruit (sink) for metabolism and storage [44,45]. Compared with the control, the contents of sucrose and reducing sugars in fruits of strawberry, apple, and nectarine were significantly reduced under low light [4,5,18,46,47]. In this study, sucrose was the major sugar component, which was similar to other cultivars of jujube [27]. The contents of sucrose decreased at the late stage of fruit development with increased extent shading, which was also seen in sugarcane [48]. The decreased sugar content in fruits under shading would be caused by inhibition of the accumulation of primary and secondary metabolites, especially carbohydrates [3]. However, the shading did not affect the sugar concentration in the fruits of the tomato [49] and grape [11]. It is speculated that the main reason for this difference is that various species of fruits show different changes in sugar metabolism in response to low light.
With shading during the later stage of melon fruit development, sucrose accumulation was inhibited by decreased activities of SPS and SS-s [41]. In the present study, under a shading environment, the low sucrose contents in fruits were associated with down-regulated expression of ZjSS2 and ZjSPS1 in the early period suppressing sucrose synthesis and up-regulated expression of ZjnINV1 and ZjcINV3 in late-stage increased sucrose degradation. In contrast, high expression of CaSUS2 enhanced SUS activity but did not increase sucrose content in coffee fruits in shaded conditions, and it was speculated that sucrose metabolism might be redirected via other pathways [50]. Similar to leaf, up-regulated expression of ZjHK6 and ZjFK2 would promote hexose phosphorylation, inhibiting the accumulation of fructose and glucose in shaded fruits. Huang et al. [51] believed that SPS and INV should be considered key enzymes that promote sugar accumulation in the jujube cultivar ‘Junzao,’ which is different from the findings in our study under low light conditions. This difference might be attributed to altered transcription levels of related genes that affect sugar accumulation, or it might be the result of cultivar differences.
4.4. Shading May Affected Sugar Transport from Leaf to Fruit
Sucrose, as the main form of photosynthate transport from source to sink, is also a signal molecule that coordinates the source-sink relationship of plants [14]. The synthesis and transport of photosynthates to the sink might reduce under low light, which would not be conducive to the growth of the sink tissues and the accumulation of assimilates [18]. Roopendra et al. [48] showed that constant removal of sugars from the site of synthesis, in response to elevated sink demand, perhaps prevents suppression of sugar accumulation from photosynthesis. Therefore, our results implied that the decrease in sucrose concentration in jujube fruit with shading might be the result of lower sucrose import from the leaf, which was similar to cotton fiber [42].
5. Conclusions
Compared with no shading, photosynthetic pigment content in shaded leaves was increased, while specific leaf weight was decreased. Shading inhibited sugar accumulation mainly by reducing sucrose content in leaves and fruit. Under shading conditions, low expression of ZjSS2 and ZjSS3 reduced sugar content in leaf, and down-regulation expression of ZjSS2 and ZjSPS1 inhibited sugar accumulation in fruit. The decrease in sugar content in shaded leaves and fruit implied that facility cultivation in southern production areas negatively affected the growth and fruit quality of jujube. Hence, we suggested that high transmittance materials and artificial light supplements could be appropriately selected to compensate for the resulting loss of light.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8070592/s1, Table S1: Primers for qRT-PCR analysis.
Author Contributions
Y.W. drafted and edited the manuscript. S.R. performed the experiments. X.L. (Xuelian Li) analyzed the data. X.L. (Xian Luo) revised the manuscript. Q.D. was the project investigator and research supervisor. The published version of this manuscript was revised and agreed upon by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Foundation for Disciplines Construction and Development of Sichuan Agricultural University (No: 035/20211993021); the Key Science and Technology Research Project of Deyang (No: 2018CKJ018); the Special Action Plan Project of Science and Technology-Rich People and Strong County of Sichuan Province “Integration and Demonstration of Key Technologies for Industrialized Development of Ziziphus jujuba ‘Zhanshangmizao’ of Santai County” (2012–2014) (No: 06320200).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kishore, K.; Rupa, T.R.; Samant, D. Influence of shade intensity on growth, biomass allocation, yield and quality of pineapple in mango-based intercropping system. Sci. Hortic. 2021, 278, 109869. [Google Scholar] [CrossRef]
- Choi, H.G. Correlation among phenotypic parameters related to the growth and photosynthesis of strawberry (Fragaria × ananassa Duch.) grown under various light intensity conditions. Front. Plant Sci. 2021, 12, 647585. [Google Scholar] [CrossRef] [PubMed]
- Mditshwa, A.; Magwaza, L.S.; Tesfay, S.Z. Shade netting on subtropical fruit: Effect on environmental conditions, tree physiology and fruit quality. Sci. Hortic. 2019, 256, 108556. [Google Scholar] [CrossRef]
- Palmer, J.; Diack, R.; Johnston, J.; Boldingh, H. Manipulation of fruit dry matter accumulation and fruit size in ‘Scifresh’ apple through alteration of the carbon supply, and its relationship with apoplastic sugar composition. J. Hortic. Sci. Bitechnol. 2013, 88, 483–489. [Google Scholar] [CrossRef]
- Jing, C.J.; Feng, D.P.; Zhao, Z.Y.; Wu, X.H.; Chen, X.F. Effect of environmental factors on skin pigmentation and taste in three apple cultivars. Acta Physiol. Plant. 2020, 42, 69. [Google Scholar] [CrossRef]
- Li, M.J.; Ma, F.W.; Liu, J.; Li, J. Shading the whole vines during young fruit development decreases ascorbate accumulation in kiwi. Physiol. Plant. 2010, 140, 225–237. [Google Scholar] [CrossRef]
- Peavey, M.; Goodwin, I.; Mcclymont, L.; Chandra, S. Effect of shading on red colour and fruit quality in blush pears “Anp-0118” and “Anp-0131”. Plant 2020, 9, 206. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Q.; Wang, Y.; Huang, X.M.; Li, J.; Wang, H.; Li, J.G. De novo assembly and characterization of fruit transcriptome in Litchi chinensis Sonn and analysis of differentially regulated genes in fruit in response to shading. BMC Genom. 2013, 14, 552. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Dardick, C.D.; Beers, E.; Callanhan, A.M.; Xia, R.; Yuan, R.C. Transcriptomics of shading-induced and NAA-induced abscission in apple (Malus domestica) reveals a shared pathway involving reduced photosynthesis, alterations in carbohydrate transport and signaling and hormone crosstalk. BMC Plant Biol. 2011, 11, 138. [Google Scholar] [CrossRef] [Green Version]
- Jutamanee, K.; Onnom, S. Improving photosynthetic performance and some fruit quality traits in mango trees by shading. Photosynthetica 2016, 54, 542–550. [Google Scholar] [CrossRef]
- Abeysinghe, S.K.; Greer, D.H.; Rogiers, S.Y. The effect of light intensity and temperature on berry growth and sugar accumulation in Vitis vinifera ‘Shiraz’ under vineyard conditions. Vitis 2019, 58, 7–16. [Google Scholar]
- Milenkovic, L.; Mastilovic, J.; Kevresan, Z.; Bajic, A.; Gledic, A.; Stanojevic, L.; Cvetkovic, D.; Sunic, L.; Ilic, Z.S. Effect of shading and grafting on yield and quality of tomato. J. Sci. Food Agric. 2019, 100, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Li, X.B.; Li, C.N.; Sun, J.; Jackson, A. Dynamic changes of enzymes involved in sugar and organic acid level modification during blueberry fruit maturation. Food Chem. 2020, 309, 125617. [Google Scholar] [CrossRef]
- Durán-Soria, S.; Pott, D.M.; Osorio, S.; Vallarino, J.G. Sugar signaling during fruit ripening. Front. Plant Sci. 2020, 11, 564917. [Google Scholar] [CrossRef]
- Xu, Q.Y.; Liesche, J. Sugar export from Arabidopsis leaves actors and regulatory strategies. J. Exp. Bot. 2021, 72, 5275–5284. [Google Scholar] [CrossRef] [PubMed]
- Manja, K.; Aoun, M. The use of nets for tree fruit crops and their impact on the production: A review. Sci. Hortic. 2019, 246, 110–122. [Google Scholar] [CrossRef]
- Abreu, A.C.; Marín, P.; Aguilera-Sáez, L.M.; Tristán, A.I.; Peña, A.; Oliveira, I.; Simões, M.; Valera, D.; Fernández, I. Effect of a shading mesh on the metabolic, nutritional, and defense profiles of harvested greenhouse-grown organic tomato fruits and leaves revealed by NMR metabolomics. J. Agric. Food Chem. 2019, 67, 12972–12985. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J.; Kwon, J.K.; Bekhzod, K.; Park, K.S.; Lee, S.Y. Yield loss and quality degradation of strawberry fruits cultivated under the deficient insolation conditions by shading. Hortic. Environ. Biotechnol. 2014, 55, 263–270. [Google Scholar] [CrossRef]
- Meena, V.S.; Kashyap, P.; Nangare, D.D.; Singh, J. Effect of coloured shade nets on yield and quality of pomegranate (Punica granatum) cv. Mridula in semi-arid region of Punjab. Indian J. Agric. Sci. 2016, 86, 500–505. [Google Scholar]
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar] [CrossRef]
- Solomakhin, A.; Blanke, M.M. Can coloured hailnets improve taste (sugar, sugar: Acid ratio), consumer appeal (colouration) and nutritional value (anthocyanin, vitamin C) of apple fruit? LWT-Food Sci. Technol. 2010, 43, 1277–1284. [Google Scholar] [CrossRef]
- Liu, M.J.; Wang, J.; Wang, L.L.; Liu, P.; Zhao, J.; Zhao, Z.H.; Yao, S.R.; Stănică, F.; Liu, Z.G.; Wang, L.X.; et al. The historical and current research progress on jujube–a superfruit for the future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.H.; Wang, G.X.; Liang, L.S. Development and characterization of SSR markers in Chinese jujube (Ziziphus jujuba Mill.) and its related species. Sci. Hortic. 2011, 129, 597–602. [Google Scholar] [CrossRef]
- Jiang, W.Q.; Li, N.; Zhang, D.; Meinhardt, L.; Cao, B.; Li, Y.J.; Song, L.H. Elevated temperature and drought stress significantly affect fruit quality and activity of anthocyanin-related enzymes in jujube (Ziziphus jujuba Mill. cv. ‘Lingwuchangzao’). PLoS ONE 2020, 15, e0241491. [Google Scholar] [CrossRef]
- Ren, S.; Wangy, Y.; Tang, Y.; Zhou, Q.; Deng, Q.X. Effect of shade treatment on active oxygen metabolism and fruit quality of Zizyphus jujuba fruit. Acta Agric. Boreali-Occident. Sin. 2020, 29, 709–717. [Google Scholar]
- Wu, X.Y.; Khan, R.; Gao, H.J.; Liu, H.B.; Zhang, J.; Ma, X.H. Low light alters the photosynthesis process in cigar tobacco via modulation of the chlorophyll content, chlorophyll fluorescence, and gene expression. Agriculture 2021, 11, 755. [Google Scholar] [CrossRef]
- Gao, Q.H.; Wu, C.S.; Yu, J.G.; Wang, M.; Ma, Y.J.; Li, C.L. Textural characteristic, antioxidant activity, sugar, organic acid, and phenolic profiles of 10 promising jujube (Ziziphus jujuba Mill.) selections. J. Food Sci. 2012, 77, 1218–1225. [Google Scholar] [CrossRef]
- Atanasova, L.; Stefanov, D.; Yordanov, I.; Kornova, K.; Kavardzikov, L. Comparative characteristics of growth and photosynthesis of sun and shade leaves from normal and pendulum walnut (Juglans regia L.) trees. Photosynthetica 2003, 41, 289–292. [Google Scholar] [CrossRef]
- Sekhar, S.; Panda, D.; Kumar, J.; Biswal, M.; Baig, M.J.; Kumar, A.; Umakanta, N.; Sangamitra, S.; Pradhan, S.K.; Shaw, B.P.; et al. Comparative transcriptome profiling of low light tolerant and sensitive rice varieties induced by low light stress at active tillering stage. Sci. Rep. 2019, 9, 5753. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, W.J.; Yu, H.J. Effects of exogenous epibrassinolide on photosynthetic characteristics in tomato (Lycopersicon esculentum Mill.) seedlings under weak light stress. J. Agric. Food Chem. 2010, 58, 3642–3645. [Google Scholar] [CrossRef]
- Yu, H.J.; Zhao, W.C.; Wang, M.; Yang, X.Y.; Jiang, W.J. The exogenous application of spermidine alleviates photosynthetic inhibition and membrane lipid peroxidation under low-light stress in tomato (Lycopersicon esculentum Mill.) seedlings. Plant Growth Regul. 2015, 78, 413–420. [Google Scholar] [CrossRef]
- Qiu, T.; Wu, Y.Y.; Shen, Z.L.; Wu, Y.Y.; Lu, D.; He, J.W. Effects of shading on leaf physiology and morphology in the ‘Yinhong’ grape plants. Rev. Bras. De Frutic. 2018, 40, 37. [Google Scholar]
- Wolske, E.T.; Branham, B.E. Growth and productivity of ‘consort’ black currant grown under varying levels of artificial shade. Hortscience 2021, 56, 3–7. [Google Scholar] [CrossRef]
- Wu, H.Y.; Tang, H.K.; Liu, L.A.; Shi, L.; Zhang, W.F.; Jiang, C.D. Local weak light induces the improvement of photosynthesis in adjacent illuminated leaves in maize seedlings. Physiol. Plant. 2020, 171, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Kozuka, T.; Horiguchi, G.; Kim, G.T.; Ohgishi, M.; Sakai, T.; Tsukaya, A.H. The different growth responses of the Arabidopsis thaliana leaf blade and the petiole during shade avoidance are regulated by photoreceptors and sugar. Plant Cell Physiol. 2005, 46, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Jian, Y.; Wu, G.; Zhou, D.; Hu, Z.; Quan, Z.; Zhou, B. Effects of shading on carbohydrates of Syzygium samarangense. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Huang, W.D.; Zhan, J.C. Effect of low light on the activity of sucrose synthase in leaves of nectarine. J. Hortic. Sci. Biotechnol. 2005, 80, 358–362. [Google Scholar]
- Shen, S.; Li, B.B.; Deng, T.; Xiao, Z.D.; Chen, X.M.; Hu, H.; Zhang, B.C.; Wu, G.; Li, F.; Zhao, X.; et al. The equilibrium between sugars and ethylene is involved in shading and drought-induced kernel abortion in maize. Plant Growth Regul. 2020, 91, 101–111. [Google Scholar] [CrossRef]
- Hussain, S.; Mumtaz, M.; Manzoor, S.; Shuxian, L.; Ahmed, I.; Skalicky, M.; Brestic, M.; Rastogi, A.; Ulhassan, Z.; Shafiq, I.; et al. Foliar application of silicon improves growth of soybean by enhancing carbon metabolism under shading conditions. Plant Physiol. Bioch. 2021, 159, 43–52. [Google Scholar] [CrossRef]
- Wang, Q.M.; Hou, F.Y.; Dong, S.X.; Xie, B.T.; Li, A.X.; Zhang, H.Y.; Zhang, L.M. Effects of shading on the photosynthetic capacity, endogenous hormones and root yield in purple-fleshed sweet potato (Ipomoea batatas (L.) Lam). Plant Growth Regul. 2014, 72, 113–122. [Google Scholar] [CrossRef]
- Yang, L.Y.; Chen, J.J.; Sun, X.M.; Li, J.X.; Chen, N.L. Inhibition of sucrose and galactosyl-sucrose oligosaccharide metabolism in leaves and fruits of melon (Cucumis melo L.) under low light stress. Sci. Hortic. 2019, 244, 343–351. [Google Scholar] [CrossRef]
- Hu, W.; Ma, Y.N.; Lv, F.J.; Liu, J.R.; Zhao, W.Q.; Chen, B.L.; Meng, Y.L.; Wang, Y.H.; Zhou, Z.G. Effects of late planting and shading on sucrose metabolism in cotton fiber. Environ. Exp. Bot. 2016, 131, 161–172. [Google Scholar] [CrossRef]
- Dayer, S.; Murcia, G.; Prieto, J.A.; Durán, M.; Martínez, L.; Píccoli, P.; Peña, J.P. Non-structural carbohydrates and sugar export in grapevine leaves exposed to different light regimes. Physiol. Plantarum 2020, 171, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Lecourieux, F.; Kappel, C.; Lecourieux, D.; Serrano, A.; Torres, E.; Arce-Johnson, P.; Delrot, S. An update on sugar transport and signalling in grapevine. J. Exp. Bot. 2014, 65, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.; Cho, L.H.; Tun, W.; Jeon, J.S.; An, G. Sucrose signaling in higher plants. Plant Sci. 2021, 302, 110703. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Huang, W.D.; Zhan, J.C. Effects of low light on phloem ultrastructure and subcellular localization of sucrose synthase in Prunus persica L. var. nectarina Ait. fruit. Russ. J. Plant Physiol. 2009, 56, 462–469. [Google Scholar] [CrossRef]
- Watson, R.; Wright, C.J.; Mcburney, T.; Taylor, A.J.; Linforth, R.S.T. Influence of harvest date and light integral on the development of strawberry flavour compounds. J. Exp. Bot. 2002, 53, 2121–2129. [Google Scholar] [CrossRef]
- Roopendra, K.; Chandra, A.; Saxena, S. Increase in sink demand in response to perturbed source-sink communication by partial shading in sugarcane. Sugar Tech 2019, 21, 672–677. [Google Scholar] [CrossRef]
- Hernández, V.; Hellín, P.; Fenoll, J.; Flores, P. Interaction of nitrogen and shading on tomato yield and quality. Sci. Hortic. 2019, 255, 255–259. [Google Scholar] [CrossRef]
- Geromel, C.; Ferreira, L.C.P.; Davrieux, F.; Guyot, B.; Ribeyre, F.; Scholz, M.B.G.D.; Pereira, L.F.P.; Vaast, P.; Pot, D.; Leroy, T.; et al. Effects of shade on the development and sugar metabolism of coffee (Coffea arabica L.) fruits. Plant Physiol. Bioch. 2008, 46, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Chen, X.; He, A.B.; Ma, Z.B.; Gong, T.Q.; Xu, K.N.; Chen, R.H. Integrative morphological, physiological, proteomics analyses of jujube fruit development provide insights into fruit quality domestication from wild jujube to cultivated jujube. Front. Plant Sci. 2021, 12, 773825. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).