
Evolution of Phytochemical Variation in Myrtle (Myrtus communis L.) Organs during Different Phenological Stages




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. EO Isolation
2.3. EO Analysis
2.4. Preparation of Extracts
2.5. Determination of Total Phenolic Content
2.6. Determination of Total Flavonoid Content
2.7. Antioxidant Activity (DPPH Radical Scavenging Assay)
2.8. Statistical Analysis
3. Results and Discussion
3.1. EO Content
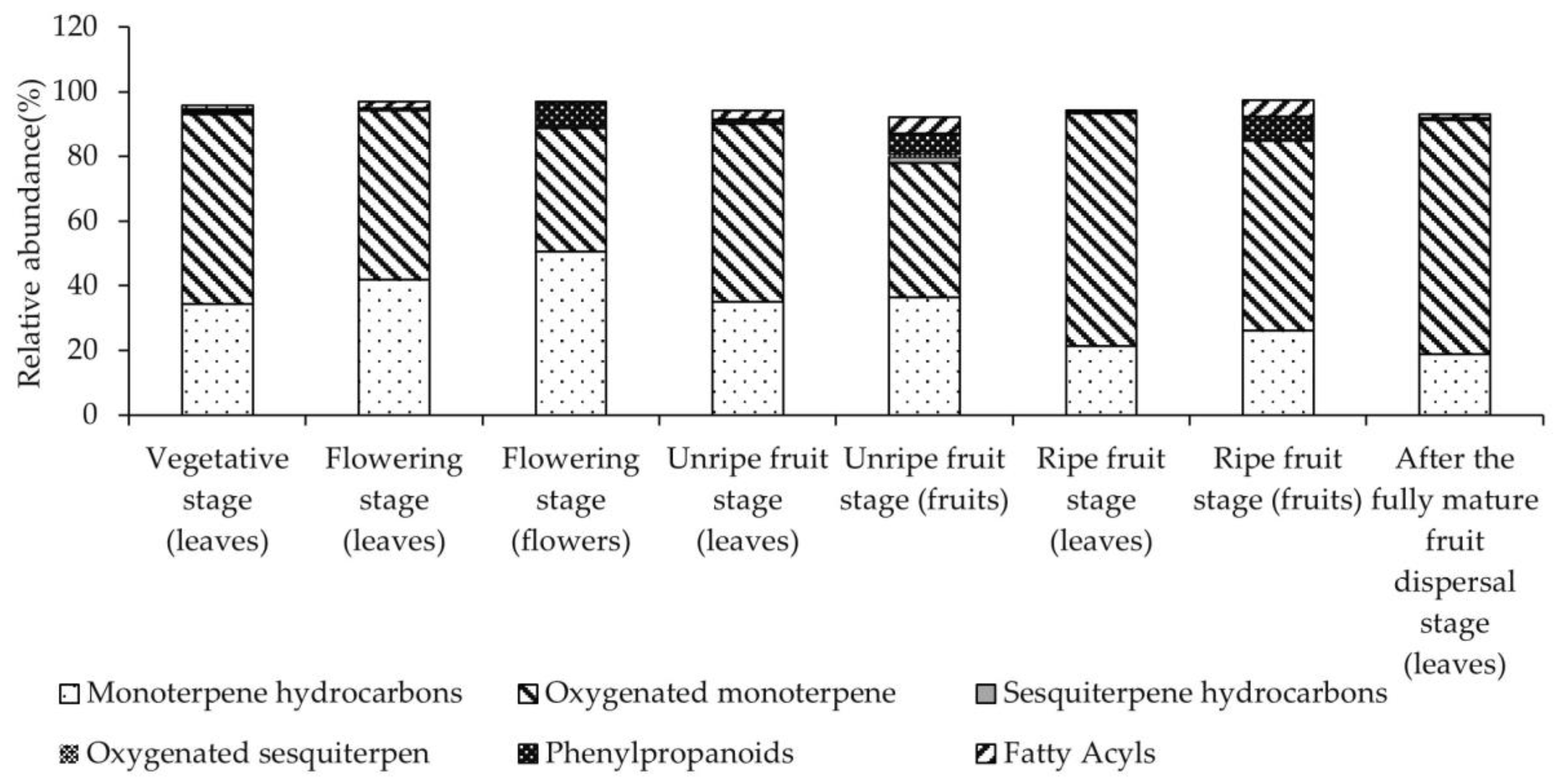
3.2. EO Composition
3.3. Antioxidant Activity, Total Phenol and Flavonoid Contents
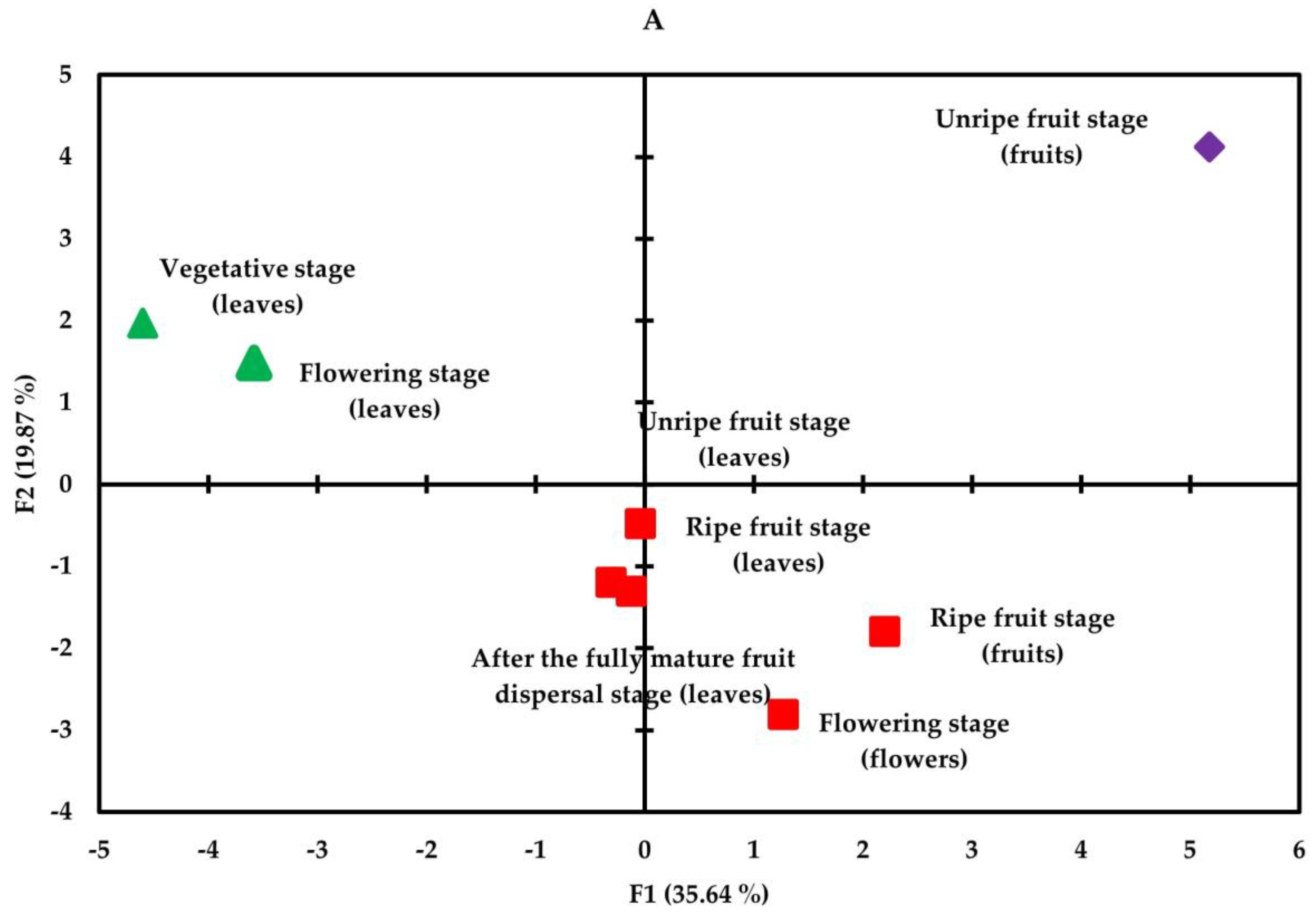
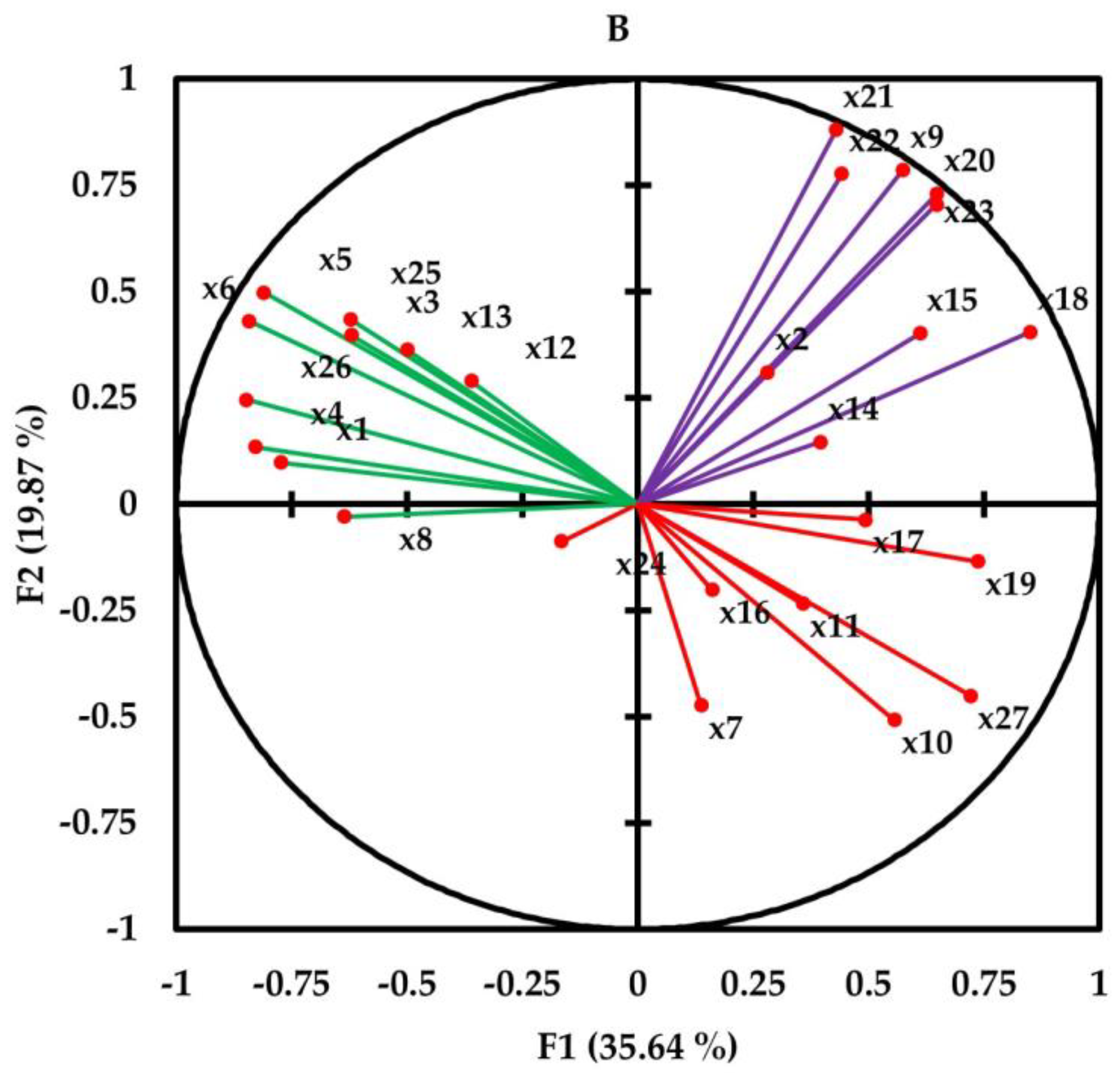
3.4. Correlation and Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahimmalek, M.; Mirzakhani, M.; Pirbalouti, A.G. Essential oil variation among 21 wild myrtle (Myrtus communis L.) populations collected from different geographical regions in Iran. Ind. Crops Prod. 2013, 51, 328–333. [Google Scholar] [CrossRef]
- Pezhmanmehr, M.; Dastan, D.; Ebrahimi, S.N.; Hadian, J. Essential oil constituents of leaves and fruits of Myrtus communis L. from Iran. J. Essent. Oil-Bear. Plants. 2010, 13, 123–129. [Google Scholar] [CrossRef]
- Anwar, S.; Ahmed, N.; Al Awwad, N.; Ansari, S.Y.; Wagih, M.E. Myrtle (Myrtus communis L.) oils. In Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2016; pp. 581–592. [Google Scholar]
- Omezzine, F.; Ladhari, A.; Haouala, R. Physiological and biochemical mechanisms of allelochemicals in aqueous extracts of diploid and mixoploid Trigonella foenum-graecum L. S. Afr. J. Bot. 2014, 93, 167–178. [Google Scholar] [CrossRef]
- Sumbul, S.; Ahmad, M.A.; Asif, M.; Saud, I.; Akhtar, M. Evaluation of Myrtus communis Linn. berries (common myrtle) in experimental ulcer models in rats. Hum. Exp. Toxicol. 2010, 29, 935–944. [Google Scholar] [CrossRef]
- Raeiszadeh, M.; Pardakhty, A.; Sharififar, F.; Farsinejad, A.; Mehrabani, M.; Hosseini-Nave, H.; Mehrabani, M. Development, physicochemical characterization, and antimicrobial evaluation of niosomal myrtle essential oil. Int. J. Pharm. Sci. Rev. Res. 2018, 13, 250. [Google Scholar] [CrossRef]
- Zam, W.; Ali, A. Evaluation of mechanical, antioxidant and antimicrobial properties of edible film containing myrtle berries extract. Nat. Prod. J. 2018, 8, 323–330. [Google Scholar] [CrossRef]
- Aleksic, V.; Knezevic, P. Antimicrobial and antioxidative activity of extracts and essential oils of Myrtus communis L. Microbiol. Res. 2014, 169, 240–254. [Google Scholar] [CrossRef]
- Aydın, C.; Özcan, M.M. Determination of nutritional and physical properties of myrtle (Myrtus communis L.) fruits growing wild in Turkey. J. Food Eng. 2017, 79, 453–458. [Google Scholar] [CrossRef]
- Bajalan, I.; Ghasemi Pirbalouti, A.G. Variation in antibacterial activity and chemical compositions of essential oil from different populations of myrtle. Ind. Crops Prod. 2014, 61, 303–307. [Google Scholar] [CrossRef]
- Zolfaghari, P.; Salmanian, A.; Riazi, G. Clinical trial of efficacy of myrtle oil in the treatment of Herpes simplex. Iran. J. Med. Sci. 1997, 22, 137–139. [Google Scholar]
- Alamanni, M.C.; Cossu, M. Radical scavenging activity and antioxidant activity of liquors of myrtle (Myrtus communis L.) berries and leaves. Ital. J. Food Sci. 2004, 16, 197–208. [Google Scholar]
- Deriu, A.; Branca, G.; Molicotti, P.; Pintore, G.; Chessa, M.; Tirillini, B.; Paglietti, B.; Mura, A.; Sechi, L.A.; Fadda, G.; et al. In vitro activity of essentialoil of Myrtus communis L. against Helicobacter pylori. Int. J. Antimicrob. Agents. 2017, 30, 562–563. [Google Scholar] [CrossRef] [PubMed]
- Cannas, S.; Molicotti, P.; Usai, D.; Maxia, A.; Zanetti, S. Antifungal, anti-biofilm and adhesion activity of the essential oil of Myrtus communis L. against Candida species. Nat. Prod. Res. 2014, 28, 2173–2177. [Google Scholar] [CrossRef] [PubMed]
- Tretiakova, I.; Blaesius, D.; Maxia, L.; Wesselborg, S.; Schulze-Osthoff, K.; Cinalt, J., Jr.; Michaelis, M.; Werz, O. Myrtucommulone from Myrtus communis L. indices apoptosis in cancer cells via the mitochondrial pathway involvingcaspase-9. Apoptosis 2008, 13, 119–131. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; Taoubi, K.; El-Haj, S.; Bessiere, J.M.; Rammal, S. Insecticidal properties of essential plant oils against the mosquito Culex pipiens molestus (Diptera: Culicidae). Pest. Manag. Sci. 2002, 58, 491–495. [Google Scholar] [CrossRef]
- Mimica-Dukić, N.; Bugarin, D.; Grbović, S.; Mitić-Ćulafić, D.; Vuković-Gačić, B.; Orčić, D.; Couladis, M. Essential oil of Myrtus communis L. as a potential antioxidant and antimutagenic agents. Molecules 2010, 15, 2759–2770. [Google Scholar] [CrossRef] [PubMed]
- Hayder, N.; Bouhlel, I.; Skandrani, I.; Kadri, M.; Steiman, R.; Guiraud, P.; Mariotte, A.M.; Ghedira, K.; Dijoux-Franca, M.G.; Chekir-Ghedira, L. In vitro antioxidant and antigenotoxic potentials of myricetin-3-O-galactoside and myricetin-3-O-rhamnoside from Myrtus communis: Modulation of expression of genes involved in cell defence system using cDNA microarray. Toxicol. Vitr. 2008, 22, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Elfellah, M.S.; Akhter, M.H.; Khan, M.T. Anti-hyperglycaemic effect of an extract of Myrtuscommunis in streptozotocin induced diabetes in mice. J. Ethnopharmacol. 1984, 11, 275–281. [Google Scholar] [CrossRef]
- Aidi Wannes, W.A.; Mhamdi, B.; Sriti, J.; Jemia, M.B.; Ouchikh, O.; Hamdaoui, G.; Marzouk, B. Antioxidant activities of the essential oils and methanol extracts from myrtle (Myrtus communis var. italica L.) leaf, stem and flower. Food Chem. Toxicol. 2010, 48, 1362–1370. [Google Scholar] [CrossRef]
- Kanoun, K.; Belyagoubi-Benhammou, N.; Ghembaza, N.; Atik, B.F. Comparative studies on antioxidant activities of extracts from the leaf, stem and berry of Myrtus communis L. Int. Food Res. J. 2014, 21, 1957–1962. [Google Scholar]
- Moghaddam, M.; Pirbalouti, A.G.; Mehdizadeh, L.; Pirmoradi, M.R. Changes in composition and essential oil yield of Ocimum ciliatum at different phenological stages. Eur. Food Res. Technol. 2015, 240, 199–204. [Google Scholar] [CrossRef]
- Bouyahya, A.; Dakka, N.; Talbaoui, A.; Et-Touys, A.; El-Boury, H.; Abrini, J.; Bakri, Y. Correlation between phenological changes, chemical composition and biological activities of the essential oil from Moroccan endemic Oregano (Origanum compactum Benth) 2017. Ind. Crops Prod. 2017, 108, 729–737. [Google Scholar] [CrossRef]
- Shadi, A.; Saharkhiz, M.J. Changes in essential oil contents and chemical compositions of Artemisiasieberi at different phenological growth stages. Anal. Chem. Lett. 2016, 6, 249–256. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, A.R.; Patel, B.D.; Shrestha, K. Investigation on phytochemical, antimicrobial activity and essential oil constituents of Nardostachys jatamansi DC. in different regions of Nepal. J. Coast. Life Med. 2016, 4, 56–60. [Google Scholar] [CrossRef]
- Aidi Wannes, W.A.; Mhamdi, B.; Marzouk, B. Variations in essential oil and fatty acid composition during Myrtus communis var. italica fruit maturation. Food Chem. 2009, 112, 621–626. [Google Scholar] [CrossRef]
- Fadda, A.; Mulas, M. Chemical changes during myrtle (Myrtus communis L.) fruit development and ripening. Sci. Hortic. 2010, 125, 477–485. [Google Scholar] [CrossRef]
- Bouzabata, A.; Cabral, C.; Gonçalves, M.J.; Cruz, M.T.; Bighelli, A.; Cavaleiro, C.; Salgueiro, L. Myrtus communis L. as source of a bioactive and safe essential oil. Food Chem. Toxicol. 2015, 75, 166–172. [Google Scholar] [CrossRef]
- Saifullah, M.; McCullum, R.; McCluskey, A.; Vuong, Q. Effects of different drying methods on extractable phenolic compounds and antioxidant properties from lemon myrtle dried leaves. Heliyon 2019, 5, e03044. [Google Scholar] [CrossRef]
- Abu-Darwish, M.S.; Alu’datt, M.H.; Al-Tawaha, A.R.; Ereifej, K.; Almajwal, A.; Odat, N.; Al Khateeb, W. Seasonal variation in essential oil yield and composition from Thymus vulgaris L. during different growth stages in the south of Jordan. Nat. Prod. Res. 2012, 26, 1310–1317. [Google Scholar] [CrossRef]
- Saharkhiz, M.J.; Kamyab, A.A.; Kazerani, N.K.; Zomorodian, K.; Pakshir, K.; Rahimi, M.J. Chemical compositions and antimicrobial activities of Ocimum sanctum L. essential oils at different harvest stages. Jundishapur J. Microbiol. 2015, 8, e13720. [Google Scholar] [CrossRef]
- Kim, D.Y.; Won, K.J.; Hwang, D.I.; Park, S.M.; Kim, B.; Lee, H.M. Chemical composition, antioxidant and anti-melanogenic activities of essential oils from Chrysanthemum boreale makino at different harvesting Stages. Chem. Biodivers. 2018, 15, e1700506. [Google Scholar] [CrossRef] [PubMed]
- Khalvandi, M.; Amerian, M.; Pirdashti, H.; Keramati, S.; Hosseini, J. Essential oil of peppermint in symbiotic relationship with Piriformospora indica and methyl jasmonate application under saline condition. Ind. Crops Prod. 2019, 127, 195–202. [Google Scholar] [CrossRef]
- Esmaeili, H.; Karami, A.; Maggi, F. Essential oil composition, total phenolic and flavonoids contents, and antioxidant activity of Oliveri adecumbens Vent. (Apiaceae) at different phenological stages. J. Clean. Prod. 2018, 198, 91–95. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E. In vitro antioxidant activity and scavenging effects of Cinnamomum verum leaf extract assayed by different methodologies. Food Chem. Toxicol. 2006, 44, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Introduction: Biochemistry, physiology and ecological functions of secondary metabolites. Annu. Rev. Plant. Biol. 2018, 40, 1–19. [Google Scholar] [CrossRef]
- Lemoine, N.P.; Doublet, D.; Salminen, J.P.; Burkepile, D.E.; Parker, J.D. Responses of plant phenology, growth, defense, and reproduction to interactive effects of warming and insect herbivory. Ecology 2017, 98, 1817–1828. [Google Scholar] [CrossRef]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of light on secondary metabolites in selected leafy greens: A review. Front. Plant. Sci. 2020, 11, 497. [Google Scholar] [CrossRef]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S.A. Effect of salicylic acid and sodium nitroprusside on growth parameters, photosynthetic pigments and secondary metabolites of safflower under drought stress. Sci. Hortic. 2020, 259, 108823. [Google Scholar] [CrossRef]
- Özcan, M.M.; Al Juhaimi, F.; Ahmed, I.A.M.; Babiker, E.E.; Ghafoor, K. Antioxidant activity, fatty acid composition, phenolic compounds and mineral contents of stem, leave and fruits of two morphs of wild myrtle plants. J. Food Meas. Charact. 2020, 14, 1376–1382. [Google Scholar] [CrossRef]
- Asik, S.; Atbakan Kalkan, T.; Topuz, A. Optimization of spray drying condition and wall material composition for myrtle extract powder using response surface methodology. Dry. Technol. 2021, 39, 1869–1882. [Google Scholar] [CrossRef]
- Kang, E.J.; Lee, J.K.; Park, H.R.; Kim, H.; Kim, H.S.; Park, J. Antioxidant and anti-inflammatory activities of phenolic compounds extracted from lemon myrtle (Backhousia citriodora) leaves at various extraction conditions. Food Sci. Biotechnol. 2020, 29, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Crouch, R.A.; Awadh Ali, N.A.; Al-Fatimi, M.A.; Setzer, W.N.; Wessjohann, L. Hierarchical cluster analysis and chemical characterisation of Myrtus communis L. essential oil from Yemen region and its antimicrobial, antioxidant and anti-colorectal adenocarcinoma properties. Nat. Prod. Res. 2017, 31, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Hayder, N.; Abdelwahed, A.; Kilani, S.; Ben Ammar, R.; Mahmoud, A.; Ghedira, K.; Chekir-Ghedira, L. Anti-genotoxic and free radical scavenging activity of extracts from (Tunisian) Myrtus communis. Mutat. Res. 2004, 564, 89–95. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Academic press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Akroum, S.; Bendjeddou, D.; Satta, D.; Lalaoui, K. Antibacterial, antioxidant and acute toxicity tests on flavonoids extracted from some medicinal plants. Int. J. Green Pharm. 2010, 4, 165–169. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant. Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Yosr, Z.; Hnia, C.; Rim, T.; Mohamed, B. Changes in essential oil composition and phenolic fraction in Rosmarinus officinalis L. var. typicus Batt. organs during growth and incidence on the antioxidant activity. Ind. Crops Prod. 2013, 43, 412–419. [Google Scholar] [CrossRef]
- Musa Özcan, M.; Claude Chalchat, J. Effect of collection time on chemical composition of the essential oil of Foeniculum vulgare subsp. piperitum growing wild in Turkey. Eur. Food Res. Technol. 2006, 224, 279–281. [Google Scholar] [CrossRef]
- Mollaei, S.; Ebadi, M.; Hazrati, S.; Habibi, B.; Gholami, F.; Sourestani, M.M. Essential oil variation and antioxidant capacity of Mentha pulegium populations and their relation to ecological factors. Biochem. Syst. Ecol. 2020, 91, 104084. [Google Scholar] [CrossRef]
- Bajalan, I.; Rouzbahani, R.; Ghasemi Pirbalouti, A.; Maggi, F. Quali-quantitative variation of essential oil from Iranian rosemary (Rosmarinus officinalis L.) accessions according to environmental factors. J. Essent. Oil-Bear. Plants 2018, 30, 16–24. [Google Scholar] [CrossRef]
- Bakhtiar, A.; Khaghani, S.; Ghasemi Pirbalouti, A.; Gomarian, M.; Chavoshi, S. Essential oil variation among different populations of Ziziphora tenuior L. cultivated at semiarid climate. J. Essent. Oil-Bear. Plants 2021, 33, 385–393. [Google Scholar] [CrossRef]
- Puri, S.K.; Habbu, P.V.; Kulkarni, P.V.; Kulkarni, V.H. Nitrogen containing secondary metabolites from endophytes of medicinal plants and their biological/pharmacological activities—A review. Sys. Rev. Pharm. 2018, 9, 22–30. [Google Scholar] [CrossRef]
- Alhaithloul, H.A.; Soliman, M.H.; Ameta, K.L.; El-Esawi, M.A.; Elkelish, A. Changes in ecophysiology, osmolytes, and secondary metabolites of the medicinal plants of Mentha piperita and Catharanthus roseus subjected to drought and heat stress. Biomolecules 2019, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Karahan, F.; Ozyigit, I.I.; Saracoglu, I.A.; Yalcin, I.E.; Ozyigit, A.H.; Ilcim, A. Heavy metal levels and mineral nutrient status in different parts of various medicinal plants collected from eastern Mediterranean region of Turkey. Biol. Trace Elem. Res. 2020, 197, 316–329. [Google Scholar] [CrossRef]
- Usai, M.; Marchetti, M.; Culeddu, N.; Mulas, M. Chemical composition of myrtle (Myrtus communis L.) berries essential oils as observed in a collection of genotypes. Molecules 2018, 23, 2502. [Google Scholar] [CrossRef]
- Siracusa, L.; Napoli, E.; Tuttolomondo, T.; Licata, M.; La Bella, S.; Gennaro, M.C.; Ruberto, G. A Two-Year Bio-Agronomic and Chemotaxonomic Evaluation of Wild Sicilian Myrtle (Myrtus communis L.) Berries and Leaves. Chem. Biodivers. 2019, 16, e1800575. [Google Scholar] [CrossRef]
- Giampieri, F.; Cianciosi, D.; Forbes-Hernández, T.Y. Myrtle (Myrtus communis L.) berries, seeds, leaves, and essential oils: New undiscovered sources of natural compounds with promising health benefits. Food Front. 2020, 1, 276–295. [Google Scholar] [CrossRef]
- Boelens, M.H.; Jimenez, R. The chemical composition of Spanish myrtle oils. Part II. J. Essent. Oil-Bear. Plants 1992, 4, 349–353. [Google Scholar] [CrossRef]
- Sarmoum, R.; Haid, S.; Biche, M.; Djazouli, Z.; Zebib, B.; Merah, O. Effect of salinity and water stress on the essential oil Rosemary essential oil composition (Rosmarinus officinalis). Argonomy 2019, 9, 214. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Mirbagheri, H.; Hamedi, B.; Rahimi, E. Antibacterial activity of the essential oils of myrtle leaves against Erysipelothrix rhusiopathiae. Asian Pac. J. Trop Biomed. 2014, 4, S505–S509. [Google Scholar] [CrossRef]
- Bouaziz, A.; Abdalla, S.; Baghiani, A.; Charef, N. Phytochemical analysis, hypotensive effect and antioxidant properties of Myrtus communis L. growing in Algeria. Asian Pac. J. Trop Biomed. 2015, 5, 19–28. [Google Scholar] [CrossRef]
- Romani, A.; Coinu, R.; Carta, S.; Pinelli, P.; Galardi, C.; Vincieri, F.F.; Franconi, F. Evaluation of antioxidant effect of different extracts of Myrtus communis L. Free Radic. Res. 2004, 38, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Amensour, M.; Sendra, E.; Abrini, J.; Pérez-Alvarez, J.A.; Fernandez-Lopez, J. Antioxidant activity and total phenolic compounds of myrtle extracts actividad antioxidante y contenido de compuestos fenólicos totales en extractos de myrtus. CYTA J. Food 2010, 8, 95–101. [Google Scholar] [CrossRef]
- Tuberoso, C.I.; Melis, M.; Angioni, A.; Pala, M.; Cabras, P. Myrtle hydroalcoholic extracts obtained from different selections of Myrtus communis L. Food Chem. 2007, 101, 806–811. [Google Scholar] [CrossRef]
- Montoro, P.; Tuberoso, C.I.G.; Piacente, S.; Perrone, A.; De Feo, V.; Cabras, P.; Pizza, C. Stability and antioxidant activity of poliphenols in extract of Myrtus communis L. berries used for the preparation of myrtle liqueur. J. Pharm. Biomed. Anal. 2006, 41, 1614–1619. [Google Scholar] [CrossRef] [PubMed]
- Riipi, M.; Ossipov, V.; Lempa, K.; Haukioja, E.; Koricheva, J.; Ossipova, S.; Pihlaja, K. Seasonal changes in birch leaf chemistry: Are there trade-offs between leaf growth and accumulation of phenolics? Oecologia 2000, 130, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, Y.; Li, W.; Zhao, L.; Meng, F.; Wang, Y.; Tan, H.; Yang, H.; Wei, C.; Wan, X.; et al. Tissue-specific, development dependent phenolic compounds accumulation profile and gene expression pattern in tea plant [Camellia sinensis]. PLoS ONE 2013, 8, e62315. [Google Scholar] [CrossRef]
- Maina, S.; Ryu, D.H.; Bakari, G.; Misinzo, G.; Nho, C.W.; Kim, H.Y. Variation in phenolic compounds and antioxidant activity of various organs of African Cabbage (Cleome gynandra L.) accessions at different growth stages. Antioxidants 2021, 10, 1952. [Google Scholar] [CrossRef]
- Vvedenskaya, I.O.; Vorsa, N. Flavonoid composition over fruit development and maturation in American cranberry, Vaccinium macrocarpon Ait. Plant. Sci. 2004, 167, 1043–1054. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Dupont, N.; Guillermin, P. Concentration and characteristicsof procyanidins and other phenolics in apples during fruit growth. Phytochemistry 2007, 68, 1128–1138. [Google Scholar] [CrossRef]
- Rekha, C.; Poornima, G.; Manasa, M.; Abhipsa, V.; Devi, J.P.; Kumar, H.T.V.; Kekuda, T.R.P. Ascorbic acid, total phenol content and antioxidant activity of fresh juices of four ripe and unripe citrus fruits. Chem. Sci. Trans. 2012, 1, 303–310. [Google Scholar] [CrossRef]
- Kainama, H.; Fatmawati, S.; Santoso, M.; Papilaya, P.M.; Ersam, T. The relationship of free radical scavenging and total phenolic and flavonoid contents of Garcinia lasoar PAM. Pharm. Chem. J. 2020, 53, 1151–1157. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- El Atki, Y.; Aouam, I.; Taroq, A.; Lyoussi, B.; Taleb, M.; Abdellaoui, A. Total phenolic and flavonoid contents and antioxidant activities of extracts from Teucrium polium growing wild in Morocco. Mater. Today Proc. 2019, 13, 777–783. [Google Scholar] [CrossRef]
- Ghafoor, K.; Al Juhaimi, F.; Özcan, M.M.; Uslu, N.; Babiker, E.E.; Ahmed, I.A.M. Total phenolics, total carotenoids, individual phenolics and antioxidant activity of ginger (Zingiber officinale) rhizome as affected by drying methods. LWT—Food Sci Technol. 2020, 126, 109354. [Google Scholar] [CrossRef]
- Sunil, C.; Kumar, V.; Van Staden, J. In vitro alpha-glucosidase inhibitory, total phenolic composition, antiradical and antioxidant potential of Heteromorpha arborescens (Spreng.) Cham. & Schltdl. leaf and bark extracts. S. Afr. J. Bot. 2019, 124, 380–386. [Google Scholar] [CrossRef]
- Saffaryazdi, A.; Ganjeali, A.; Farhoosh, R.; Cheniany, M. Variation in phenolic compounds, α-linolenic acid and linoleic acid contents and antioxidant activity of purslane (Portulaca oleracea L.) during phenological growth stages. Physiol. Mol. Biol. Plants 2020, 26, 1519–1529. [Google Scholar] [CrossRef]
- Hazrati, S.; Beidaghi, P.; Beyraghdar Kashkooli, A.; Hosseini, S.J.; Nicola, S. Effect of harvesting time variations on essential oil yield and composition of Sage (Salvia officinalis). Horticulturae 2022, 8, 149. [Google Scholar] [CrossRef]
- Soltani Howyzeh, M.; Sadat Noori, S.A.; Shariati, J.V.; Niazian, M. Essential oil chemotype of iranian ajowan (Trachyspermum ammi L.). J. Essent. Oil-Bear. Plants 2018, 21, 273–276. [Google Scholar] [CrossRef]
- Pank, F.; Pfefferkorn, A.; Kruger, H. Evaluation of a summer savory collection (Satureja hortensis L.) with regard to morphology, precocity, yield components and essential oil and carcacrol content. Z. Fur Arznei-Gewurzpflanzen 2004, 9, 72–78. [Google Scholar]
- Hadian, J.; Ebrahimi, S.N.; Salehi, P. Variability of morphological and phytochemical characteristics among Satureja hortensis L. accessions of Iran. Ind. Crops Prod. 2010, 32, 62–69. [Google Scholar] [CrossRef]
- Hadian, J.; Esmaeili, H.; Nadjafi, F.; Khadivi-Khub, A. Essential oil characterization of Satureja rechingeri in Iran. Ind. Crops Prod. 2014, 61, 403–409. [Google Scholar] [CrossRef]
- Hazrati, S.; Rowshan, V.; Hosseini, S.J.; Sedaghat, M.; Mohammadi, H. Variation of essential oil composition and antioxidant activity in aerial parts of Stachys schtschegleevi Sosn at different growing stages. J. Essent. Oil-Bear. Plants 2020, 23, 1054–1071. [Google Scholar] [CrossRef]
- Hazrati, S.; Mollaei, S.; Rabbi Angourani, H.; Hosseini, S.J.; Sedaghat, M.; Nicola, S. How do essential oil composition and phenolic acid profile of Heracleum persicum fluctuate at different phenological stages? Food Sci. Nutr. 2020, 8, 6192–6206. [Google Scholar] [CrossRef] [PubMed]
- Ibraliu, A.; Mi, X.; Elezi, F. Variation in essential oils to study the biodiversity in Satureja montana L. J. Med. Plant. Res. 2011, 5, 2978–2989. [Google Scholar] [CrossRef] [PubMed]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Component | RI | Vegetative Stage (Leaves) | Flowering Stage (Leaves) | Flowering Stage (Flowers) | Unripe Fruit Stage (Leaves) | Unripe Fruit Stage (Fruits) | Ripe Fruit Stage (Leaves) | Ripe Fruit Stage (Fruits) | After the Fully Mature Fruit Dispersal Stage (Leaves) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Isobutyl isobutyrate | 911 | 0.12 ± 0.00 a | - | - | - | - | 0.10 ± 0.00 a | 0.04 ± 0.02 b | 0.10 ± 0.00 a |
| 2 | α-Thujene | 927 | 0.12 ± 0.01 a | 0.12 ± 0.00 a | 0.14 ± 0.00 a | 0.12 ± 0.00 a | 0.15 ± 0.01 a | 0.11 ± 0.01 a | 0.11 ± 0.01 a | 0.05 ± 0.03 b |
| 3 | α-Pinene | 938 | 25.03 ± 0.62 b | 32.84 ± 0.32 a | 19.01 ± 0.58 c | 19.42 ± 0.72 c | 15.00 ± 2.08 d | 10.09 ± 0.61 e | 11.50 ± 2.15 e | 8.88 ± 0.33 e |
| 4 | β-Pinene | 979 | 1.32 ± 0.36 a | 0.60 ± 0.01 b | 0.34 ± 0.01 b | 0.52 ± 0.08 b | 0.10 ± 0.00 b | 0.27 ± 0.02 b | 0.43 ± 0.22 b | 0.65 ± 0.26 b |
| 5 | δ−3-Carene | 1013 | 0.78 ± 0.33 a | 0.65 ± 0.05 a | 0.18 ± 0.01 b | 0.18 ± 0.01 b | 0.20 ± 0.01 b | 0.13 ± 0.01 b | 0.11 ± 0.01 b | 0.25 ± 0.03 b |
| 6 | p-Cymene | 1028 | 0.98 ± 0.51 a | 1.00 ± 0.05 a | 0.20 ± 0.01 b | 0.43 ± 0.05 ab | 0.20 ± 0.03 b | 0.32 ± 0.01 b | 0.24 ± 0.01 b | 0.17 ± 0.01 b |
| 7 | Limonene | 1031 | 1.42 ± 0.12 b | 0.80 ± 0.06 c | 23.97 ± 0.09 a | 0.85 ± 0.18 c | 0.64 ± 0.31 cd | 1.07 ± 0.08 bc | 0.14 ± 0.01 e | 0.33 ± 0.11 de |
| 8 | 1,8-Cineole | 1035 | 45.98 ± 2.50 a | 33.22 ± 0.98 c | 12.56 ± 0.26 f | 22.77 ± 0.38 d | 16.01 ± 0.15 e | 38.52 ± 0.37 b | 34.74 ± 0.20 c | 44.54 ± 0.85 a |
| 9 | γ-Terpinene | 1060 | 0.37 ± 0.14 b | 0.11 ± 0.00 c | 0.11 ± 0.01 c | 0.19 ± 0.01 bc | 1.34 ± 0.13 a | 0.15 ± 0.01 c | 0.13 ± 0.01 c | 0.20 ± 0.01 bc |
| 10 | Linalool | 1106 | 8.12 ± 0.01 e | 10.56 ± 0.27 d | 16.14 ± 0.64 c | 15.54 ± 0.35 c | 16.18 ± 1.08 c | 23.37 ± 0.01 a | 17.39 ± 0.92 c | 19.59 ± 0.47 b |
| 11 | α-Terpineol | 1198 | 3.57 ± 0.25 e | 5.74 ± 0.16 d | 8.45 ± 0.03 b | 15.17 ± 0.81 a | 8.34 ± 0.16 b | 8.90 ± 0.49 b | 6.15 ± 1.38 cd | 7.73 ± 0.22 bc |
| 12 | Methyl chavicol | 1206 | 0.62 ± 0.29 bc | 1.54 ± 0.05 a | 0.90 ± 0.06 b | 0.78 ± 0.19 b | 0.66 ± 0.03 bc | 0.48 ± 0.08 b-d | 0.27 ± 0.03 cd | 0.16 ± 0.01 d |
| 13 | β-Citronellol | 1237 | 0.41 ± 0.12 b | 1.22 ± 0.38 a | 0.16 ± 0.02 b | 0.67 ± 0.12 b | 0.41 ± 0.08 b | 0.56 ± 0.08 b | 0.23 ± 0.02 b | 0.36 ± 0.12 b |
| 14 | Linalyl acetate | 1257 | 2.79 ± 0.28 c | 2.71 ± 0.35 c | 2.45 ± 0.03 c | 6.86 ± 0.51 a | 5.57 ± 0.75 b | 5.18 ± 0.08 b | 3.22 ± 0.31 c | 4.96 ± 0.00 b |
| 15 | Methyl citronellate | 1262 | 0.77 ± 0.44 cd | 2.04 ± 0.02 b | 0.18 ± 0.01 d | 2.62 ± 0.28 b | 5.04 ± 0.34 a | 0.31 ± 0.03 d | 5.00 ± 0.08 a | 1.19 ± 0.01 c |
| 16 | α-Terpinyl acetate | 1348 | 1.34 ± 0.12 d | 2.85 ± 0.23 c | 1.13 ± 0.04 d | 5.79 ± 0.33 a | 2.34 ± 0.14 cd | 2.86 ± 0.18 c | 4.46 ± 1.10 b | 3.26 ± 0.14 bc |
| 17 | Neryl acetate | 1363 | 0.11 ± 0.06 c | 0.14 ± 0.00 c | 0.69 ± 0.01 a-c | 0.12 ± 0.00 c | 0.81 ± 0.05 ab | 0.10 ± 0.00 c | 0.27 ± 0.08 bc | 1.14 ± 0.56 a |
| 18 | Geranyl acetate | 1380 | 0.45 ± 0.18 d | 0.13 ± 0.01 d | 2.99 ± 1.27 c | 0.71 ± 0.05 d | 10.83 ± 0.73 a | 1.18 ± 0.38 d | 5.81 ± 0.19 b | 0.14 ± 0.01 d |
| 19 | Methyleugenol | 1404 | 0.38 ± 0.14 e | 0.19 ± 0.01 e | 7.73 ± 0.19 a | 1.00 ± 0.06 d | 6.46 ± 0.19 c | 0.10 ± 0.01 e | 7.19 ± 0.02 b | 0.10 ± 0.00 e |
| 20 | trans-Caryophyllene | 1416 | 0.11 ± 0.00 b | 0.12 ± 0.01 b | 0.10 ± 0.01 b | 0.05 ± 0.03 b | 1.35 ± 0.14 a | 0.11 ± 0.01 b | 0.05 ± 0.03 b | 0.12 ± 0.01 b |
| 21 | α-Humulene | 1451 | 0.23 ± 0.06 b | 0.13 ± 0.01 cd | - | 0.18 ± 0.01 bc | 0.60 ± 0.02 a | 0.12 ± 0.01 cd | 0.07 ± 0.04 de | 0.12 ± 0.01 cd |
| 22 | Spathulenol | 1583 | 0.11 ± 0.00 b | 0.14 ± 0.02 b | - | 0.10 ± 0.01 bc | 0.31 ± 0.06 a | 0.13 ± 0.00 b | 0.12 ± 0.00 b | 0.16 ± 0.03 b |
| 23 | Caryophyllene oxide | 1609 | 0.12 ± 0.00 b | 0.13 ± 0.01 b | 0.10 ± 0.00 b | 0.14 ± 0.02 b | 0.43 ± 0.08 a | 0.15 ± 0.02 b | 0.11 ± 0.01 b | 0.18 ± 0.01 b |
| Total (%) | 94.91 | 96.98 | 97.53 | 94.21 | 92.97 | 94.31 | 97.78 | 94.38 | ||
| Traits | Vegetative Stage (Leaves) | Flowering Stage (Leaves) | Flowering Stage (Flowers) | Unripe Fruit Stage (Leaves) | Unripe Fruit Stage (Fruits) | Ripe Fruit Stage (Leaves) | Ripe Fruit Stage (Fruits) | After the Fully Mature Fruit Dispersal Stage (Leaves) |
|---|---|---|---|---|---|---|---|---|
| Total flavonoids content (mg QE/g DW) | 7.49 ± 0.18 a | 5.41 ± 0.29 c | 3.68 ± 0.10 e | 5.71 ± 0.15 c | 4.79 ± 0.10 d | 6.23 ± 0.15 b | 2.69 ± 0.16 f | 5.84 ± 0.09 bc |
| Total phenolic content (mg GAE/g DW) | 66.52 ± 0.40 a | 43.92 ± 0.05 c | 27.50 ± 0.29 e | 49.67 ± 0.77 b | 18.83 ± 0.60 f | 30.22 ± 1.75 d–f | 15.30 ± 0.75 g | 32.33 ± 1.45 d |
| IC50(μg/mL) | 31.04 ± 0.54 f | 273.30 ± 8.83 e | 519.89 ± 20.84 b | 355.26 ± 2.97 d | 469.50 ± 1.04 c | 476.17 ± 6.99 c | 835.68 ± 18.23 a | 294.27 ± 14.45 e |
| Compounds | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| Isobutyl isobutyrate (x1) | −0.772 | 0.098 | −0.339 | −0.335 | 0.123 |
| α-Thujene (x2) | 0.280 | 0.310 | 0.761 | 0.160 | −0.190 |
| α-Pinene (x3) | −0.621 | 0.398 | 0.571 | 0.311 | −0.082 |
| β-Pinene (x4) | −0.828 | 0.135 | −0.040 | −0.352 | −0.106 |
| δ−3-Carene (x5) | −0.810 | 0.498 | 0.230 | −0.172 | 0.035 |
| p-Cymene (x6) | −0.843 | 0.431 | 0.228 | 0.074 | −0.198 |
| Limonene (x7) | 0.137 | −0.474 | 0.736 | −0.009 | 0.411 |
| 1,8-Cineole (x8) | −0.636 | −0.029 | −0.589 | −0.455 | −0.114 |
| γ-Terpinene (x9) | 0.574 | 0.786 | 0.002 | −0.114 | 0.138 |
| Linalool (x10) | 0.556 | −0.507 | −0.500 | 0.052 | 0.191 |
| α-Terpineol (x11) | 0.358 | −0.234 | −0.250 | 0.782 | 0.000 |
| Methyl chavicol (x12) | −0.361 | 0.290 | 0.617 | 0.566 | 0.067 |
| β-Citronellol (x13) | −0.500 | 0.363 | −0.038 | 0.621 | −0.056 |
| Linalyl acetate (x14) | 0.396 | 0.146 | −0.676 | 0.511 | 0.024 |
| Methyl citronellate (x15) | 0.611 | 0.402 | −0.055 | −0.010 | −0.572 |
| α-Terpinyl acetate (x16) | 0.161 | −0.201 | −0.521 | 0.515 | −0.561 |
| Neryl acetate (x17) | 0.494 | −0.036 | −0.125 | −0.283 | 0.668 |
| Geranyl acetate (x18) | 0.851 | 0.405 | 0.204 | −0.208 | −0.155 |
| Methyleugenol (x19) | 0.738 | −0.134 | 0.581 | −0.248 | −0.161 |
| trans-Caryophyllene (x20) | 0.648 | 0.731 | 0.050 | −0.044 | 0.195 |
| α-Humulene (x21) | 0.430 | 0.881 | −0.120 | −0.006 | 0.062 |
| Spathulenol (x22) | 0.442 | 0.777 | −0.387 | −0.050 | 0.040 |
| Caryophyllene oxide (x23) | 0.647 | 0.705 | −0.157 | 0.009 | 0.243 |
| Essential oil content (x24) | −0.165 | −0.087 | −0.061 | 0.547 | 0.751 |
| Flavonoids (x25) | −0.622 | 0.435 | −0.574 | −0.034 | 0.020 |
| Total phenols (x26) | −0.848 | 0.245 | −0.016 | 0.142 | −0.005 |
| IC50 (x27) | 0.721 | −0.450 | 0.106 | −0.054 | −0.402 |
| Eigenvalue | 9.623 | 5.365 | 4.372 | 2.973 | 2.322 |
| Relative variance (%) | 35.641 | 19.871 | 16.194 | 11.012 | 8.602 |
| Cumulative variance (%) | 35.641 | 55.513 | 71.707 | 82.719 | 91.321 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazrati, S.; Hosseini, S.J.; Ebadi, M.-T.; Nicola, S. Evolution of Phytochemical Variation in Myrtle (Myrtus communis L.) Organs during Different Phenological Stages. Horticulturae 2022, 8, 757. https://doi.org/10.3390/horticulturae8090757
Hazrati S, Hosseini SJ, Ebadi M-T, Nicola S. Evolution of Phytochemical Variation in Myrtle (Myrtus communis L.) Organs during Different Phenological Stages. Horticulturae. 2022; 8(9):757. https://doi.org/10.3390/horticulturae8090757
Chicago/Turabian StyleHazrati, Saeid, Seyyed Jaber Hosseini, Mohammad-Taghi Ebadi, and Silvana Nicola. 2022. "Evolution of Phytochemical Variation in Myrtle (Myrtus communis L.) Organs during Different Phenological Stages" Horticulturae 8, no. 9: 757. https://doi.org/10.3390/horticulturae8090757
APA StyleHazrati, S., Hosseini, S. J., Ebadi, M. -T., & Nicola, S. (2022). Evolution of Phytochemical Variation in Myrtle (Myrtus communis L.) Organs during Different Phenological Stages. Horticulturae, 8(9), 757. https://doi.org/10.3390/horticulturae8090757