1. Introduction
Peony (
Paeonia suffruticosa Andr.) is a deciduous flowering shrub of the genus
Paeonia in the family
Paeoniaceae. The cultivation of peony originates from China. Peony is not only abundant in China but is also widely distributed. From the three provinces in northeast China to the border of southern Xinjiang and from the coast of the East China Sea to the Qinghai–Tibet Plateau, there are wild peony resources as well as areas of artificial cultivation. Due to the influence of climate, history, cultural customs, and many other factors, the distribution of the cultivated peony population in China has resulted in the Yellow River Basin being the main cultivation area. Peony can be roughly divided into three populations: the cold-tolerant peony represented by the purple-spot peony of Gansu province and the heat-resistant peony represented by the Tongling peony of Anhui province; peony plants from Luoyang, Henan Province; and peony plants from Caozhou, Shandong province. Peony has large and beautiful flowers as well as abundant flower colors and a pleasant fragrance, resulting in peony being known as the “king of flowers”. They have a good condition of industrialization: the traditional culture of peonies with unique characteristics has created a good social atmosphere for the industrialization of peonies; the natural beauty of peonies and the rich variety peony resources lay the foundation for the industrialization of peonies, and the increasingly mature cultivation methods as well as the application technologies for peony cultivation provide the technical premises and guarantee peony industrialization. The broad domestic and international market prospects and huge developmental potential also represent positives for peony industrialization. Peony has ornamental, economic, and medicinal values. Due to different processing conditions, the peony alias Danpi can be divided into guudan or even Dan. It is commonly used as a material in Chinese medicine. It uses root bark for medicinal purposes. It has the functions of reducing fever and cooling blood, dispersing stasis, and passing through the meridian. It has effects on symptoms such as fever, hematemesis, amenorrhea, abdominal pain, stasis, hypertension, stroke, acute and chronic appendicitis, rhinitis, etc. Pharmacology shows that paeonol can lower blood pressure, in addition to analgesic, sedative, antipyretic, anti-inflammatory, and bactericidal activity. Currently, four large tree peony groups have been described, with nearly 1000 cultivars existing in China. Peony is widely used as a potted or cut flower, and the essential oil made from its petal components has medicinal properties, including anti-cancer, anti-inflammatory, and anti-oxidation activities. More recently, the seeds from some peony cultivars have been identified as excellent sources of α-linolenic acid, so peony seeds can be used to produce plant oil. Peony plants prefer warm, cool, and dry environments, and the high temperatures in southern summers seriously affect the growth and development of peony, with some cultivars being on the verge of extinction [
1,
2,
3,
4]. Photosynthesis is the most basic and complex process of all green plants. High-temperature stress significantly reduces the photosynthetic capacity of plants and further restricts the growth and development of plants. Chloroplasts are the main organelle involved in photosynthesis, and the photosynthetic pigments, two photosystems (PS II and PS I), electron transport system, and CO
2 reduction pathway are important components of photosynthesis. The disruption of any part of the photosynthesis system is sufficient to inhibit the overall photosynthetic mechanism of plants [
5,
6], reducing stomatal conductance, the photochemical efficiency of reaction centers, the CO
2 assimilation capacity, and water-use efficiency [
7]. Studies have shown that high-temperature stress can lead to PSIIfailure, reduce electron transport efficiency, and increase the production of reactive oxygen species [
8]. For example, high temperatures reduce the photosynthetic capacity of cotton leaves and affect the growth and development of cotton [
9]. The maximum photosynthetic rate (Pn
max) and apparent quantum efficiency (AQE) of grape leaves were significantly decreased after high-temperature stress [
10], and irreversible damage to peony chloroplasts and their photosynthetic machinery was found after high-temperature stress treatment, resulting in the relative content of chlorophyll (SPAD) and the PS II parameters of Fo, Fm, and Fv/Fm as well as other chlorophyll fluorescence parameters being significantly reduced, resulting in the yellowing of plant leaves and a significant decrease in photosynthetic capacity [
11]. Obviously, studying the structure and function of PSII can reflect the responses of plants to high-temperature stress, and the rapid chlorophyll fluorescence induction curve and fluorescence parameters can be used to analyze a lot of information about the primary photochemistry of PSII reaction centers [
12]. In recent years, numerous studies have demonstrated that chlorophyll fluorescence analysis techniques can reflect the damage induced by high temperatures on plants and are an important technical tool for screening heat-tolerant plant species that are widely used in research on plant heat tolerance [
13,
14,
15,
16]. For example, by analyzing the relevant parameters of fast chlorophyll fluorescence-induced kinetic curves, it was found that the ability of PS II light energy capture and transfer to the downstream area of Q
A decreased more significantly in high-temperature-sensitive wheat varieties compared to high-temperature-insensitive wheat varieties and led to a rapid decrease in the comprehensive performance of photosynthesis, while the efficiency of electron transfer from the Q
A (the maximum rate of primary quinone receptor) receptor side to the end of PS I and the activity of the oxygen release complex did not decrease significantly [
17]. In conclusion, the rapid chlorophyll fluorescence induction kinetic analysis (JIP test) of PSII is an important tool to study thermal stability, which is of great significance when exploring the effects of heat stress and the damage mechanism and to rapidly identify the heat tolerance of different varieties of peony.
At present, studies on high-temperature stress in peony mainly focus on the effects of high-temperature stress on physiological characteristics [
18,
19] and the photosynthetic mechanism [
20,
21,
22,
23] of peony, with examples of findings being how high temperatures lead to a decrease in Pn, a decrease in the soluble protein content and SOD activity, and an increase in the proline content and MDA content in peony leaves; an increase in the electrical conductivity of peony leaves; the inhibition of chlorophyll synthesis or chlorophyll degradation; and a decreased photosynthetic rate decreased and increased lipid membrane permeability. However, there are no reports on the correlation between the photosynthetic characteristic curve, the kinetic curve of rapid chlorophyll fluorescence induction, and the heat tolerance of different peony varieties under high-temperature stress. In addition, there are few reports on the selection of photosynthetic fluorescence indexes in response to the heat resistance of peony, and the determination of these indexes is of great significance for the selection of heat-resistant varieties of peony, which could provide an effective reference for the introduction of peony varieties or for the evaluation of adaptability to high temperatures. Therefore, in this paper, three four-year-old peony varieties, ‘Fengdanbai’, ‘Huhong’, and ‘Yinhongqiaodui’, were used as the materials, and a high-temperature stress treatment was carried out via an artificial control test. Based on the previous analysis of the changes in the photosynthetic parameters and chlorophyll fluorescence kinetic indexes of ‘Fengdanbai’ under high-temperature stress [
24], the response differences in the photosystems of different peony varieties under high-temperature stress was systematically studied by photosynthetic characteristic parameters and fluorescence parameters. Through a horizontal comparison of different peony varieties, the key photosynthetic characteristic parameters and fluorescence parameters of peony varieties responding to high temperatures were screened, providing a theoretical basis and important reference for the rapid screening of peony varieties with high-temperature tolerance.
4. Discussion
Abiotic stress factors such as extreme temperatures, droughts and floods, and salinity are influential factors that govern plant growth and development [
26]. Studies have shown that the responses of plants to high-temperature stress is a complex and coordinated biological regulation process that results in changes in plant morphology [
27], while high-temperature stress affects the physiological responses of plants and can severely damage their own adversity-control ability and produce heat damage [
28]. Peony, which “likes mildness and avoids heat”, has a high light compensation point and a low light saturation point, and its effective use of light energy is narrow, so its photosynthetic physiology experiences stress under the high light and high temperature conditions during southern summers [
23,
29,
30]. To enrich the number of peony varieties in Jiangnan, high temperature is one of the most important factors among all constraints, and therefore, it is imperative to select and breed excellent heat-resistant peony varieties. Based on the analysis of the photosynthetic parameters and the chlorophyll fluorescence kinetic indexes of ‘Fengdanbai’ under high-temperature stress [
24], this study aimed to clarify the mechanism of peony’s response and adaptation to high-temperature stress, determine the indexes that can quickly screen the varieties responding to high temperatures, and lay the foundation for screening peony varieties with high-temperature tolerance by comparing and analyzing the differences in the photosynthetic curve parameters and the chlorophyll fluorescence kinetic curves of different peony varieties under high-temperature stress.
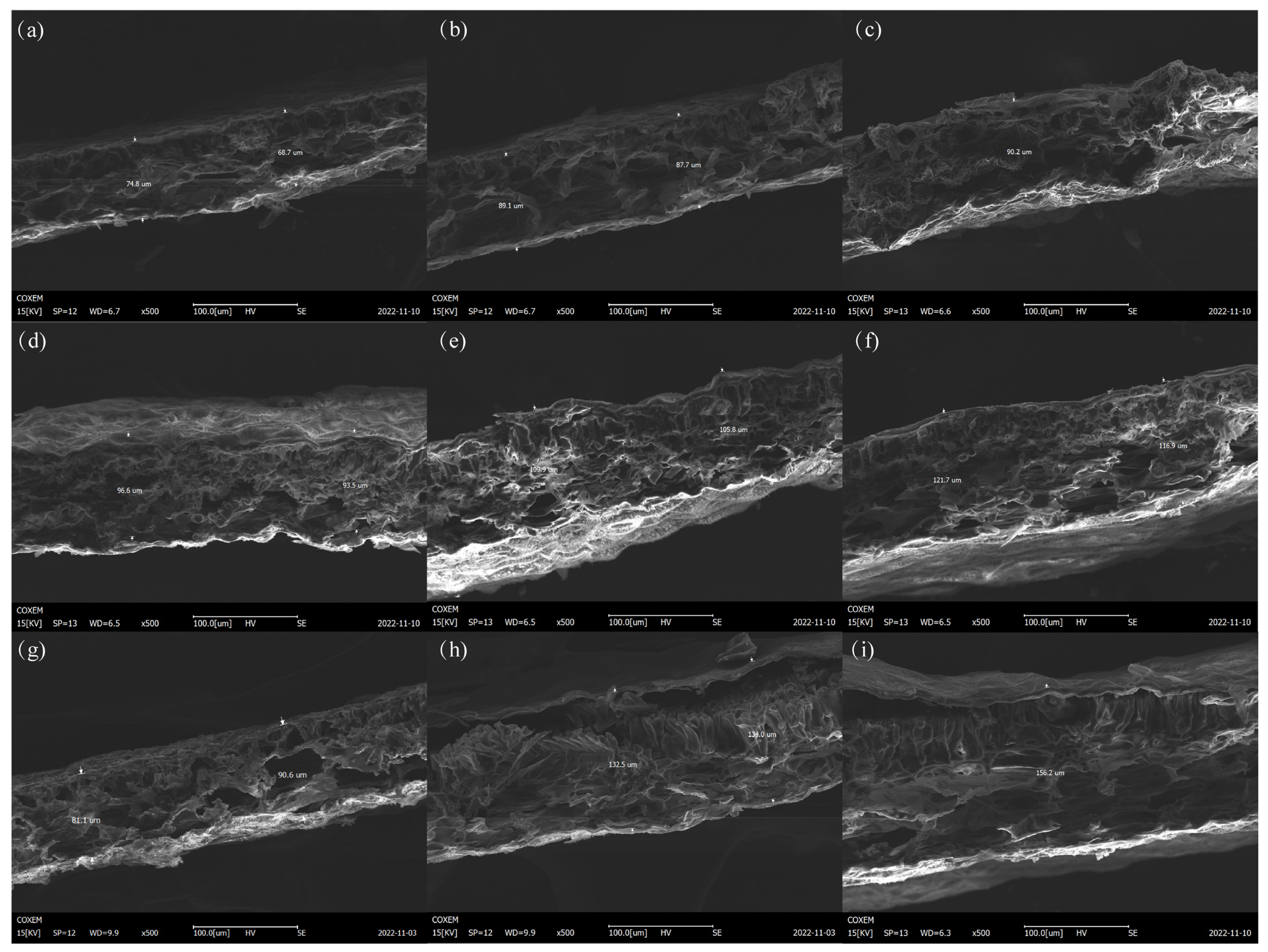
The leaf thicknesses of the different peony varieties are different and can be used as one of the indexes to measure the heat resistance of plants. Generally, thicker leaves have slower water transpiration and experience less water loss. In hot environments, the rate of water loss can also be reduced to alleviate the damage incurred by high temperature to leaves [
31]. In this study, the leaf thicknesses of the three peony cultivars increased as the high-temperature stress time was extended, with ‘Huhong’ increasing the most, which may have been due to the thickening of the leaves to plant reduce caused by the high temperatures.
Photosynthesis is sensitive to changes in temperature and plays an important role in plant growth and development, and studies have shown that high-temperature stress leads to photoinhibition in plants, damaging their photosystems and inactivating the Rubisco-activating enzyme for photosynthesis, resulting in a decrease in their ability to assimilate CO
2 and decreasing the photosynthetic rate [
32,
33,
34]. In the present study, the characteristic parameters of the photosynthetic curves of peony leaves: Pn
max and AQE, showed an overall decreasing trend with high-temperature stress, and this result was consistent with the results of Yuanda Zhang et al. [
35] and Chao Xu et al. [
36], indicating that high-temperature stress has a significant inhibitory effect on photosynthesis in peony leaves. For Pn
max, a parameter that reflects the potential photosynthetic capacity of the plant, the higher this value is, the lower the probability of the photoinhibition response occurring in plants under strong light conditions [
37], and usually, AQE is positively correlated with light energy utilization, and the higher the AQE value, the higher light energy utilization is [
38]. In this study, high-temperature stress significantly limited the Pn
max of peony, and the AQE of most peony species decreased significantly at the beginning of the stress period. This suggests that its leaves were severely damaged by chloroplasts at 40℃ and that fewer pigment protein complexes could absorb and convert the light energy in plants [
11,
24], while the short-term heat pressure did not significantly affect ‘Huhong’s light energy utilization capacity. This indicates that ‘Huhong’ is a more heat-tolerant species that can respond to high temperatures quickly in a relatively short period of time, and its ability to resist high-temperature stress during the early stages is stronger, while the ‘Fengdanbai’ variety needs cooling protection measures to prevent over-summering in the Jiangsu and Zhejiang regions.
LSP and LCP are two parameter indicators that can be used to gauge how well plants tolerate shade, which, in turn, can gauge how well plants react to high temperatures [
39]. At the early stage of high-temperature stress, the LSP of ‘Huhong’ decreased significantly, and LCP increased significantly, indicating that both the low and high light adaptations of this variety were weakened and that the light adaptation range was reduced to different degrees to alleviate the damage caused by high-temperature stress, which is consistent with the findings of Heli Zhao et al. [
40]. The reason for the decrease in Pn
max may be that high temperatures significantly reduce the accumulation of carbon in plants. All species undergo photosynthetic and respiratory adjustments to heat stress [
41], which result in LCP being different from LSP.
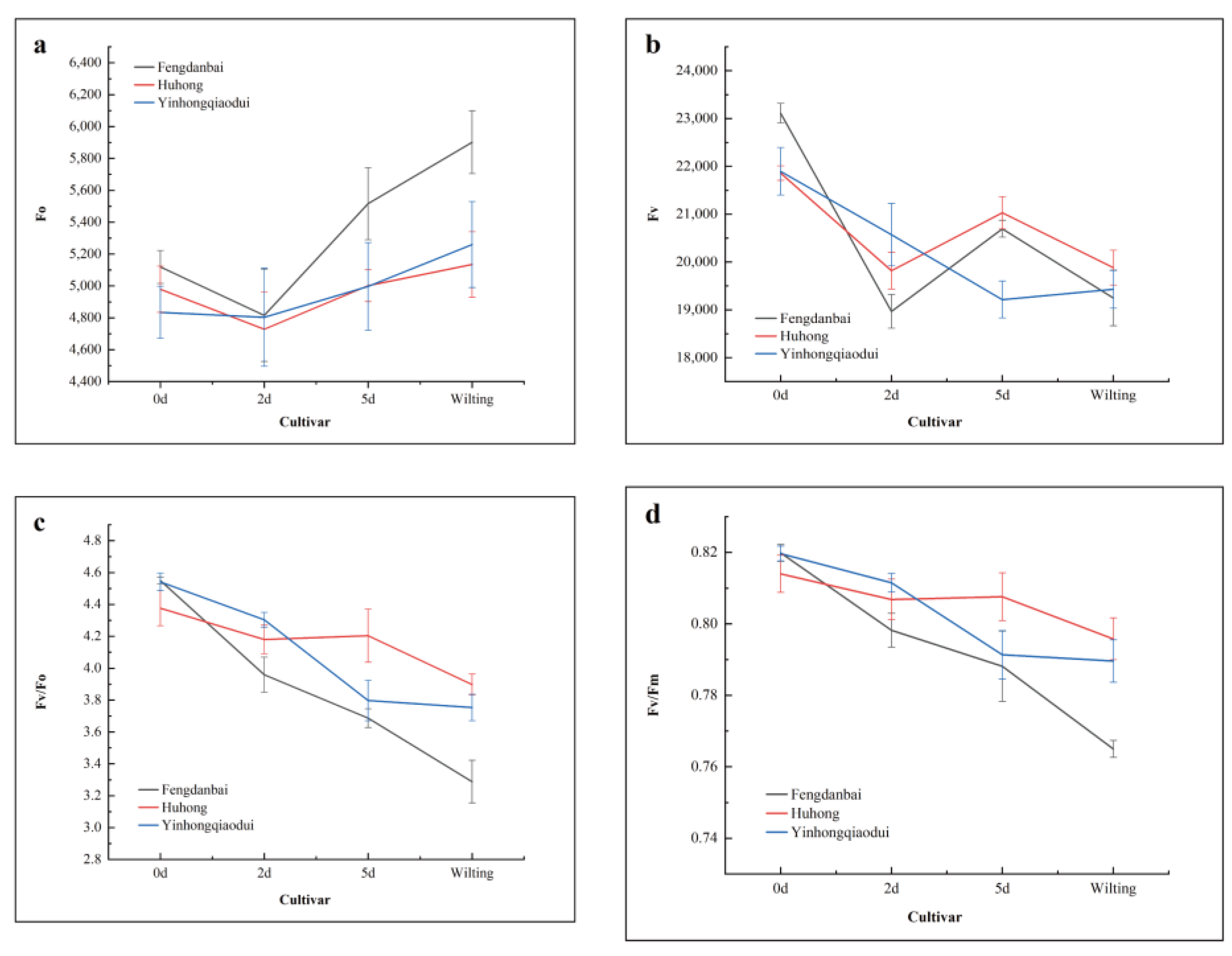
PS II, located on the membranes of plant cysts, is the most heat-sensitive part of the electron transport chain, and its electron transfer process is evident under chlorophyll fluorescence detection [
42,
43]. The rise and fall of J-point fluorescence in the OJIP curve can reflect the electron transfer activity from Q
A to Q
B on the P
680 receptor side [
44]; the J-I point reflects the complete reduction of the reduced plastoquinone (PQ) pool during the electron transfer process; the P point is when Q
A is completely in the reduced state and the PS II reaction center is closed: when the fluorescence yield is maximum. Studies by Murata et al. [
45] have shown that high temperatures can inhibit the synthesis of the D
1 protein by accelerating the production of H
2O
2, resulting in a significant reduction in electron transport between Q
A and Q
B, resulting in light inhibition. The OJIP curve of the J-I phase in the peony leaves was significantly reduced, indicating that the total reduced plastoquinone (PQ) library of peony leaves decreased after high-temperature stress, the photosynthetic properties were impaired, and the electron transport process of PS II was indeed inhibited, which is similar to the conclusions of previous studies [
40].
Donor-side and receptor-side activities can reflect the PS II activity in plants. High temperatures hinder PS II-related central vesicle energy transfer and can cause the inhibition of the electron transfer efficiency between the PS II donor side of cucumber (
Cucumis sativus L.) and tomato (
Lycopersicon esculentum Miller) and the acceptor side of winter wheat (
Triticum aestivum L.) and peony [
36,
40,
46,
47,
48]. The increase in Mo and V
j, is a specific sign that reflects the blocked electron transfer from the PS II receptor side (Q
A) to the secondary quinone receptor (Q
B), indicating an increased rate of Q
A reduction [
44,
49]. In this experiment, the receptor sides of three different peony varieties were inhibited, with ‘Fengdanbai’ showing more drastic changes before the onset of wilting, and this expression was the most significant among the Mo indicators. This indicates that the PS II receptor side of ‘Fengdanbai’ was more severely damaged and that electron transfer was hindered at the early stages of high-temperature stress compared with the other varieties, while both ‘Huhong’ and ‘Yinhongqiaodui’ showed different degrees of response to high-temperature stress, with ‘Huhong’ responding to high temperatures the most rapidly. High-temperature stress also led to a decrease in Fv/Fm, φ
Eo, and Ψo all of which indicated a decrease in the ability of the PS II receptor side Q
A to transfer electrons, suggesting a decrease in the opening of the active reaction centers, which is consistent with the findings of Jiale Yuan et al. [
50].
While light energy is captured by the photosynthetic machinery for electron transfer, a portion of the energy is dissipated in the form of heat and fluorescence. These three products are in competition with each other, and changes in any one of them will lead to changes in the other two [
42,
51]. Under high-temperature stress, many higher plants turn off the PS II reaction center and deactivate it transiently. It reduces the conversion efficiency of the absorbed light energy by increasing heat dissipation and the dissipation of the fluorescent form of chlorophyll, alleviating the damage caused by excessive light energy transfer to PS II [
46]. Nevertheless, partial inactivation of the reaction center increases the burden on the remaining active reaction centers [
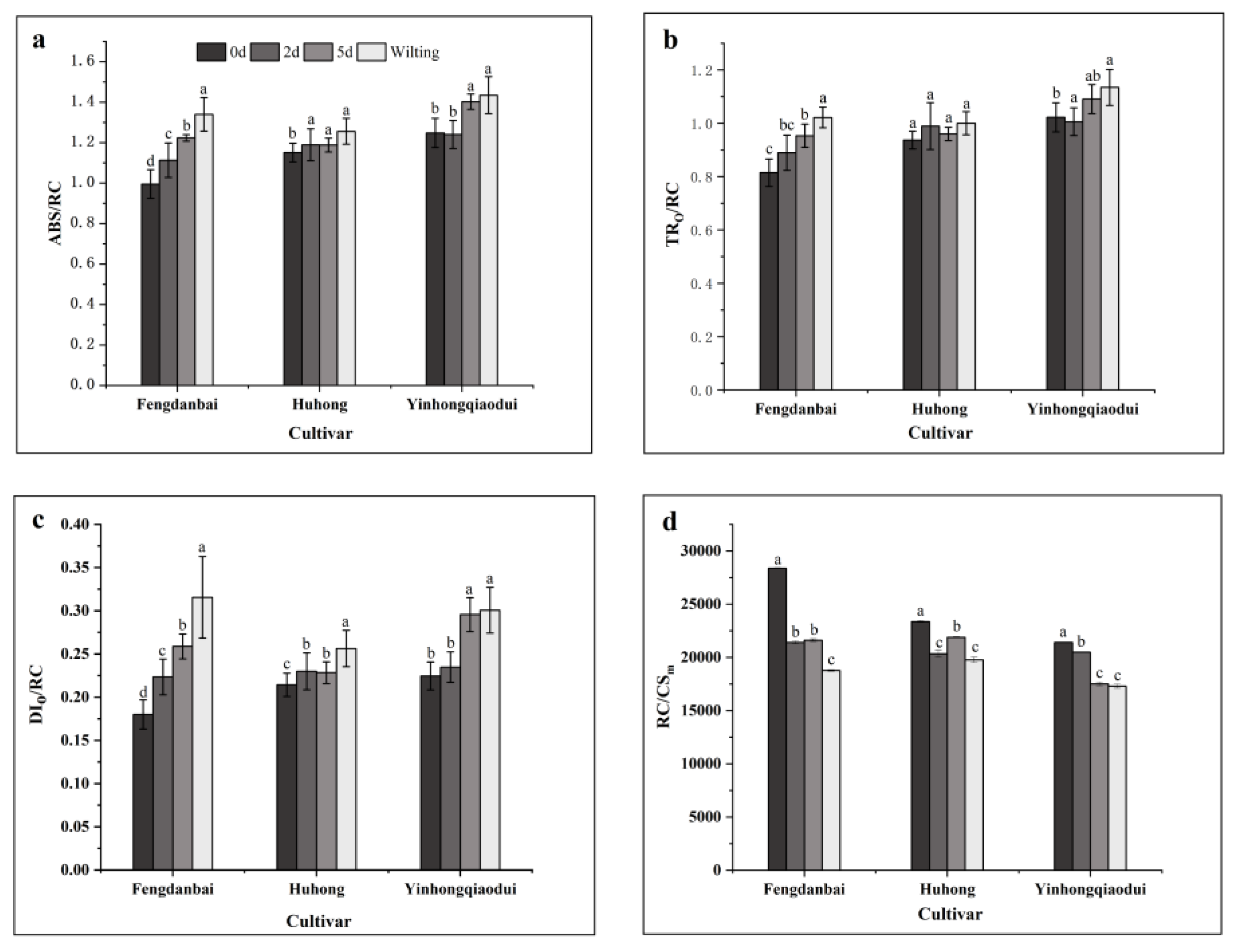
44]. In this experiment, the parameters ABS/RC, TRo/RC, and Dio/RC increased and RC/CS
m decreased, while the total light energy absorption per unit leaf area of the three peony species still showed an overall decreasing trend, indicating that the overall activity of the reaction centers on the leaves of these peony species decreased after the high-temperature stress and the overall light energy absorption decreased, consistent with the results of Yi Sun et al. [
52] on alum root (
Heuchera micrantha Douglas ex Lindl.). Due to the overall damage to the reaction centers on peony leaves under high-temperature stress, the number of active unit reaction centers decreased, resulting in a rise in the amount of light energy required to be absorbed and captured by the remaining unit reaction centers, which is consistent with the findings of Ji Wen et al. [
24]. The increase in ABS/RC may be due to a decreased proportion of active RCs and an increased proportion of absorbed light energy, which may be related to the inactivation of RCs or to an increased antenna size. Previous studies [
53] showed that the RC activity in plants is related to light inhibition, that is, reducing it more than the amount of light absorbed by the plant can result in the plant using its own photosystem. TR
O/RC represent the maximum speed at which the RC captures the excitons [
54]. TR
O/RC and DI
O/RC were significantly higher, indicating that all of the Q
A-values were reduced and could not be reoxidized due to increased temperatures and excessive light exposure. When the reoxidation of Q
A-is suppressed, Q
A-cannot efficiently transfer electrons to Q
B because the inactive center cannot capture a large number of photons, and thus, this excess of photons is considered for energy dissipation [
55]. Therefore, the inhibition of the overall activity of the reaction center also leads to an obstruction of the subsequent electron transfer process, and too much light energy is pooled and stored in the reaction center, resulting in the partial shutdown of the unit’s reaction center and increased light energy dissipation in the antenna complex. This provides slight protection to the subsequent PS I and the whole electron transport chain, but the photosynthetic apparatus was damaged, and the photosynthetic capacity still showed a downward trend. Therefore, the damage degree to the reaction center of PS II unit could be inferred by the increased ratios of ABS/RC, TRo/RC, and Dio/RC indexes.
The number of active PS II reaction centers in ‘Huhong’ showed a significant increase after 5 d of treatment compared after 2 days of treatment, indicating that the inactivation of its PS II reaction centers was self-regulated in response to high-temperature stress, while most of the unit reaction centers of ‘Fengdanbai’ were closed by irreversible damage, probably due to the poor regulation ability of its photosynthetic mechanism under high-temperature stress, and the PS II reaction centers failed to respond to high-temperature stress and to regulate themselves in time after the onset of stress.
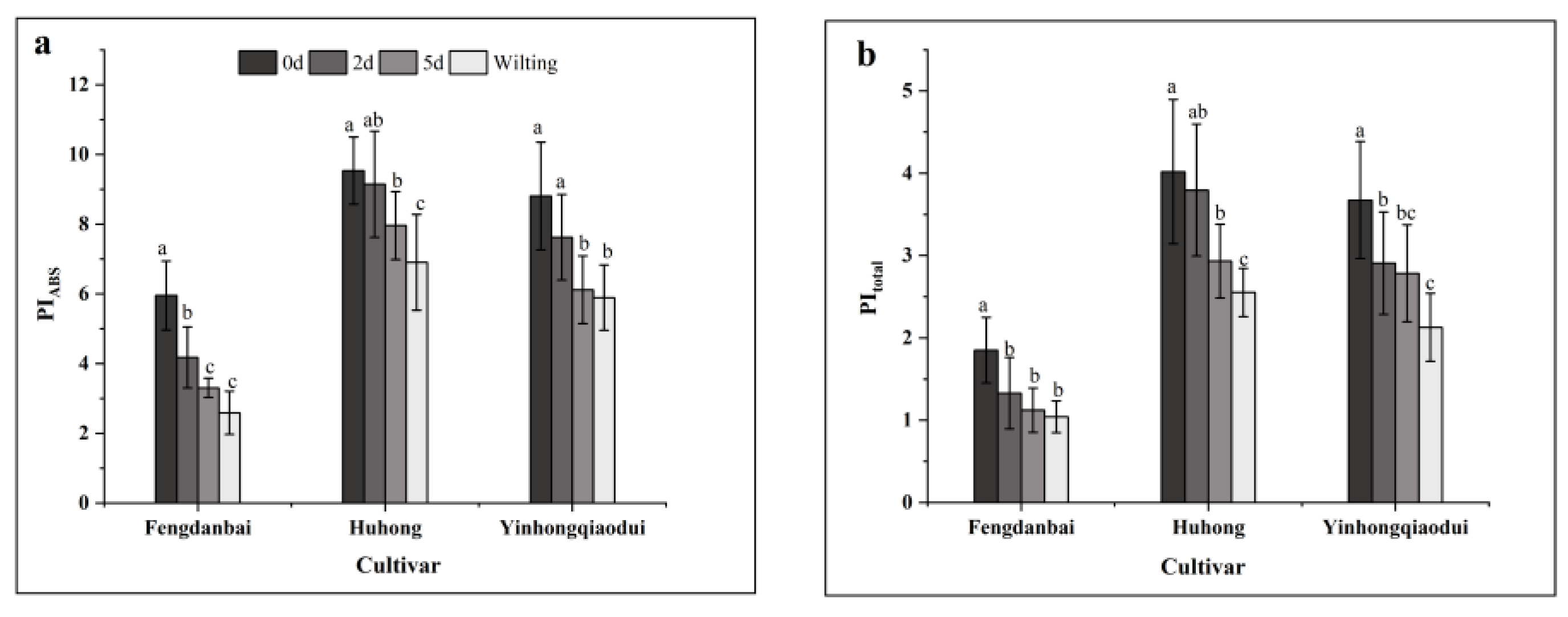
PI
ABS includes three mutually independent parameters: RC/ABS, φ
Po, and Ψo, which reflect the three components of PS II’s active reaction center concentration, the primary photochemical reaction, and electron transfer as well as the photosystem activity in an integrated manner, thus making it more sensitive than Fv/Fm [
24,
44]. Previous studies have shown that PI
ABS and PI
total can accurately respond to the electron transfer between PS I and PS II and to the state of their photosynthetic systems [
56]. Therefore, PI
ABS can be used as a valid fluorescence parameter indicator for different species of peony in response to high-temperature stress [
57]. Zhang et al. [
58] showed that high-temperature stress decreased the PI
ABS of cyanobacteria, mainly due to damage on the donor side of PS II, over-reduction on the acceptor side, and lower energy transfer from the unit’s PS II center. In the present study, the PI
ABS values showed a significant decreasing trend, and high-temperature stress led to a decrease in PS II activity in peony leaves with different degrees of reduction related to its degree of heat tolerance, similar to the results of previous studies [
59].









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}