Abstract
Endophytic bacteria play a crucial role in the life cycle of Spermatophyta, and exploring the beneficial interactions between seeds and endophytic bacteria is an effective strategy for sustainable agricultural development and improved crop productivity. In this study, we employed Illumina Hiseq high-throughput sequencing technology to analyze the amplified fragments of microbial 16S rRNA sequences from melon seeds of MR-1, HH and their F1 generation, which displayed varying levels of resistance to bacterial fruit blotch disease. Initially, we extracted endophytes from eight melon samples and performed sequencing, obtaining an average of 84,758 effective tags per sample. At the 97% similarity level, the number of operational taxonomic units (OTUs) derived from melon seed endophytes ranged from 447 to 492. After filtering out low-level OTUs, we obtained an average of 65,866 valid tags from the eight samples, representing approximately 160 genera and 167 species across eight phyla. It was found that the composition of the endophyte communities in the eight samples was basically similar, and their abundance showed some variability. Furthermore, we utilized the KO and COG databases to functionally predict the genes of endophytes in melon seeds. The analysis revealed a significant enrichment of genes associated with pathways related to protein synthesis, carbohydrate metabolism, substance transport, and metabolite synthesis. These findings greatly contribute to our understanding of endophytic bacteria in melon seeds and serve as a vital reference for the selection and breeding of high-quality melon seeds, as well as the prevention of seed diseases.
1. Introduction
Melon (Cucumis melo L.) is a significant crop globally, being known for its economic value [1]. However, several diseases dramatically reduce the yield and quality of melons, with bacterial fruit blotch being one of the major seed-borne diseases [2]. Seeds contaminated with the Gram-negative bacterium Acidovorax citrulli spread the disease [3]. Chemical and biochemical approaches have been used to decontaminate melon seeds, and attempts have been made to develop resistant varieties [4,5]. Regrettably, there are no commercial cultivars that are resistant to this disease [2,6]. It is coincidental that two cultivars with different levels of resistance to bacterial fruit blotch were found in our experimental field [5]. It is theorized that this may be due to differences in the composition of their seed endophytes.
Plant endophytes can coexist with plants or colonize them for a certain period without causing significant diseases during various stages of plant growth [7]. It is widely recognized that seed endophytes play a crucial role in seed metabolism and disease resistance [8]. Mastretta et al. [9] demonstrated that inoculating cadmium-resistant seed endophytes improved the cadmium resistance of tobacco plants. Puente et al. [10] identified six endophytic bacteria isolated from wheat seeds that inhibited the growth of Fusarium graminearum. Verma et al. [11] identified three endophytes in rice seeds and found that eliminating seed-associated endophytes using antibiotics resulted in stunted rice development and reduced disease resistance. Khalaf et al. [12] isolated 169 endophytic bacteria from melon, watermelon, cucumber, and other cucurbit seeds, among which 118 exhibited antagonistic activity against blight-causing organisms such as Fusarium graminearum, Phytophthora capsici, Pythium aphanidermatum, and powdery mildew of cucurbits. These findings provide theoretical support that endophytic bacteria can prevent bacterial fruit blotch in melon seeds. Currently, endophytic bacteria reported to be effective against fruit blotch in the Cucurbitaceae family include Bacillus spp., Pichia anomala, and Pesudomonas fluorescens [13,14,15].
With the rapid development of next-generation sequencing, DNA sequencing strain identification analysis has become a dominant molecular technique for identifying microbiomes [16,17,18,19]. This method mainly involves PCR amplification and analyzing the 16S rRNA sequences of colonies to confirm the identity of the strains and has been used in the study of seed endophytes. Du et al. [20] utilized Illumina Miseq high-throughput sequencing technology to sequence amplified fragments of 16S RNA V3-V4 and ITS1 regions from four Bupleurum chinense DC. seeds and detected diverse endophyte community structures in these seeds. Yang et al. [21] analyzed the amplified sequences of the 16S rDNA V4 region and ITS1 region of Dendrobium nobile Lindl. seed endophytes using culture-free and high-throughput sequencing technologies to reveal the composition and abundance of endophytes. While numerous studies have employed high-throughput sequencing to investigate the colony structure of seed endophytes [20,21,22,23], there have been relatively few investigations specifically focused on the colony structure of melon seed endophytes.
Previous studies have shown that the melon cultivars MR-1 and HH have different levels of resistance to bacterial fruit blotch [5]. In this study, we used melon MR-1 resistant to bacterial fruit blotch, susceptible melon HH, and the F1 generation seeds of their crosses as experimental materials. Using a combination of traditional microbial culture methods, culture-free techniques, and high-throughput sequencing technology, we explored the bacterial flora structure of different melon seed endophytes and evaluated their relationship with disease resistance. These results provide valuable data for the identification of endophytic bacteria with resistance to bacterial fruit blotch and serve as a theoretical basis for the biological control of melon diseases.
2. Materials and Methods
2.1. Plant Material
Eight melon seed materials with varying levels of bacterial fruit blotch resistance and harvested in different years were included in this study. The details are as shown in Table 1.

Table 1.
Sources of the eight tested materials.
2.2. Cultivable Endophytic Bacteria Isolation and Growth Conditions
Melon seeds were surface-sterilized according to the following procedure. In brief, immersion in sterile distilled water for 1 min was followed by subsequent immersion in 70% (v/v) ethanol (5 min), 5–7 rinses in sterile distilled water, 100% (v/v) ethanol (1 min), and finally drying on sterile filter paper. To obtain homogenates containing endophytes, surface-sterilized seeds were soaked in 70% ethanol for 5 min followed by 1 min in 100% ethanol. Afterwards, hybridization tweezers were used to pick up the internal seeds, which were transferred to a mortar and ground with 3 mL of phosphate-buffered solution. These milled mixtures were added to the poured LB liquid medium and placed in an oscillating incubator and incubated for 19 h at 37 °C with 180 r/min agitation. A total of 200 µL of the culture solution was transferred to LB solid-medium plates to be incubated at a constant temperature for 45 h. To confirm the success of the sterilization process, 200 µL of the water previously used to rinse surface-sterilized seeds was dropped on the LB liquid medium and incubated in accordance with the endophyte culture protocol. These operations were performed on a super-clean bench. The experiment was repeated six times.
2.3. Extraction of DNA
The CTAB extraction method was used as a reference and modified for the extraction of endophytic DNA [24]. For this purpose, nine thoroughly surface-disinfected melon seeds were selected, with every third seed being placed in an EP tube. Three tubes were loaded in total, and steel beads were added to facilitate seed disruption using a plant tissue crusher (40 Hz, 180 s).
Next, 500 µL of TE suspension buffer was added to each tube, followed by the addition of 30 µL of 10% SDS and 50 µL of PD buffer (with the same function as proteinase K). The mixture was thoroughly mixed and incubated at 37 °C for 1 h. Subsequently, 100 µL 5 mol/L NaCl was added and mixed well, and then 100 µL of CTAB solution was added and mixed again. The mixture was heated at 65 °C for 10 min.
To extract the DNA, an equal volume of phenol/chloroform/isoamyl alcohol (25:24:1) was added, followed by centrifugation at 12,000 rpm for five minutes. The supernatant was carefully transferred to a new EP tube. The extraction process was repeated using an equal volume of chloroform/isoamyl alcohol (24:1), and the resulting supernatant was transferred to a new EP tube.
DNA precipitation was performed by adding 0.6 volumes of isopropanol to the supernatant, gently mixing it through inversion, and allowing it to stand at room temperature for 10 min. The mixture was then centrifuged at 12,000 rpm for 10 min, the supernatant was discarded, and the DNA pellet was rinsed with 70% ethanol. Finally, the DNA pellet was air-dried, and the electrophoretic bands were visualized by performing 0.7% agarose gel electrophoresis.
2.4. Amplification of 16S rDNA Gene Fragments
For amplification of the 16S rRNA gene fragments, the extracted DNA was used as the template for PCR with the following primer set: 338F (5′-CCTACGGGAGGCAG-3′) as the upstream primer and 1492R (5′-GGTTACCTTGTTACGACTT-3′) as the downstream primer. The PCR reaction program consisted of the following steps: pre-denaturation at 94 °C for 5 min, denaturation at 94 °C for 1 min, annealing at 58 °C for 1 min, extension at 72 °C for 1 min, and a final extension at 72 °C for 8 min after 30 cycles. The resulting PCR product had an approximate length of 1000 base pairs (bp).
2.5. Library Construction and Sequencing
Following the extraction of total DNA from the samples, primers were designed based on conserved regions. Sequencing adaptors were added to the end of the primers. The target sequences were amplified using PCR, and the resulting products were purified, quantified, and standardized to create a sequencing library. Library quality control (QC) was conducted to ensure the construction of high-quality libraries. Qualified libraries were subsequently sequenced using the Illumina HiSeq 2500 platform.
2.6. Sequencing Data Analysis
According to the overlap relation between PE reads, the paired-end sequence data obtained from Hiseq sequencing were merged into tags of one sequence, and then quality control and filtering were performed for read quality and the merge effect. There were three main steps, as follows. (1) Merging PE reads: FLASH v1.2.7 software was used to merge reads through overlap; the obtained merged sequences were raw tags. (2) Filtering tags: Trimmomatic v0. 33 software was used to filter merged raw tags to obtain high-quality clean tags. (3) Removing Chimera: UCHIME v4. 2 software was used to identify and remove chimeric sequences to obtain effective tags.
Usearch software was then performed for taxonomic annotation for OTUs on the basis of the Silva (Bacteria) taxonomic database [25]. Given that the original OTU clustering results may contain OTUs with very low abundance (species richness less than 0.005%), we needed to filter for low-content OUTs. Following this, QIIME software (http://www.drive5.com/usearch/download.html, accessed on 8 October 2023) was used to generate a species richness table of samples at different taxonomic levels, and then the R language tool was used to a draw community structure graph of samples at different taxonomic levels. Mothur (version v.1.30) software was used to evaluate the Alpha diversity index of samples.
2.7. 16S Functional Gene Prediction
PICRUSt software was used to predict functional gene composition in the samples by comparing species composition information obtained from 16S sequencing data, and then functional differences between samples or groups were analyzed [26]. First, the obtained OTU table needed to be standardized (bacteria from different species and genera have different 16S copies); then, according to the unique greengene id corresponding to each OTU, the KEGG and COG family information related to OTUs could be obtained. Then, the abundance of KEGG and COG information, and the pathway, EC information, and abundance of each type of function were obtained from the KEGG database. At the genus level, the species richness of different samples was tested using the G-TEST in STAMP (large sample: the number of annotated functional genes is greater than 20) and Fisher (small sample: the number of annotated functional genes is less than 20) for significant difference test of samples from pair to pair. T-test was performed for groups from pair to pair, the threshold of p-value was 0.05 (<0.05 means significant).
BugBase was first used to normalize the OTU with the predicted 16S copy number, and then the microbial phenotype was predicted using the provided precomputed documentation [27]. First, for each sample in the biological dataset, the relative abundance of traits was estimated across the entire range of the threshold (0 to 1, increments of 0.01). BugBase then selected the highest coverage threshold for each feature among all samples in the user data. After setting the threshold, BugBase generated the organism-level trait prediction table, which contained the relative abundance of predicted traits for each sample. Based on user-specified metadata, automatic hypothesis testing and visualization were performed for differential trait selection, and group contribution graphs describing the relative abundance of groups with characteristic traits were generated. The output included statistical summary files for non-parametric differential tests (Mann–Whitney U or Kruskal–Wallis).
3. Results
3.1. Assessment of Sequencing Data
The diversity analysis of melon seed endophytes was conducted using Illumina HiSeq sequencing technology. Initially, PCR amplification of the primary screened endophytic bacterial DNA was performed using the universal primers 338F/1492R, resulting in the expected 16S rRNA gene fragments (Figure S1). Subsequently, paired-end sequencing on the Illumina platform was employed to sequence the eight samples, generating a total of 817,053 clean tags after implementing quality control and filtering steps. Each sample produced at least 101,520 clean tags, with an average of 84,758 effective tags per sample (Table 2). The number of operational taxonomic units (OTUs) obtained from melon seed endophytes varied from 447 to 492 at a 97% similarity level (Figure S2). These findings confirm the soundness of the experimental design, high-quality sequencing outcomes, and the provision of informative results.
Table 2.
Statistics for sample sequencing data processing result.
3.2. Counts of Endophytic Bacterial in Melon Seeds
In order to obtain the taxonomic classification information for each OTU, the representative sequences of the OTUs were aligned against a microbial reference database. The community composition of each sample was then determined at different taxonomic levels, including phylum, class, order, family, genus, and species. After filtering out low-level OTUs, an average of 65,866 effective tags were obtained from the eight samples (Table S1). These tags represented approximately 160 genera and 167 species from eight phyla, namely Tenericutes, Spirochaetes, Actinobacteria, Bacteroidetes, Proteobacteria, thick-cell-wall Firmicutes, Epsilonbacteraeota, and Kiritimatiellaeota (Figure 1a, Table S2).
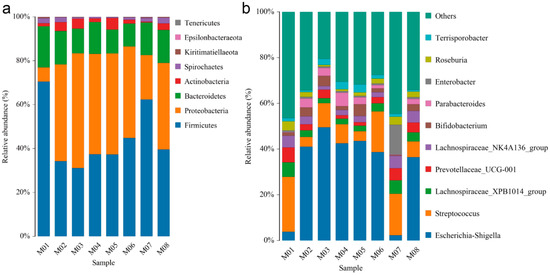
Figure 1.
Distribution of endophyte community in eight melon seeds. (a) Dominance phylum and (b) dominant genus of endophyte community in melon seeds.
The dominant phylum varied among the samples, with Proteobacteria dominating in M02-M06, Tenericutes in M07, and Kiritimatiellaeota in M01. Among the top ten genera in terms of abundance, Escherichia-Shigella, Lachnospiraceae_XPB1014_group, Streptococcus, Lachnospiraceae_NK4A136_group, Prevotellaceae_UCG-001, Bifidobacterium, Parabacteroides, Enterobacter, Roseburia, and Terrisporobacter were observed (Figure 1b). Notably, Escherichia-Shigella accounted for more than 35% of the endophytes in all materials except for M01 and M07 (Table S3).
3.3. Species Diversity Analysis for Melon Seed Endophytes
Alpha diversity is a measure of species richness and diversity within an individual sample, assessed using various indices such as Chao1, Ace, Shannon, and Simpson [8]. The Shannon and Simpson indices specifically evaluate species diversity and are influenced by the species richness within a sample community [28]. In our study, we found that when the sequencing throughput reached 10 kb, neither the number of OTUs nor the Shannon index significantly increased, indicating that our data adequately captured the species diversity in the samples (Figure S3).
The results reveal that the eight materials exhibited Shannon’s indices ranging from 2.6377 to 4.273 and Simpson’s indices ranging from 0.0585 to 0.2618. Moreover, the sample coverages were above 99% (Table 3). Notably, compared to the other six materials, M01 and M07 displayed greater species diversity in their seed endophytes.
Table 3.
Alpha diversity index statistics.
3.4. Prediction and Analysis of Functional Genes from Melon Seed Endophytes
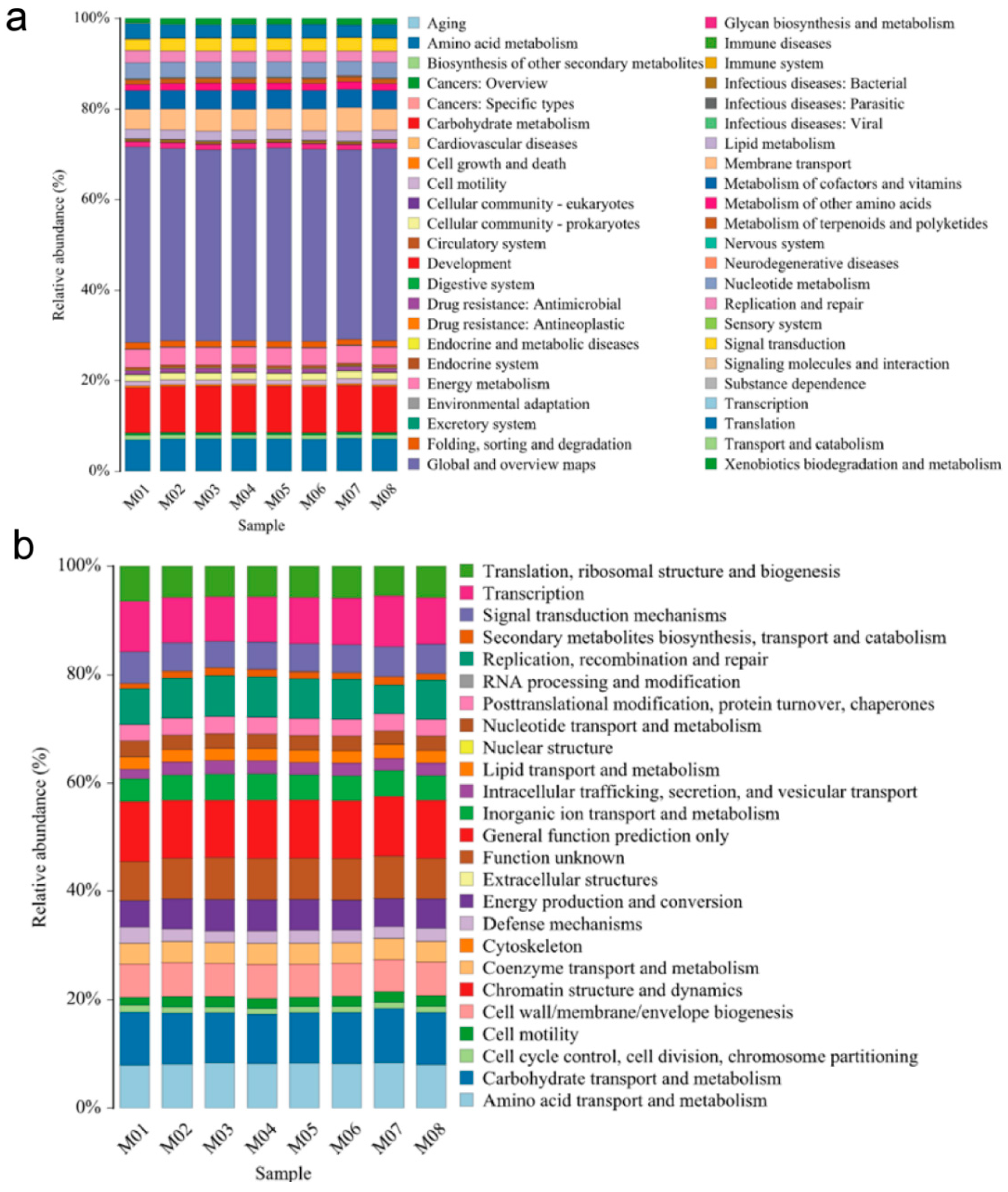
Differences in KEGG metabolic pathways can provide insights into the changes and adaptations of microbial community functional genes to environmental variations [29]. Our results demonstrate that the abundance of KEGG metabolic pathways in the endophytic bacteria of melon seeds across the eight samples was similar. The majority of the pathways were associated with metabolism-related information (41.83–43.21%), followed by carbohydrate metabolism (9.92–10.25%), amino acid metabolism (6.67–6.87%), and transmembrane transport (4.41–5.26%). Various subspecies also exhibited resistance-related information against bacteria, with an abundance range of 0.84–1.04%. Additionally, there was low-abundance information related to bacterial infectious diseases (0.43–0.54%) and parasitic infectious diseases (0.06–0.08%) (Figure 2a).
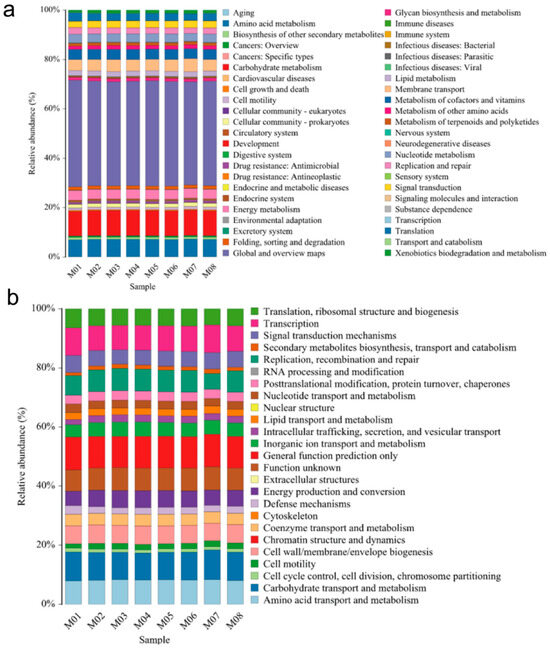
Figure 2.
Functional prediction of endophytic bacteria in melon seeds. (a) KEEG and (b) COG analysis for gene function at the OTU level from 8 melon seed endophytes.
The COG (Clusters of Orthologous Groups of proteins) database is commonly employed for functional classification of proteins in prokaryotes [30]. In our study, the COG functional classification revealed that the functions of melon seed endophytic bacteria mainly involved substance and energy transport, catabolism, cell biological behavior occurrence, and regulation. The descending order of functional abundance (≥1% in each subsample) included general functional predictor genes (10.58–11.11%); carbohydrate transport and metabolism (9.08–10.01%); transcription (8.27–9.36%); amino acid transport and metabolism (7.98–8.41%); function unknown (7.23–7.83); replication, recombination, and repair (5.33–7.55%); translation, ribosome structure, and biogenesis (5.48–6.49%); cell wall, cell membrane, and envelope biogenesis (5.90–6.28%); signal transduction mechanism (4.87–5.71%); energy production and turnover (4.87–5.73%); inorganic ion transport and metabolism (4.20–4.90%); coenzyme transport and metabolism (3.83–3.93%); posttranslational modification, protein turnover, and concomitant protein (2.98–3.18%); nucleotide trafficking and metabolism (2.52–2.52%); intracellular trafficking and vesicular trafficking mechanisms (2.28–2.57%); intracellular trafficking and trafficking (2.77%); cell viability (1.43–2.04%); cell cycle control, cell division, and chromosome segmentation (1.08–1.36%); and biosynthesis, trafficking, and catabolism of secondary metabolites (1.00–1.52%) (Figure 2b).
Comparing the KO and COG databases, the gene function prediction results indicate that most of the information detected from the eight melon seed endophytic bacteria corresponded to beneficial functional processes.
3.5. Structural Analysis of Endophytic Community in Melon Seeds
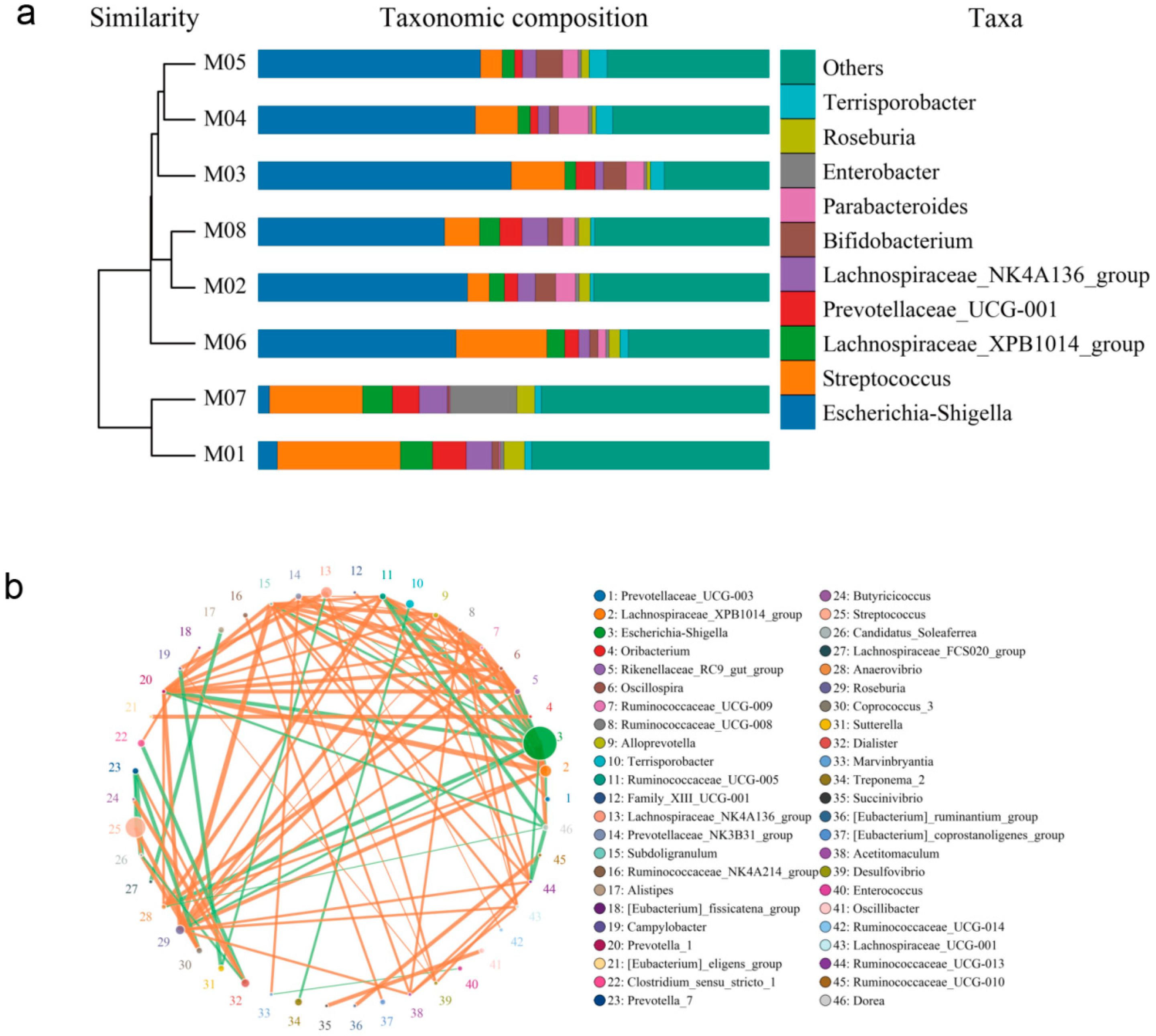
Cluster analysis was employed to investigate the differences in endophytic bacterial communities among different melon seed samples. As shown by the cluster analysis, the endophyte genera composition of M01 and M07 was similar, while the genera of M02, M03, M04, M05, M06, and M08 were more closely related to each other (Figure 3a). Notably, the distribution of endophyte genera in M01 and M07 was slightly different compared to that in the other six samples (Table S4). It is interesting to note that the parents of M01 and M07 are known to be resistant to fruit blotch disease.
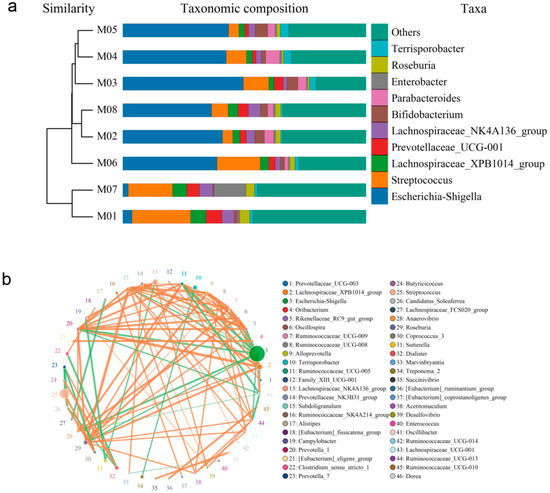
Figure 3.
Correlation analysis of endophytic community of melon seeds. (a) Combined drawing of clustering tree and histogram; (b) species network at genus level.
To further examine the composition of endophytic bacterial communities in the melon seed samples, PCoA analysis based on Bray–Curtis distance was performed. At the genus level, the first principal component (PC1) accounted for 79.15% of the variation, while the second principal component (PC2) accounted for 9.46%. These two principal components were the primary factors influencing the structure of the endophytic bacterial community in the melon seeds. PC1 exhibited a significantly greater contribution rate compared to PC2. Based on the PCoA results, the eight seed samples were clustered into five categories. Specifically, the M01 and M07 samples, M02 and M08 samples, and M03 and M04 samples showed relatively small distances between them, while M05 and M06 samples formed separate clusters (Figure S4). This suggests that extended storage time may lead to a decrease in the beneficial population associated with fruit blotch resistance in melon seeds. Additionally, strong correlations were observed between certain endophytic communities in melon seeds, such as Escherichia-Shigella and Lachnospiraceae_XPB1014_group, which displayed a negative correlation (Figure 3b, Table S5).
4. Discussion
In this study, we collected melon seeds of the cultivars MR-1 and HH, which have exhibited different levels of resistance to bacterial fruit blotch in different years, and F1-generation seeds produced from their combination in order to analyze the endophyte community composition of melon seeds and the relationship between disease resistance and the endophytes of melon seeds. The Illumina high-throughput sequencing technique was employed to analyze the endophytic bacterial community structure and diversity in eight melon seed samples, resulting in the identification of 8 phyla, 14 classes, 21 orders, 48 families, 160 genera, 167 species, and 492 epiphytic bacterial OTUs (Figure 1a,b; Tables S1 and S3). Functional predictions revealed a significant number of genes related to protein synthesis, carbohydrate metabolism, substance transport, and metabolite synthesis, which may collectively regulate growth development and energy metabolism in melon (Figure 2a,b).
The abundance of endophytic bacteria in plant seeds is influenced by several factors, including seed genotype [31], harvest season [32], and storage time [33]. According to the Alpha diversity index, we found that the diversity of the resistance material M01 and the sensitive material M05 harvested in 19 years were slightly higher than those harvested in previous years (Table 3). Furthermore, the diversity of seed endophytes was significantly higher in M01 and M07 (with MR as the mother plant) than in M05 and M08 (with HH as the mother plant) harvested in 2019 (Figure 3a). In order to create a more congenial growth environment, seeds tend to spread more beneficial endophytes to the next generation, so seeds with a richer endophyte community structure may have better vigor and disease resistance [34]. Generally, the proportional distribution of endophytic colonies in melon seeds at the phylum level is similar to that of other species, yet it varies considerably at the genus level [20,35,36]. The identification results for the eight samples also partially reveal differences between resistant and susceptible materials at the genus level (Figure 3a, Table S4). Our results show that the fruit-blotch-resistant material M01 contained 24.02% Streptococcus, 6.31% Lachnospiraceae_XPB1014_group, and 3.82% Escherichia-Shigella, while the fruit-blotch-sensitive material M04 contained 42.52% Escherichia-Shigella, 8.29% Streptococcus, and 2.44% Lachnospiraceae_XPB1014_group. Previous studies showed that some streptococci can produce antibacterial metabolites to enhance the ability of symbiotic plants against bacterial diseases [37,38]. In our study, in the endophytic communities in the eight samples, both beneficial and harmful bacteria were present (Figure 1b, Tables S3 and S4). Escherichia-Shigella is a pathogenic genus causing plant diseases and is capable of damaging the leaves and stems [39,40]. Lachnospira is a potentially beneficial bacterium that can provide energy support to host cells [41,42]. Beneficial and harmful endophytes are widely present in the stem, leaf, and seeds of plants [5,43]; however, the relationship between the bacterial community in melon seed and melon resistance to bacterial fruit blotch still needs to be further explored.
Our study presents a comprehensive analysis of the endophytic community in eight melon seeds, including both structural and functional aspects. The results consistently indicate that the majority of bacteria found in the seed endophyte community are beneficial, with predicted gene functions primarily associated with the growth and development of melons. Moreover, our findings reveal significant interactions among the bacteria within the seed endophyte, suggesting the presence of specific bacterial strains that may have antagonistic effects against melon fruit blotch disease (Table S4). Previous researchers had already identified some endophytes related to fruit blotch disease through microbial co-culture and morphological characterization [14,15], and the combination of our results can be used to further screen endophytes related to disease resistance at the macroscopic level. Considering these results, exploring the potential of reintroducing these bacteria into the endophytic community becomes an attractive avenue for enhancing melon’s disease resistance. Overall, our study provides valuable insights into plant endophytes while establishing a theoretical foundation for disease resistance breeding in melons.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9111195/s1. Figure S1: PCR amplification analysis of the 16S rRNA gene of endophytic bacteria from melon seeds. (A) and (B) are the electrophoresis plots before and after template dilution, respectively. Primers are universal primers 338F/1492R; NEG represents negative control. Figure S2: distribution map of OTU count of each sample. Figure S3: sample rarefaction curve. Note: X-axis represents the number of sequences picked randomly, Y-axis represents the OTU number based on the clustering of these sequences. Each curve stands for a sample, and they are marked by different colors. Figure S4: principal coordinate analysis of endophytic bacteria of 8 melon samples seeds at phylum level. Table S1: sample ID mapping table. Table S2: statistical table of per-level tags of samples. Table S3: statistical table of per-level species of samples. Table S4: species distribution statistics in eight samples at the genus level. Table S5: edge properties table.
Author Contributions
Conceived and designed for study: K.Z., Y.L. and L.X.; experimental work: K.Z., Z.W., Y.D. and M.F.; collected data and literature: K.Z. and Y.L.; drafted the manuscript: K.Z.; corrected the manuscript: Y.L. and L.X.; project fund management: K.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (32060688).
Data Availability Statement
Unpublished data.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chikh-Rouhou, H.; Abdedayem, W.; Solmaz, I.; Sari, N.; Garcés-Claver, A. Melon (Cucumis melo L.): Genomics and Breeding. In Smart Plant Breeding for Vegetable Crops in Post-Genomics Era; Springer Nature Singapore: Singapore, 2023; pp. 25–52. [Google Scholar]
- Klomchit, A.; Calderin, J.D.; Jaidee, W.; Watla-iad, K.; Brooks, S. Napthoquinones from Neocosmospora sp.—Antibiotic Activity against Acidovorax citrulli, the Causative Agent of Bacterial Fruit Blotch in Watermelon and Melon. J. Fungi. 2021, 7, 370. [Google Scholar]
- Burdman, S.; Walcott, R. Acidovorax citrulli: Generating basic and applied knowledge to tackle a global threat to the cucurbit industry. Mol. Plant Pathol. 2012, 13, 805–815. [Google Scholar] [PubMed]
- Wang, H.; Qian, C.; Jiang, H.; Liu, S.; Yang, D.; Cui, J. Visible-Light-Driven Zinc Oxide Quantum Dots for the Management of Bacterial Fruit Blotch Disease and the Improvement of Melon Seedlings Growth. J. Agric. Food Chem. 2023, 71, 2773–2783. [Google Scholar] [CrossRef] [PubMed]
- Ning, X.F. Identification of resistance to Acidovorax citrulli among different melon accessions and detection of QTLs for bacterial fruit blotch and downy mildew resistance in melon. Ph.D. Thesis, Xinjiang University, Xinjiang, China, 2018. [Google Scholar]
- Bahar, O.; Kritzman, G.; Burdman, S. Bacterial fruit blotch of melon: Screens for disease tolerance and role of seed transmission in pathogenicity. Eur. J. Plant Pathol. 2009, 123, 71–83. [Google Scholar] [CrossRef]
- Wani, Z.A.; Ashraf, N.; Mohiuddin, T.; Riyaz-Ul-Hassan, S. Plant-endophyte symbiosis, an ecological perspective. Appl Microbiol Biotechnol. 2015, 99, 2955–2965. [Google Scholar] [PubMed]
- Thomas, P.; Sahu, P.K. Vertical transmission of diverse cultivation-recalcitrant endophytic bacteria elucidated using watermelon seed embryos. Front. Microbiol. 2021, 12, 635810. [Google Scholar]
- Mastretta, C.; Taghavi, S.; van der Lelie, D.; Mengoni, A.; Galardi, F.; Gonnelli, C.; Vangronsveld, J. Endophytic bacteria from seeds of Nicotiana tabacum can reduce cadmium phytotoxicity. Int. J. Phytoremed. 2009, 11, 251–267. [Google Scholar] [CrossRef]
- Puente, M.E.; Li, C.Y.; Bashan, Y. The development of cactus seedlings can be improved by the presence of endophytic bacteria in cactus seeds. Environ. Exp. Bot. 2009, 66, 402–408. [Google Scholar] [CrossRef]
- Verma, S.; Kingsley, K.; Irizarry, I.; Bergen, M.; Kharwar, R.N.; White, J. Seed-vectored endophytic bacteria modulate development of rice seedlings. J. Appl. Microbiol. 2017, 122, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, E.M.; Raizada, M.N. Bacterial Seed Endophytes of Domesticated Cucurbits Antagonize Fungal and Oomycete Pathogens Including Powdery Mildew. Front. Microbiol. 2018, 9, 42. [Google Scholar]
- Fessehaie, A.; Walcott, R.R. Biological Control to Protect Watermelon Blossoms and Seed from Infection by Acidovorax avenae subsp. citrulli. Phytopathology 2005, 95, 413–419. [Google Scholar]
- Jiang, C.-H.; Wu, F.; Yu, Z.-Y.; Xie, P.; Ke, H.-J.; Li, H.-W.; Yu, Y.-Y.; Guo, J.-H. Study on screening and antagonistic mechanisms of Bacillus amyloliquefaciens 54 against bacterial fruit blotch (BFB) caused by Acidovorax avenae subsp. citrulli. Microbiol. Res. 2015, 170, 95–104. [Google Scholar] [CrossRef]
- de Melo, E.A.; Mariano, R.d.L.R.; Laranjeira, D.; dos Santos, L.A.; Gusmão, L.d.O.; de Souza, E.B. Efficacy of Yeast in the Biocontrol of Bacterial Fruit Blotch in Melon Plants. Trop. Plant Pathol. 2015, 40, 56–64. [Google Scholar] [CrossRef]
- Edwards, J.; Santos-Medellín, C.; Sundaresan, V. Extraction and 16S rRNA sequence analysis of microbiomes associated with rice roots. Bio-Protocol 2018, 8, e2884. [Google Scholar] [CrossRef] [PubMed]
- Majeed, A.; Kaleem Abbasi, M.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar]
- Qaisrani, M.M.; Mirza, M.S.; Zaheer, A.; Malik, K.A. Isolation and identification by 16s rRNA sequence analysis of achromobacter, azospirillum and rhodococcus strains from the rhizosphere of maize and screening for the beneficial effect on plant growth. Pak. J. Agric. Sci. 2014, 51, 91–99. [Google Scholar]
- Oliveira, C.; Shakiba, E.; North, D.; McGraw, M.; Ballard, E.; Barrett-D’amico, M.; Glazko, G.; Rahmatallah, Y. 16S rRNA gene-based metagenomic analysis of rhizosphere soil bacteria in arkansas rice crop fields. Agronomy 2022, 12, 222. [Google Scholar] [CrossRef]
- Du, K.; Geng, Y.; Liu, L.; Gao, D.M. Community structure and diversity of endophytes of Bupleurum chinense DC. seeds. Biotic Resour. 2022, 44, 36–44. [Google Scholar]
- Yang, L.; Wu, Q.; Gao, Y.; Chen, Q.; Wang, Y.; Niu, X.; Weng, Q. Analysis of the composition of endophytic community of Dendrobium nobile Lindl. seeds based on high-throughput sequencing. Seed 2020, 39, 94–98. [Google Scholar]
- Gerna, D.; Clara, D.; Allwardt, D.; Mitter, B.; Roach, T. Tailored media are key to unlocking the diversity of endophytic bacteria in distinct compartments of germinating seeds. Microbiol. Spectr. 2022, 10, e00172-22. [Google Scholar]
- Sha, W.; Hong, D.; Che, Y.; Xue, Y.; Kong, Y.; Yi, X.; Liu, B. Differences in Root Endophytic Bacterial Communities of Chinese Cork Oak (Quercus variabilis) Seedlings in Different Growth Years. Forests 2023, 14, 1489. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Yu, J.; Wang, L.; Zhou, S. A modified CTAB protocol for plant DNA extraction. Chin. Bull. Bot. 2013, 48, 72. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Knights, D. BugBase predicts organism-level microbiome phenotypes. BioRxiv 2017, 133462, 1–19. [Google Scholar]
- Roswell, M.; Dushoff, J.; Winfree, R. A conceptual guide to measuring species diversity. Oikos 2021, 130, 321–338. [Google Scholar] [CrossRef]
- Hemme, C.L.; Deng, Y.; Gentry, T.J.; Fields, M.W.; Wu, L.; Barua, S.; Barry, K.; Tringe, S.G.; Watson, D.B.; He, Z.; et al. Metagenomic insights into evolution of a heavy metal-contaminated groundwater microbial community. ISME J. 2010, 4, 660–672. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Wali, P.R.; Helander, M. Genetic compatibility determines endophyte-grass combinations. PLoS ONE 2010, 5, e11395. [Google Scholar] [CrossRef]
- Hume, D.; Schmid, J.; Rolston, M.; Vijayan, P.; Hickey, M. Effect of climatic conditions on endophyte and seed viability in stored ryegrass seed. Seed Sci. Technol. 2011, 39, 481–489. [Google Scholar] [CrossRef]
- Welty, R.E. Influence of moisture content, temperature, and length of storage on seed germination and survival of endophytic fungi in seeds of tall fescue and perennial ryegrass. Phytopathology 1987, 77, 893. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Han, S.; Liu, C.; Liu, F. Effect of two seaweed polysaccharides on intestinal microbiota in mice evaluated by illumina PE250 sequencing. Int. J. Biol. Macromol. 2018, 112, 796–802. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Ayangbenro, A.S.; Babalola, O.O. Metagenomic profiling of the community structure, diversity, and nutrient pathways of bacterial endophytes in maize plant. Antonie Van Leeuwenhoek 2020, 113, 1559–1571. [Google Scholar] [PubMed]
- Purushotham, N.; Jones, E.; Monk, J.; Ridgway, H. Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata. Plants 2020, 9, 156. [Google Scholar] [CrossRef]
- Park, K.; Paul, D.; Kim, E.; Kloepper, J.W. Hyaluronic acid of Streptococcus sp. as a potent elicitor for induction of systemic resistance against plant diseases. World J. Microbiol. Biotechnol. 2007, 24, 1153–1158. [Google Scholar] [CrossRef]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef]
- Brandl, M.T. Plant lesions promote the rapid multiplication of Escherichia coli O157: H7 on postharvest lettuce. Appl. Environ. Microbiol. 2008, 74, 5285–5289. [Google Scholar] [CrossRef]
- Simko, I.; Zhou, Y.; Brandl, M.T. Downy mildew disease promotes the colonization of romaine lettuce by Escherichia coli O157: H7 and Salmonella enterica. BMC Microbiol. 2015, 15, 19. [Google Scholar] [CrossRef]
- Muralidharan, J.; Galiè, S.; Hernández-Alonso, P.; Bulló, M.; Salas-Salvadó, J. Plant-based fat, dietary patterns rich in vegetable fat and gut microbiota modulation. Front. Nutr. 2019, 6, 157. [Google Scholar]
- Ajmeer, A.J.; Selvarajan, R.; Mearns, K.; Pandian, J. Unveiling the Bacterial Communities and Its Potential Agricultural Applications from Organic Manure (Panchagavya) Using Targeted Amplicon Analysis. Pol. J. Environ. Stud. 2013, 6, 1–9. [Google Scholar] [CrossRef]
- Mushtaq, S.; Shafiq, M.; Tariq, M.R.; Sami, A.; Nawaz-Ul-Rehman, M.S.; Bhatti, M.H.T.; Haider, M.S.; Sadiq, S.; Abbas, M.T.; Hussain, M.; et al. Interaction between bacterial endophytes and host plants. Front. Plant Sci. 2023, 13, 1092105. [Google Scholar] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).