
Effects of Root and Foliar Application of Corn Steep Liquor on Pepper Plants: A Physiological, Nutritional, and Morphological Study

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Condicitons
2.2. Experimental Design
2.3. Measurements of Gas Exchange and Chlorophylls Fluorescence
2.3.1. Photosynthetic Efficiency
2.3.2. Fluorescence of Chlorophyll a
2.4. Analysis of Plant Samples
2.4.1. Concentrations of Oxidative Indicators (MDA, H2O2, and O2−)
2.4.2. Hormonal Profile
2.4.3. Aminogram
2.4.4. Enzymatic Activities Involved in Nitrogen Metabolism
2.4.5. Concentrations of Total Nutrients and Nitrate and Nitrogen Use Efficiency
2.5. Statistical Analysis
3. Results
3.1. Biomass and Leaf Area
3.2. Gas Exchange Parameters
3.3. Chl a Fluoresence
3.4. Study of Oxidative Stress
3.5. Hormonal Profile
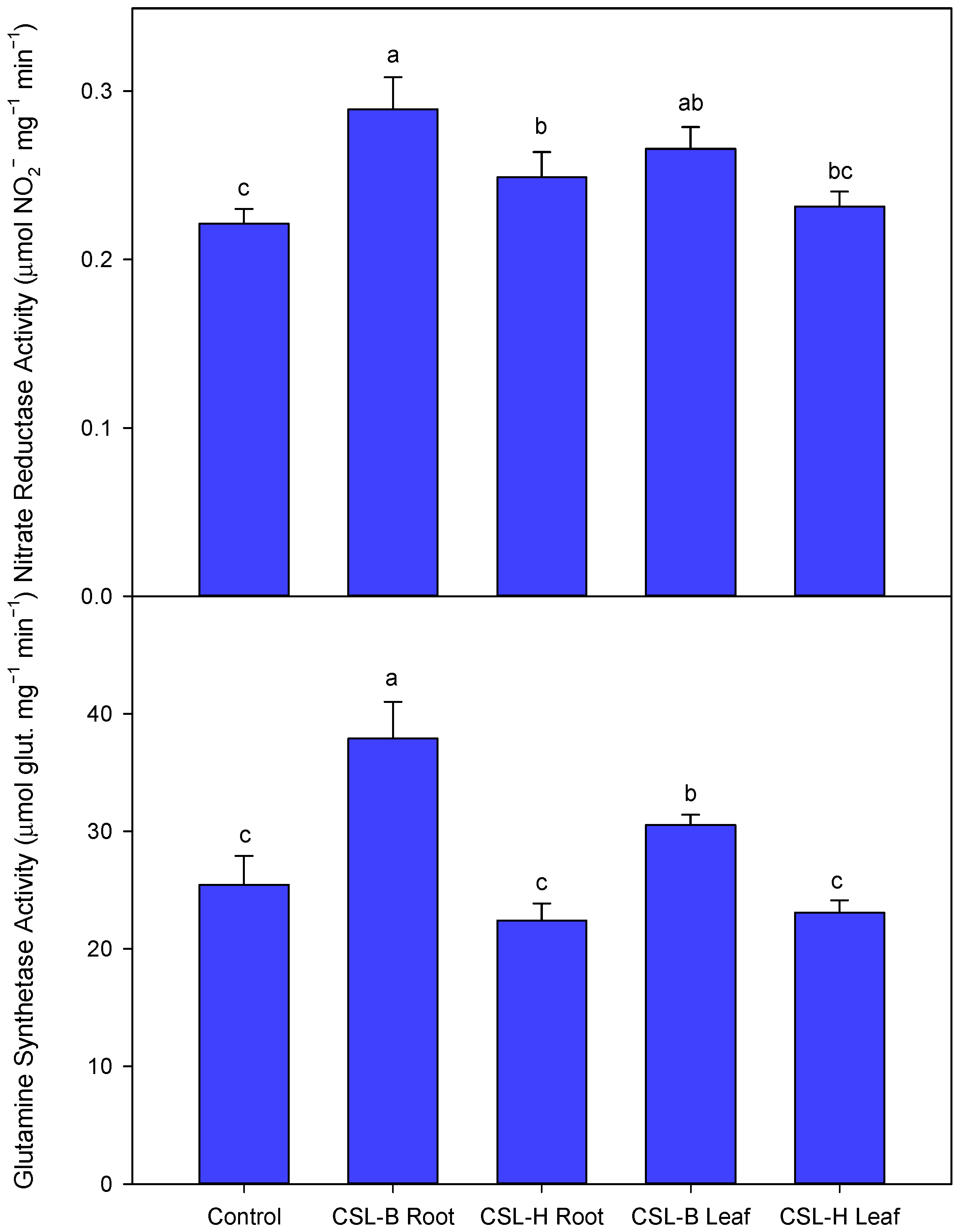
3.6. Enzymatic Activities Associated with Nitrogen Metabolism (Nitrate Reductase, NR, and Glutamine Synthetase, GS)
3.7. Aminogram
- Root application of CSL-B produced the highest values of proline, serine, and threonine;
- Root application of CSL-H gave the highest concentrations of asparagine, aspartate, isoleucine, leucine, phenylalanine, proline, valine, and cysteine;
- Foliar application of CSL-B produced the highest values of asparagine, phenylalanine, and tryptophan;
- The foliar CSL-H treatment produced the highest values of alanine, glycine, histidine, lysine, phenylalanine, serine, and tryptophan.
3.8. Concentrations of Nutrients
4. Discussion
- An increase in the hormones AIA, tZ, GA1, GA3, and GA4 (Table 5).
- An increase in the foliar concentrations of P, K, Ca, Mg, and S of 19, 19, 16, 41, and 15%, respectively, without reaching values toxic for plants (Table 7).
- Changes in the aminogram, with increases in the concentrations of alanine, proline, and tryptophan of more than 30%, and a decrease of more than 30% for tyrosine and methionine, for the 4 CSL treatments. There were also differences among the treatments, with the concentrations of 7, 10, 4, and 9 amino acids increasing by more than 30% for the treatments CSL-B Root, CSL-H Root, CSL-B Foliar, and CSL-H Foliar, respectively (Table 6).
- Increased NR and glutamine synthetase activity in the CSL-B Root treatment.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cornejo-Villegas, M.; Rincón-Londoño, N.; Real-López, D.; Rodríguez-García, M.E. The efect of Ca2+ ions on the pasting, morphological, structural, vibrational, and mechanical properties of corn starch-water system. J. Cereal Sci. 2018, 79, 174–182. [Google Scholar] [CrossRef]
- Martínez-Arcos, A.; Moldes, A.B.; Vecino, X. Adding value to secondary streams of corn wet milling industry. CyTA-J. Food 2021, 19, 675–681. [Google Scholar] [CrossRef]
- Lawford, H.G.; Rousseau, J.D. Corn steep liquor as a cost-efective nutrition adjunct in high-performance Zymomonas ethanol fermentation. Appl. Biochem. Biotech. 1997, 63–65, 287–304. [Google Scholar] [CrossRef]
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.N.; Huhnke, R.L. Ethanol production from Syngas by Clostridium strain P11 using corn steep liquor as a nutrient replacement to yeast extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- Hofer, A.; Hauer, S.; Kroll, P.; Fricke, J.; Herwig, C. In-depth characterization of the raw material corn steep liquor and its bioavailability in bioprocesses of Penicillium chrysogenum. Process Biochem. 2018, 70, 20–28. [Google Scholar] [CrossRef]
- Sekoai, P.T.; Ayeni, A.O.; Daramola, M.O. Parametric optimization of citric acid production from apple pomace and corn steep liquor by a wild type strain of Aspergillus niger: A response surface methodology approach. Int. J. Eng. Res. Afr. 2018, 36, 98–113. [Google Scholar] [CrossRef]
- López-Prieto, A.; Martínez-Padrón, H.; Rodríguez-López, L.; Moldes, A.B.; Cruz, J.M. Isolation and characterization of a microorganism that produces biosurfactants in corn steep water. CyTA-J. Food 2019, 17, 509–516. [Google Scholar] [CrossRef]
- Hull, S.R.; Yang, B.Y.; Venzke, D.; Kulhavy, K.; Montgomery, R. Composition of corn steep water during steeping. J. Agr. Food Chem. 1996, 44, 1857–1863. [Google Scholar] [CrossRef]
- Shin, H.D.; McClendon, S.; Le, T.; Taylor, F.; Chen, R.R. A complete enzymatic recovery of ferulic acid from corn residues with extracellular enzymes from Neosartorya spinosa NRRL 185. Biotechnol. Bioeng. 2006, 95, 1108–1115. [Google Scholar] [CrossRef]
- Amiri, A.; Bundur, Z.B. Use of corn-steep liquor as an alternative carbon source for biomineralization in cement-based materials and its impact on performance. Constr. Build. Mater. 2018, 165, 655–662. [Google Scholar] [CrossRef]
- Obayori, O.S.; Ilori, M.O.; Adebusoye, S.A.; Oyetibo, G.O.; Omotayo, A.E.; Amund, O.O. Efects of corn steep liquor on growth rate and pyrene degradation by Pseudomonas strains. Curr. Microbiol. 2010, 60, 407–411. [Google Scholar] [CrossRef]
- Obayori, O.S.; Salam, L.B.; Anifowoshe, W.T.; Odunewu, Z.M.; Amosu, O.E.; Ofulue, B.E. Enhanced degradation of petroleum hydrocarbons in corn-steep-liquor-treated soil microcosm. Soil Sediment Contam. 2015, 24, 731–743. [Google Scholar] [CrossRef]
- Hamedani, S.R.; Rouphael, Y.; Colla, G.; Colantoni, A.; Cardarelli, M. Biostimulants as a tool for improving environmental sustainability of greenhouse vegetable crops. Sustainability 2020, 12, 5101. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Findura, P.; Treder, K. Modification of yield and fiber fractions biosynthesis in Phaseolus vulgaris by treatment with biostimulants containing amino acids and seaweed extract. Agronomy 2020, 10, 1338. [Google Scholar] [CrossRef]
- Caradonia, F.; Ronga, D.; Flore, A.; Barbieri, R.; Moulin, L.; Terzi, V.; Francia, E. Biostimulants and cherry rootstock increased tomato fruit yield and quality in sustainable farming systems. Ital. J. Agron. 2020, 15, 121–131. [Google Scholar] [CrossRef]
- Poberezny, J.; Szczepanek, M.; Wszelaczynska, E.; Prus, P. The quality of carrot after field biostimulant application and after storage. Sustainability 2020, 12, 1386. [Google Scholar] [CrossRef]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The use of a plant-based biostimulant improves plant performances and fruit quality in tomato plants grown at elevated temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hort. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Zarzecka, K.; Gugala, M.; Mystkowska, I.; Sikorska, A. Total and true protein content in potato tubers depending on herbicides and biostimulants. Agronomy 2020, 10, 1106. [Google Scholar] [CrossRef]
- Conesa, M.R.; Espinosa, P.J.; Pallarés, D.; Pérez-Pastor, A. Influence of plant biostimulant as technique to harden citrus nursery plants before transplanting to the field. Sustainability 2020, 12, 6190. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Testolin, R.; Zanotelli, D.; Andreotti, C. Effect of biostimulants on apple quality at harvest and after storage. Agronomy 2020, 10, 1214. [Google Scholar] [CrossRef]
- Dalal, A.; Bourstein, R.; Haish, N.; Shenhar, I.; Wallach, R.; Moshelion, M. Dynamic physiological phenotyping of drought-stressed pepper plants treated with “productivity-enhancing” and “survivability-enhancing” biostimulants. Front. Plant Sci. 2019, 10, 905. [Google Scholar] [CrossRef]
- Cozzolino, E.; Giordano, M.; Fiorentino, N.; El-Nakhel, C.; Pannico, A.; Di Mola, I.; Mori, M.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Appraisal of biodegradable mulching films and vegetal-derived biostimulant application as eco-sustainable practices for enhancing lettuce crop performance and nutritive value. Agronomy 2020, 10, 427. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Hellequin, E.; Monard, C.; Chorin, M.; Lebris, N.; Daburon, V.; Klarzynski, O.; Binet, F. Responses of active soil microorganisms facing to a soil biostimulant input compared to plant legacy effects. Sci. Rep. 2020, 10, 13727. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Grange, E.; Mannino, G.; van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A biostimulant seed treatment improved heat stress tolerance during cucumber seed germination by acting on the antioxidant system and glyoxylate cycle. Front. Plant Sci. 2020, 11, 836. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-M.; Liu, E.-Q.; Bao, Y.; Duan, S.-L.; She, J.; Liu, H.; Wu, T.-T.; Cao, X.-Q.; Zhang, J.; Li, B.; et al. Low concentration of corn steep liquor promotes seed germination, plant growth, biomass production and flowering in soybean. Plant Growth Regul. 2019, 87, 29–37. [Google Scholar] [CrossRef]
- Strasser, R.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation, 1st ed.; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor & Francis: London, UK, 2000; pp. 443–480. [Google Scholar]
- Marcelis, D.F.M.; Heuvelink, E.; Goudriaan, J. Modelling biomass production and yield of horticultural crops: A review. Sci. Hortic. 1998, 74, 83–111. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Barrameda-Medina, Y.; Montesinos-Pereira, D.; Romero, L.; Blasco, B.; Ruiz, J.M. Role of GSH homeostasis under Zn toxicity in plants with different Zn tolerance. Plant Sci. 2014, 227, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef] [PubMed]
- Bieleski, R.L.; Turner, N.A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Anal. Biochem. 1996, 17, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Navarro-León, E.; Barrameda-Medina, Y.; Lentini, M.; Esposito, S.; Ruiz, J.M.; Blasco, B. Comparative study of Zn deficiency in L. sativa and B. oleracea plants: NH4+ assimilation and nitrogen derived protective compounds. Plant Sci. 2016, 248, 8–16. [Google Scholar] [CrossRef]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plan. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: A study of a modified Berthelot reaction using salicylate and dichloroisocyanurate. Analysis 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plan. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Zhou, K.; Yu, J.; Ma, Y.; Cai, L.; Zheng, L.; Gong, W.; Liu, Q. Corn Steep Liquor: Green Biological Resources for Bioindustry. Appl. Biochem. Biotech. 2022, 194, 3280–3295. [Google Scholar] [CrossRef]
- Fernandez-Salvador, J.; Strick, B.C.; Bryla, D.R. Liquid Corn and Fish Fertilizers Are Good Options for Fertigation in Blackberry Cultivars Grown in an Organic Production System. HortScience 2015, 50, 225–233. [Google Scholar] [CrossRef]
- Kano, K.; Kitazawa, H.; Suzuki, K.; Widiastuti, A.; Odani, H.; Zhou, S.; Chinta, Y.D.; Eguchi, Y.; Shinohara, M.; Sato, T. Effects of Organic Fertilizer on Bok Choy Growth and Quality in Hydroponic Cultures. Agronomy 2021, 11, 491. [Google Scholar] [CrossRef]
- Chinta, Y.D.; Kano, K.; Widiastuti, A.; Fukahori, M.; Kawasaki, S.; Eguchi, Y.; Misu, H.; Odani, H.; Zhou, S.; Narisawa, K.; et al. Effect of corn steep liquor on lettuce root rot (Fusarium oxysporum f. sp. lactucae) in hydroponic cultures. J. Sci. Food Agr. 2014, 94, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, J.W.; Liu, J.; Chen, K.J.; Liu, L.M.; Chen, J.A. Development of chemically defined media supporting high cell density growth of Ketogulonicigenium vulgare and Bacillus megaterium. Bioresour. Technol. 2011, 102, 4807–4814. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Hou, Y.Y.; Liu, Y.; Liu, Y.J.; Zhao, H.Z.; Dong, L.; Du, J.; Wang, Y.; Bai, G.; Luo, G.A. Classification and analysis of corn steep liquor by UPLC/Q-TOF MS and HPLC. Talanta 2013, 107, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Hofer, A.; Herwiga, C. Quantitative determination of nine water-soluble vitamins in the complex matrix of corn steep liquor for rawmaterial quality assessment. J. Chem. Technol. Biot. 2017, 92, 2106–2113. [Google Scholar] [CrossRef]
- Cho, M.-H.; Park, H.L.; Hahn, T.-R. Engineering leaf carbon metabolism to improve plant productivity. Plant Biotechnol. Rep. 2015, 9, 1–10. [Google Scholar] [CrossRef]
- Blázquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of plant hormone response pathways. Annu. Rev. Plant Biol. 2020, 71, 327–353. [Google Scholar] [CrossRef]
- Iqbal, A.; Qiang, D.; Alamzeb, M.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Nianchang, P.; Xiling, Z.; Meizhen, S. Untangling the molecular mechanisms and functions of nitrate to improve nitrogen use efficiency. J. Sci. Food Agr. 2020, 100, 904–914. [Google Scholar] [CrossRef]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef]
- D’Mello, J.P.F. (Ed.) Amino Acids in Higher Plants; CABI International: Edinburgh, UK, 2015. [Google Scholar]
- Mustafa, A.; Hussain, A.; Naveed, M.; Ditta, A.; Nazli, Z.E.H.; Sattar, A. Response of okra (Abelmoschus esculentus L.) to soil and foliar applied L-tryptophan. Soil Environ. 2016, 35, 76–84. [Google Scholar]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Van der Sar, S.; Kim, H.K.; Meissner, A.; Verpoorte, R.; Choi, Y.H. Nuclear Magnetic Resonance spectroscopy for plant metabolite profiling. In The Handbook of Plant Metabolomics, 1st ed.; Weckwerth, W., Kahl, G., Eds.; Wiley-VCH: Weinheim, Germany, 2013; pp. 57–76. [Google Scholar]
- Pervaiz, A.; Iqbal, A.; Khalid, A.; Manzoor, A.; Noreen, S.; Ayaz, A.; Zafar, Z.U.; Athar, H.; Ashraf, M. Proline induced modulation in physiological responses in wheat plants. J. Agric. Environ. Sci. 2019, 8, 112–119. [Google Scholar] [CrossRef]
- Alfosea-Simón, M.; Simón-Grao, S.; Zavala-Gonzalez, E.A.; Cámara-Zapata, J.M.; Simón, I.; Martínez-Nicolás, J.J.; Lidón, V.; Rodríguez-Ortega, W.M.; García-Sánchez, F. Application of biostimulants containing amino acids to tomatoes could favor sustainable cultivation: Implications for tyrosine, lysine, and methionine. Sustainability 2020, 12, 9729. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2005. [Google Scholar]


{kind=link}
{kind=link}
| Treatment | Biomass of the Aerial Part (Dry Weight) (g Plant−¹) | Leaf Area (dm2) | Specific Leaf Area (dm² g−¹) |
|---|---|---|---|
| Control | 4.1 ± 0.7 c | 10.4 ± 0.1 d | 0.23 ± 0.01 a |
| CSL-B Root | 6.7 ± 0.7 a | 12.6 ± 0.3 a | 0.12 ± 0.01 c |
| CSL-B Foliar | 5.1 ± 0.3 b | 11.0 ± 0.1 c | 0.22 ± 0.01 a |
| CSL-H Root | 6.4 ± 0.9 a | 12.1 ± 0.2 b | 0.13 ± 0.01 c |
| CSL-H Foliar | 5.3 ± 0.7 b | 11.4 ± 0.2 c | 0.16 ± 0.01 b |
| p-value | *** | *** | *** |
| Treatment | A (mmol m−2 s−1) | E (mmol m−2 s−1) | gs (mmol m−2 s−1) | WUE (μmol mmol−1) |
|---|---|---|---|---|
| Control | 3.8 ± 0.3 c | 0.28 ± 0.03 ab | 24 ± 3 ab | 13.5 ± 0.3 d |
| CSL-B Root | 6.6 ± 0.9 a | 0.29 ± 0.05 ab | 24 ± 5 ab | 22.6 ± 0.8 a |
| CSL-B Foliar | 5.1 ± 0.4 b | 0.25 ± 0.05 b | 21 ± 6 b | 20.5 ± 1.6 b |
| CSL-H Root | 5.9 ± 0.7 ab | 0.35 ± 0.02 a | 29 ± 2 a | 16.8 ± 1.3 c |
| CSL-H Foliar | 5.1 ± 0.6 b | 0.29 ± 0.03 ab | 32 ± 4 ab | 17.7 ± 0.8 c |
| p-value | ** | * | * | *** |
| Treatment | Fv/Fm | RC/ABS | PIABS | 1–Vj |
|---|---|---|---|---|
| Control | 0.809 ± 0.009 | 0.74 ± 0.05 b | 4.7 ± 0.8 b | 0.70 ± 0.02 |
| CSL-B Root | 0.820 ± 0.005 | 0.85 ± 0.04 a | 8.5 ± 1.3 a | 0.69 ± 0.02 |
| CSL-B Foliar | 0.811 ± 0.003 | 0.77 ± 0.07 ab | 6.0 ± 1.0 b | 0.66 ± 0.02 |
| CSL-H Root | 0.814 ± 0.003 | 0.83 ± 0.06 a | 8.2 ± 1.3 a | 0.69 ± 0.02 |
| CSL-H Foliar | 0.816 ± 0.004 | 0.81 ± 0.05 ab | 7.7 ± 1.1 a | 0.67 ± 0.03 |
| p-value | NS | * | ** | NS |
| Treatment | MDA (µmol g−¹ FW) | Concentration of O2− (µg g−¹ FW) | Concentration of H2O2 (µg g−¹ FW) |
|---|---|---|---|
| Control | 3.2 ± 0.2 | 5.5 ± 0.6 | 124 ± 9 |
| CSL-B Root | 3.4 ± 0.4 | 5.2 ± 0.6 | 117 ± 9 |
| CSL-B Foliar | 3.4 ± 0.6 | 5.4 ± 0.3 | 114 ± 13 |
| CSL-H Root | 3.1 ± 0.1 | 5.9 ± 0.9 | 117 ± 6 |
| CSL-H Foliar | 3.6 ± 0.5 | 5.4 ± 0.2 | 121 ± 15 |
| p-value | NS | NS | NS |
| Control | CSL-B Root | CSL-B Foliar | CSL-H Root | CSL-H Foliar | p-Value | |
|---|---|---|---|---|---|---|
| AIA | 1.5 ± 0.1 c | 2.2 ± 0.4 a | 1.8 ± 0.2 b | 2.0 ± 0.2 a | 1.9 ± 0.3 b | *** |
| tZ | 480 ± 10 d | 820 ± 20 a | 710 ± 20 b | 820 ± 20 a | 680 ± 20 c | *** |
| iP | 0.58 ± 0.03 | 0.62 ± 0.03 | 0.61 ± 0.02 | 0.62 ± 0.05 | 0.61 ± 0.07 | NS |
| GA1 | 0.31 ± 0.03 c | 0.52 ± 0.03 a | 0.40 ± 0.02 b | 0.50 ± 0.02 a | 0.41 ± 0.02 b | *** |
| GA3 | 0.13 ± 0.03 d | 0.21 ± 0.03 bc | 0.18 ± 0.02 c | 0.23 ± 0.03 a | 0.18 ± 0.02 c | *** |
| GA4 | 0.22 ± 0.01 c | 0.46 ± 0.06 a | 0.30 ± 0.02 b | 0.41 ± 0.03 a | 0.32 ± 0.01 b | *** |
| ABA | 57 ± 3 | 60 ± 2 | 62 ± 2 | 60 ± 2 | 59 ± 2 | NS |
| ACC | 14.5 ± 0.2 | 14.7 ± 0.1 | 15.2 ± 0.2 | 14.9 ± 0.6 | 15.3 ± 0.6 | NS |
| JA | 410 ± 6 | 403 ± 6 | 400 ± 4 | 393 ± 4 | 409 ± 6 | NS |
| SA | 3300 ± 300 | 3300 ± 200 | 3300 ± 200 | 3400 ± 300 | 3300 ± 200 | NS |
| Control | CSL-B Root | CSL-H Root | CSL-B Foliar | CSL-H Foliar | p-Value | |
|---|---|---|---|---|---|---|
| Alanine | 20 ± 1 d | 47 ± 2 b | 51 ± 3 b | 31 ± 2 c | 71 ± 4 a | *** |
| Arginine | 3.2 ± 0.2 b | 2.7 ± 0.1 c | 4.7 ± 0.2 a | 1.3 ± 0.1 d | 1.3 ± 0.1 d | *** |
| Asparagine | 4.7 ± 0.2 b | Nd | Nd | 6.3 ± 0.3 a | Nd | *** |
| Aspartate | 53 ± 3 b | 56 ± 3 b | 61 ± 3 a | 31 ± 2 c | 57 ± 3 ab | *** |
| Glutamate | 236 ± 12 b | 275 ± 13 a | 238 ± 12 b | 69 ± 3 d | 186 ± 9 c | *** |
| Glycine | Nd | Nd | 42 ± 2 b | Nd | 58 ± 3 a | *** |
| Histidine | 3.2 ± 0.2 c | 4.2 ± 0.2 b | 3.5 ± 0.2 c | 2.0 ± 0.1 d | 5.9 ± 0.2 a | *** |
| Isoleucine | 560 ± 30 d | 630 ± 30 c | 810 ± 40 a | 720 ± 40 b | 670 ± 30 bc | *** |
| Leucine | 820 ± 40 d | 920 ± 50 c | 1180 ± 60 a | 1060 ± 50 b | 970 ± 50 bc | *** |
| Lysine | 1.41 ± 0.07 b | 0.84 ± 0.04 d | 0.59 ± 0.03 e | 1.03 ± 0.05 c | 2.0 ± 0.1 a | *** |
| Phenylalanine | 179 ± 9 b | 187 ± 9 b | 220 ± 10 a | 220 ± 10 a | 220 ± 10 a | *** |
| Proline | 80 ± 4 d | 210 ± 10 a | 210 ± 10 a | 190 ± 10 b | 170 ± 9 c | *** |
| Serine | 14.8 ± 0.7 c | 23 ± 1 a | 17.4 ± 0.9 b | 12.4 ± 0.6 d | 24 ± 1 a | *** |
| Threonine | 7.4 ± 0.4 c | 11.9 ± 0.6 a | 10.0 ± 0.5 b | 4.7 ± 0.2 d | 10.1 ± 0.5 b | *** |
| Tryptophan | 244 ± 12 c | 336 ± 17 b | 327 ± 16 b | 510 ± 30 a | 520 ± 30 a | *** |
| Tyrosine | 331 ± 17 a | 148 ± 7 c | 176 ± 9 b | 134 ± 7 c | 184 ± 9 b | *** |
| Valine | 111 ± 6 e | 155 ± 5 c | 200 ± 10 a | 136 ± 7 d | 176 ± 9 b | *** |
| Methionine | 25 ± 1 a | 18 ± 1 b | 12 ± 1 c | 9 ± 1 d | 18 ± 1 b | *** |
| Cysteine | 1.16 ± 0.06 b | 1.03 ± 0.05 c | 1.53 ± 0.08 a | Nd | Nd | *** |
| 4–OH Proline | 6.1 ± 0.3 c | 9.7 ± 0.5 a | 8.7 ± 0.4 b | 4.7 ± 0.2 d | 8.4 ± 0.4 b | *** |
| Total | 270 ± 100 d | 3000 ± 200 c | 3600 ± 200 a | 3200 ± 200 bc | 3400 ± 200 ab | *** |
| Treatment | P | K | Ca | Mg | S | N | NO3− |
|---|---|---|---|---|---|---|---|
| Control | 4.6 ± 0.2 d | 41 ± 2 c | 17.5 ± 0.7 c | 4.5 ± 0.2 c | 4.01 ± 0.2 e | 65 ± 2 b | 14.2 ± 0.3 b |
| CSL-B Root | 5.3 ± 0.2 bc | 47 ± 2 b | 19.2 ± 0.8 b | 6.7 ± 0.3 a | 5.2 ± 0.2 a | 65 ± 2 c | 12.8 ± 0.6 c |
| CSL-H Root | 5.1 ± 0.2 c | 50 ± 2 a | 19.3 ± 0.8 b | 5.7 ± 0.2 b | 4.5 ± 0.2 b | 63 ± 2 a | 16 ± 1 a |
| CSL-B Foliar | 5.6 ± 0.2 b | 46 ± 2 b | 21.3 ± 0.9 a | 6.3 ± 0.3 a | 4.2 ± 0.2 d | 63 ± 2 c | 12.9 ± 0.7 c |
| CSL-H Foliar | 6.0 ± 0.2 a | 51 ± 2 a | 21.2 ± 0.9 a | 6.7 ± 0.3 a | 4.8 ± 0.2 c | 64 ± 2 b | 14.9 ± 0.3 b |
| p-value | *** | *** | *** | *** | *** | *** | *** |
| Fe | Cu | Mn | Zn | Mo | B | ||
| Control | 88 ± 4 d | 59 ± 2 | 106 ± 4 | 55 ± 2 c | 0.58 ± 0.02 c | 12.4 ± 0.4 | |
| CSL-B Root | 106 ± 4 a | 55 ± 1 | 108 ± 4 | 73 ± 3 a | 1.12 ± 0.04 a | 11.1 ± 0.5 | |
| CSL-H Root | 95 ± 4 bc | 59 ± 2 | 103 ± 4 | 64 ± 3 b | 0.91 ± 0.04 b | 14.0 ± 0.6 | |
| CSL-B Foliar | 90 ± 4 cd | 58 ± 2 | 112 ± 4 | 63 ± 3 b | 0.89 ± 0.04 b | 12.6 ± 0.5 | |
| CSL-H Foliar | 100 ± 4 ab | 57 ± 2 | 111 ± 4 | 73 ± 3 a | 0.66 ± 0.03 c | 13.1 ± 0.5 | |
| p-value | ** | NS | NS | *** | *** | NS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Morillo, I.; Navarro-Perez, V.; Perez-Millan, R.; Navarro-León, E.; Blasco, B.; Cámara-Zapata, J.M.; Garcia-Sanchez, F. Effects of Root and Foliar Application of Corn Steep Liquor on Pepper Plants: A Physiological, Nutritional, and Morphological Study. Horticulturae 2023, 9, 221. https://doi.org/10.3390/horticulturae9020221
Navarro-Morillo I, Navarro-Perez V, Perez-Millan R, Navarro-León E, Blasco B, Cámara-Zapata JM, Garcia-Sanchez F. Effects of Root and Foliar Application of Corn Steep Liquor on Pepper Plants: A Physiological, Nutritional, and Morphological Study. Horticulturae. 2023; 9(2):221. https://doi.org/10.3390/horticulturae9020221
Chicago/Turabian StyleNavarro-Morillo, Iván, Valeria Navarro-Perez, Rafael Perez-Millan, Eloy Navarro-León, Begoña Blasco, Jose Maria Cámara-Zapata, and Francisco Garcia-Sanchez. 2023. "Effects of Root and Foliar Application of Corn Steep Liquor on Pepper Plants: A Physiological, Nutritional, and Morphological Study" Horticulturae 9, no. 2: 221. https://doi.org/10.3390/horticulturae9020221
APA StyleNavarro-Morillo, I., Navarro-Perez, V., Perez-Millan, R., Navarro-León, E., Blasco, B., Cámara-Zapata, J. M., & Garcia-Sanchez, F. (2023). Effects of Root and Foliar Application of Corn Steep Liquor on Pepper Plants: A Physiological, Nutritional, and Morphological Study. Horticulturae, 9(2), 221. https://doi.org/10.3390/horticulturae9020221