
Physiological Response of Four Widely Cultivated Sunflower Cultivars to Cadmium Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Plant Culture
2.3. Plant Growth Parameters
2.4. Gas Exchange Measurement
2.5. Determination of Cd Concentrations
2.6. NBT, DAB Analysis, Starch, Total Soluble Sugar, and Conductivity Determination
2.7. Statistical Analysis
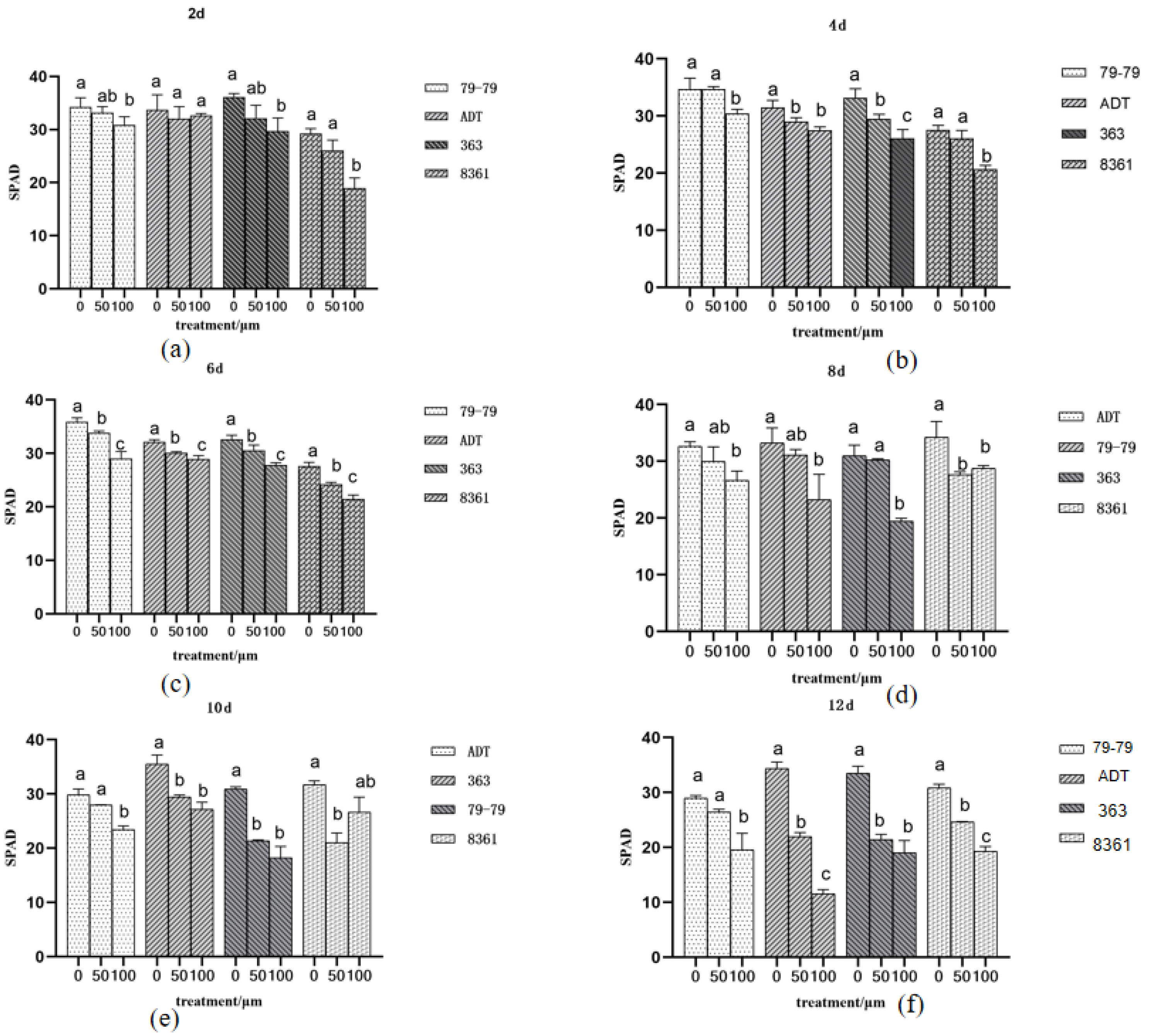
3. Results
3.1. Phenotype Changes under Different Cd Treatments
3.2. Effects on the Photosynthesis of Sunflowers
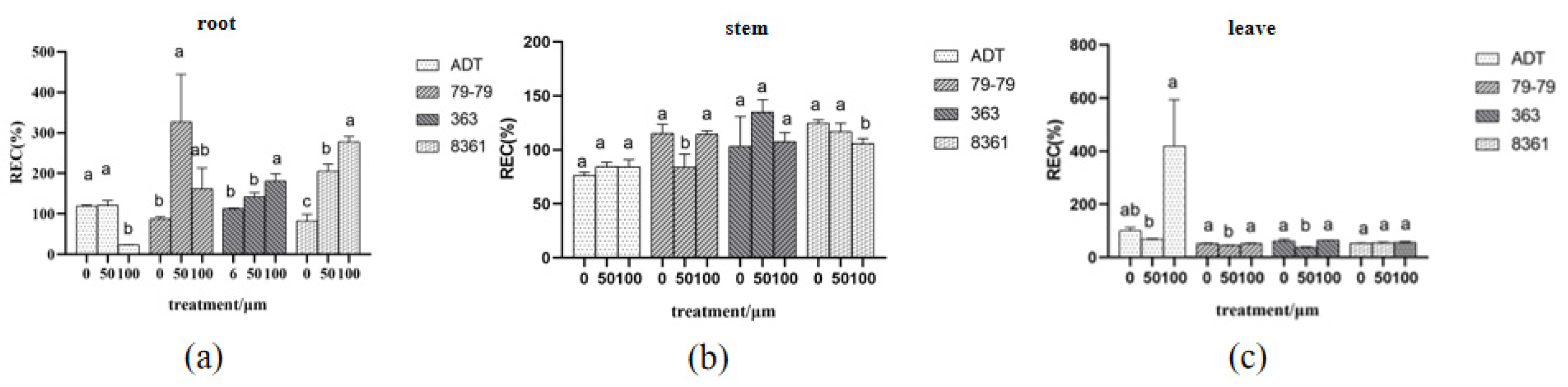
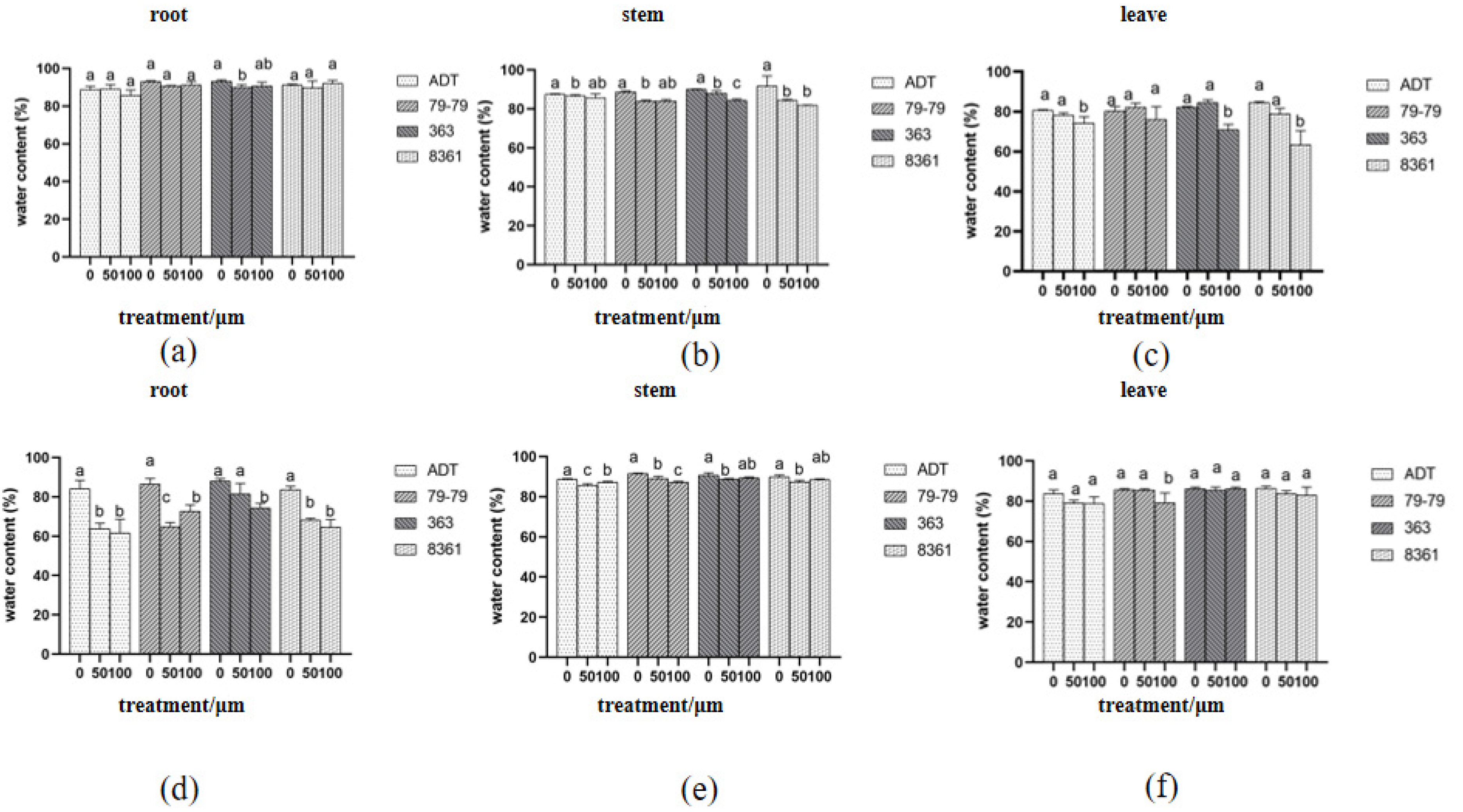
3.3. Relative Electrical Conductivity and Water Content Changed under Different Cd Concentration
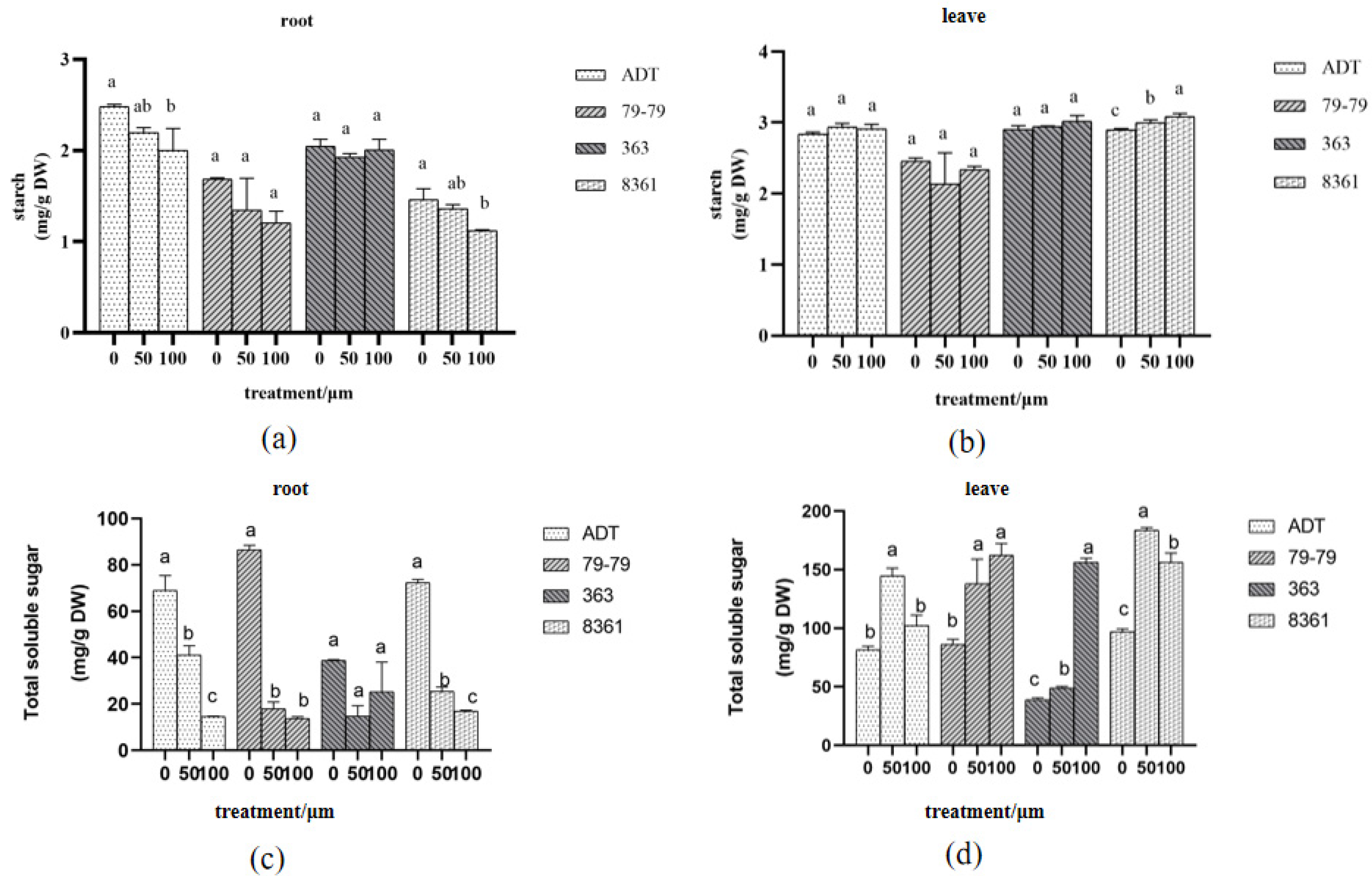
3.4. Changes of Starch and Total Soluble Sugar Content
3.5. Histochemical Detection of H2O2 and O2−
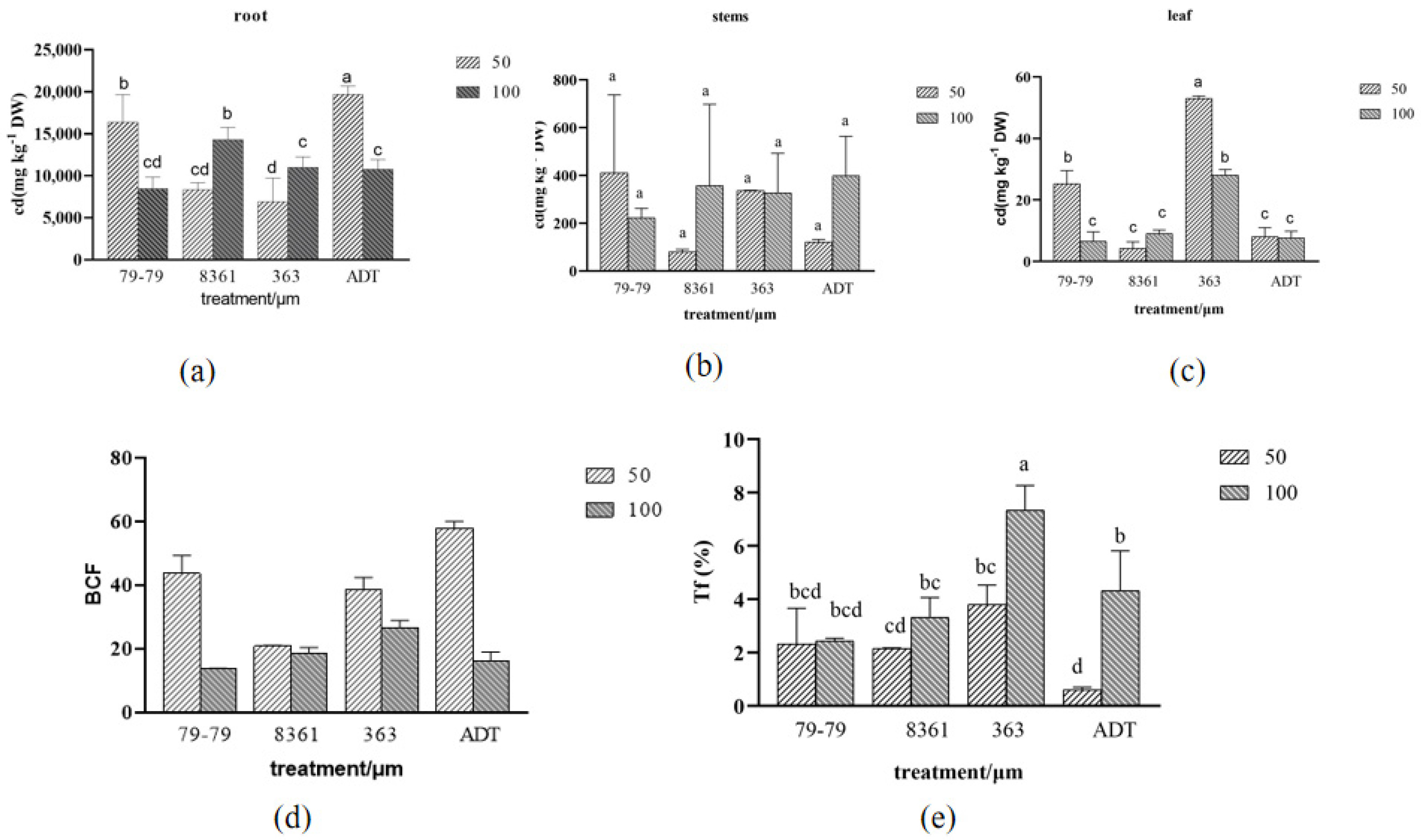
3.6. Cadmium Content in Sunflowers
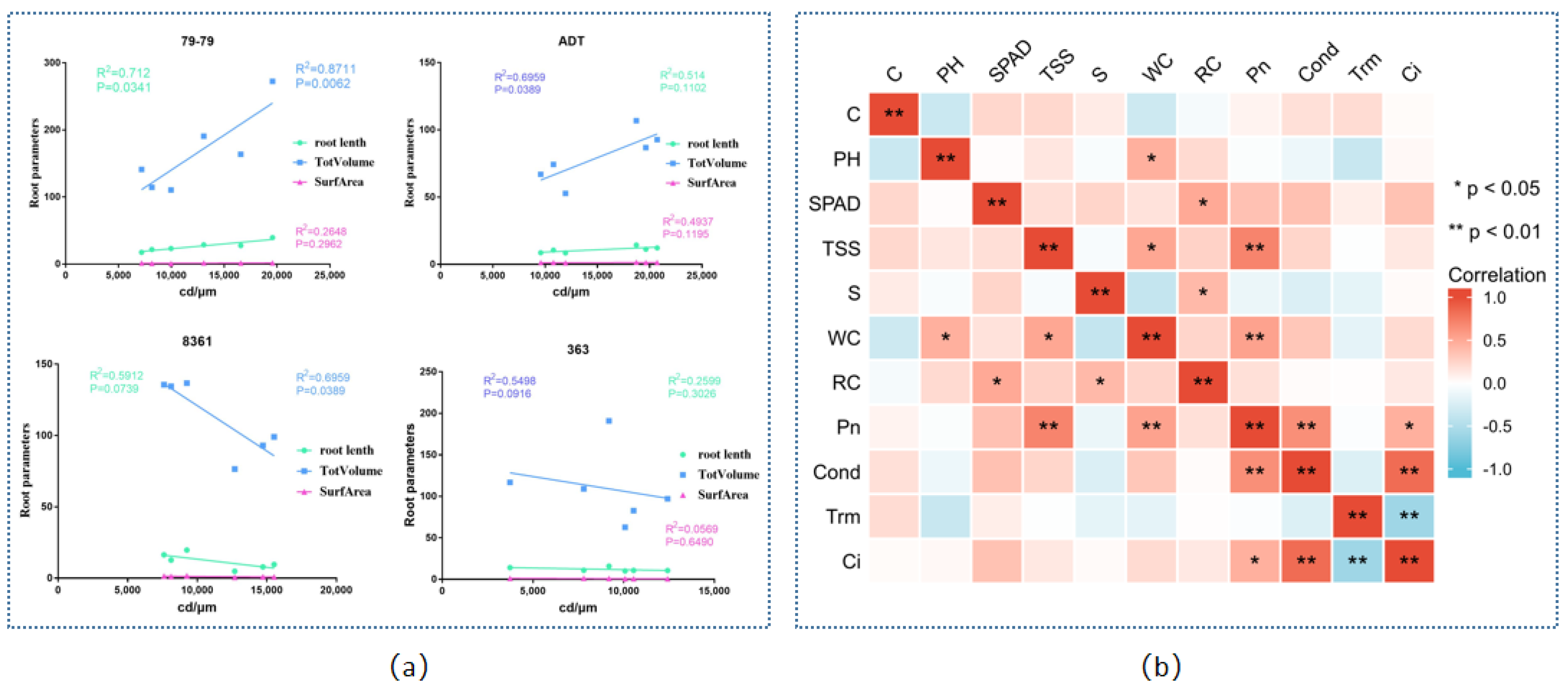
3.7. Correlation Analysis
4. Discussion
4.1. Cd Effect on Growth
4.2. Cd Tolerance Is Associated with Physiological Changes
4.3. Accumulation and Distribution of Cd in Sunflowers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benavides, M.P.; Groppa, M.D.; Recalde, L.; Verstraeten, S.V. Effects of polyamines on cadmium−and copper−mediated alterations in wheat (Triticum aestivum. L.) and sunflower (Helianthus annuus. L.) seedling membrane fluidity. Arch. Biochem. Biophys. 2018, 654, 27–39. [Google Scholar] [CrossRef]
- Luo, Q.; Bai, B.; Xie, Y.; Yao, D.; Zhang, D.; Chen, Z.; Zhuang, W.; Deng, Q.; Xiao, Y.; Wu, J. Effects of Cd uptake, translocation and redistribution in different hybrid rice varieties on grain Cd concentration. Ecotoxicol. Environ. Saf. 2022, 240, 113683. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Ma, H.; Li, L.L.; Gao, Y.F.; Li, Y.Z.; Xu, H. Subcellular distribution, chemical forms and physiological responses involved in cadmium tolerance and detoxification in Agrocybe Aegerita. Ecotoxicol. Environ. Saf. 2019, 171, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Li, L.; Duan, Q.; Liu, X.; Chen, M. Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal. Behav. 2021, 16, 1836884. [Google Scholar] [CrossRef]
- Cornu, J.Y.; Bussière, S.; Coriou, C.; Robert, T.; Maucourt, M.; Deborde, C.; Moing, A.; Nguyen, C. Changes in plant growth, Cd partitioning and xylem sap composition in two sunflower cultivars exposed to low Cd concentrations in hydroponics. Ecotoxicol. Environ. Saf. 2020, 205, 111145. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Ma, C.F.; Ma, Y.L.; Li, H.; Kang, J.Q.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Cadmium tolerance in six poplar species. Environ. Sci. Pol. 2013, 20, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Nan, Z.R.; Zhao, Z.J. Bioaccumulation and translocation of cadmium in wheat (Triticumaestivum L.) and maize (Zea mays L.) from the polluted oasis soil of Northwestern China. Chem. Speciat. Bioavailab. 2014, 26, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.H.; Liu, C.Y.; Zhao, X.Y.; Li, R.Y.; Deng, W.J. Involvement of an antioxidant defense system in the adaptive response to cadmium in maize seedlings (Zea mays L.). Bull. Environ. Contam. Toxicol. 2014, 93, 618–624. [Google Scholar] [CrossRef]
- Huang, L.; Li, W.C.; Tam, N.F.C.; Ye, Z.H. Effects of root morphology and anatomy on cadmium uptake and translocation in rice (Oryza sativa L.). J. Environ. Sci. 2019, 75, 296–306. [Google Scholar] [CrossRef]
- Ashraf, U.; Mahmood, M.H.; Hussain, S.; Abbas, F.; Anjum, S.A.; Tang, X. Lead (Pb) distribution and accumulation in different plant parts and its associations with grain Pb contents in fragrant rice. Chemosphere 2020, 248, 126003. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, P.; Zhang, S.; Dai, J.; Chen, H.P.; Lombi, E.; Howard, D.L.; van der Ent, A.; Zhao, F.J.; Kopittke, P.M. Chemical speciation and distribution of cdmium in rice grain and implications for bioavailability to humans. Environ. Sci. Technol. 2020, 54, 12072–12080. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Hamid, Y.; Zehra, A.; Sahito, Z.A.; He, Z.; Hussain, B.; Yang, X. Characterization of fava bean (Viciafaba L.) genotypes for phytoremediation of cadmium and lead co−contaminated soils coupled with agro−production. Ecotoxicol. Environ. Saf. 2019, 171, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lai, J.L.; Ji, X.H.; Luo, X.G. Unraveling response mechanism of photosynthetic metabolism and respiratory metabolism to uranium−exposure in Vicia faba. J. Hazard. Mater. 2020, 398, 122997. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Tian, S.K.; Yang, X.E.; Wang, X.C.; Brown, P.; Li, T.Q.; He, Z.L. Enhanced root−to−shoot translocation of cadmium in the hyperaccumulating ecotype of Sedum alfredii. J. Exp. Bot. 2008, 59, 3203–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.K.; Lu, L.L.; Yang, X.E.; Webb, S.M.; Du, Y.H.; Brown, P.H. Spatial imaging and speciation of lead in the accumulator plant Sedum alfredii by microscopically focused synchrotron X-ray investigation. Environ. Sci. Technol. 2010, 44, 5920–5926. [Google Scholar] [CrossRef]
- Sarwar, N.; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.C.; Zhang, Y.J.; He, X.F.; Li, X.F.; Zhang, X.X. Analysis of the report on the national general survey of soil contamination. J. Agro−Environ. Sci. 2017, 36, 1689–1692. [Google Scholar]
- Zehra, A.; Sahito, Z.A.; Tong, W.; Tang, L.; Hamid, Y.; Wang, Q.; Cao, X.; Khan, M.B.; Hussain, B.; Jatoi, S.A.; et al. Identification of high cadmium−accumulating oilseed sunflower (Helianthus annuus) cultivars for phytoremediation of an Oxisol and an Inceptisol. Ecotoxicol. Environ. Saf. 2020, 187, 109857. [Google Scholar] [CrossRef]
- Xu, L.; Li, J.J.; Najeeb, U.; Li, X.; Pan, J.M.; Huang, Q.; Zhou, W.J.; Liang, Z.S. Synergistic effects of EDDS and ALA on phytoextraction of cadmium as revealed by biochemical and ultrastructural changes in sunflower (Helianthus annuus L.) tissues. J. Hazard. Mater. 2021, 407, 124764. [Google Scholar] [CrossRef]
- Marques, A.P.; Moreira, H.; Franco, A.R.; Rangel, A.O.; Castro, P.M. Inoculating sunflower (Helianthus annuus) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria−effects on phytoremediation strategies. Chemosphere 2013, 92, 74–83. [Google Scholar] [CrossRef]
- Wang, A.; Wang, M.; Liao, Q.; He, X. Characterization of Cd translocation and accumulation in 19 maize cultivars grown on Cd−contaminated soil: Implication of maize cultivar selection for minimal risk to human health and for phytoremediation. Environ. Sci. Pollut. Res. 2016, 23, 5410–5419. [Google Scholar] [CrossRef]
- Nehnevajova, E.; Herzig, R.; Federer, G.; Erismann, K.H.; Schwitzguébel, J.P. Screening of sunflower cultivars for metal phytoextraction in a contaminated field prior to mutagenesis. Int. J. Phytoremediation 2005, 7, 337–349. [Google Scholar] [CrossRef]
- Adesodun, J.K.; Atayese, M.O.; Agbaje, T.A.; Osadiaye, B.A.; Mafe, O.F.; Soretire, A.A. Phytoremediation potentials of sunflowers (Tithonia diversifolia and Helianthus annuus) for metals in soils contaminated with zinc and lead nitrates. Water Air Soil Pollut. 2010, 207, 195–201. [Google Scholar] [CrossRef]
- Seth, C.S.; Misra, V.; Singh, R.R.; Zolla, L. EDTA−enhanced lead phytoremediation in sunflflower (Helianthus annuus L.) hydroponic culture. Plant Soil 2011, 347, 231. [Google Scholar] [CrossRef]
- Doncheva, S.; Moustakas, M.; Ananieva, K.; Chavdarova, M.; Gesheva, E.; Vassilevska, R.; Mateev, P. Plant response to lead in the presence or absence EDTA in two sunflower genotypes (cultivated H. annuus cv. 1114 and interspecific line H. annuus×H. argophyllus). Environ. Sci. Pol. 2013, 20, 823–833. [Google Scholar] [CrossRef]
- Motesharezadeh, B.; Navabzadeh, M.; Liyaghat, A.M. Modeling phytoremediation of cadmium contaminated soil with sunflower (Helianthus annus) under salinity stress. Int. J. Environ. Res. 2016, 10, 109–118. [Google Scholar]
- Hu, Z.Y.; Zhang, Y.F.; He, Y.; Cao, Q.Q.; Zhang, T.; Lou, L.Q.; Cai, Q.S. Full length transcriptome assembly of italian ryegrass root integrated with RNA−seq to identify genes in response to plant cadmium stress. Int. J. Mol. Sci. 2020, 21, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.C.; Monteiro, C.; Moutinho−Pereira, J.; Correia, C.; Goncalves, B.; Santos, C. Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol. Plant. 2013, 35, 1281–1289. [Google Scholar] [CrossRef]
- Rasafi, T.E.; Oukarroum, A.; Haddioui, A.; Song, H.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2020, 5, 675–726. [Google Scholar] [CrossRef]
- Guo, Z.H.; Zeng, P.; Xiao, X.Y.; Peng, C. Physiological, anatomical, and transcriptional responses of mulberry (Morus alba L.) to Cd stress in contaminated soil. Environ. Pollut. 2021, 284, 117387. [Google Scholar] [CrossRef]
- Manaa, A.; Goussi, R.; Derbali, W.; Cantamessa, S.; Essemine, J.; Barbato, R. Photosynthetic performance of quinoa (Chenopodium quinoa.) after exposure to a gradual drought stress followed by a recovery period. Bba−Bioenerg. 2021, 1862, 148383. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, F.; Gao, S.; Wang, X. Heavy metal accumulation in different rice cultivars as influenced by foliar application of nano−silicon. Water Air Soil Pollut. 2016, 227, 1–13. [Google Scholar] [CrossRef]
- Huang, S.; Rao, G.; Ashraf, U.; Deng, Q.; Dong, H.; Zhang, H. Ultrasonic seed treatment improved morpho−physiological and yield traits and reduced grain Cd concentrations in rice. Ecotoxicol. Environ. Saf. 2021, 214, 112119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Lei, X.J.; Wang, P.L.; Wang, Y.Y.; Lv, J.X.; Li, X.P.; Gao, C.Q. Overexpression of ThSAP30BP from Tamarix hispida improves salt tolerance. Plant Physiol. Bioch. 2020, 146, 124–132. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Ismayilov, A.I.; Mamedov, A.I.; Fujimaki, H.; Tsunekawa, A.; Levy, G.J. Soil salinity type effects on the relationship between the electrical conductivity and salt content for 1:5 soil−to−water extract. Sustainability 2021, 13, 3395. [Google Scholar] [CrossRef]
- Yan, B.F.; Nguyen, C.; Pokrovsky, O.S.; Candaudap, F.; Coriou, C.; Bussiere, S.; Robert, T.; Cornu, J.Y. Cadmium allocation to grains in durum wheat exposed to low Cd concentrations in hydroponics. Ecotoxicol. Environ. Saf. 2019, 184, 109592. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Yang, C.Y.; Zhang, L.B.; Li, L.Z.; Liu, S.J.; Yu, J.B.; You, L.P.; Zhou, D.; Xia, C.H.; Zhao, J.M.; et al. Metabolic profiling of cadmium−induced effects in one pioneer intertidal halophyte Suaeda salsa by NMR−based metabolomics. Ecotoxicology 2011, 20, 1422–1431. [Google Scholar] [CrossRef]
- Zhao, D.; Li, T.; Shen, M.; Wang, J.L.; Zhao, Z.W. Diverse strategies conferring extreme cadmium (Cd) tolerance in the dark septate endophyte (DSE), Exophiala pisciphila: Evidence from RNA−seq data. Microbiol. Res. 2015, 170, 27–35. [Google Scholar] [CrossRef]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef]
- Peco, J.D.; Campos, J.A.; Romero−Puertas, M.C.; Olmedilla, A.; Higueras, P.; Sandalio, L.M. Characterization of mechanisms involved in tolerance and accumulation of Cd in Biscutella auriculata L. Ecotoxicol. Environ. Saf. 2020, 201, 110784. [Google Scholar] [CrossRef] [PubMed]
- Shahabivand, S.; Maivan, H.Z.; Goltapeh, E.M.; Sharifi, M.; Aliloo, A.A. The effects of root endophyte and arbuscular mycorrhizal fungi on growth and cadmium accumulation in wheat under cadmium toxicity. Plant Physiol. Bioch. 2012, 60, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Mangal, M.; Agarwal, M.; Bhargava, D. Effect of cadmium and zinc on growth and biochemical parameters of selected vegetables. J. Pharmacogn. Phytochem. 2013, 2, 106–114. [Google Scholar]
- Ren, L.; Wang, M.R.; Wang, Q.C. ROS−induced oxidative stress in plant cryopreservation: Occurrence and alleviation. Planta 2021, 254, 124. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. 2−Hydroxymelatonin Promotes Seed Germination by Increasing Reactive Oxygen Species Production and Gibberellin Synthesis in Arabidopsis thaliana. Antioxidants 2022, 11, 737. [Google Scholar] [CrossRef]
- Li, W.Y.; Chen, B.X.; Chen, Z.J.; Gao, Y.T.; Chen, Z.; Liu, J. Reactive Oxygen Species Generated by NADPH Oxidases Promote Radicle Protrusion and Root Elongation during Rice Seed Germination. Int. J. Mol. Sci. 2017, 18, 110. [Google Scholar] [CrossRef]
- Kacalkova, L.; Tlustos, P.; Szakova, J. Phytoextraction of risk elements by willow and poplar trees. Int. J. Phytoremediation 2015, 17, 414–421. [Google Scholar] [CrossRef]
- Balabanova, B.; Stafifilov, T.; Liping, F.; Meicong, W. Bioavailability and bioaccumulation characterization of essential and heavy metals contents in various plant food from polluted and referent areas. J. Environ. Health Sci. Eng. 2016, 13, 2. [Google Scholar]
- Tang, X.; Pang, Y.; Ji, P.; Gao, P.; Nguyen, T.H.; Tong, Y. Cadmium uptake in above−ground parts of lettuce (Lactuca sativa L.). Ecotoxicol. Environ. Saf. 2016, 125, 102–106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Treatment | TotVolume (cm3) | Length (mm) | ProjArea (cm2) | SurfArea (cm2) | Volume (cm3) |
|---|---|---|---|---|---|---|
| 79−79 | CK | 227.02 ± 31.68 a | 32.03 ± 1.94 a | 110.61 ± 11.85 a | 2.13 ± 0.71 a | 5.76 ± 1.83 a |
| 50 | 208.96 ± 56.67 a | 31.91 ± 6.58 a | 90.25 ± 3.75 b | 1.62 ± 0.05 ab | 3.54 ± 0.49 ab | |
| 100 | 122.03 ± 16.67 b | 20.82 ± 2.97 b | 65.41 ± 9.33 c | 1.16 ± 0.39 b | 1.97 ± 0.52 b | |
| ADT | CK | 137.68 ± 10.73 a | 33.12 ± 5.98 a | 104.05 ± 18.79 a | 2.55 ± 0.39 a | 6.70 ± 2.06 a |
| 50 | 95.47 ± 10.28 b | 12.62 ± 1.61 b | 39.65 ± 5.08 b | 1.45 ± 0.14 b | 1.24 ± 0.05 b | |
| 100 | 64.69 ± 10.96 c | 9.26 ± 1.14 b | 29.09 ± 3.6 b | 1.24 ± 0.09 b | 1.04 ± 0.13 b | |
| 363 | CK | 203.41 ± 7.7 a | 26.36 ± 3.44 a | 82.81 ± 10.81 a | 2.14 ± 0.69 a | 4.31 ± 0.89 a |
| 50 | 138.93 ± 45.19 b | 13.62 ± 2.51 b | 42.79 ± 7.91 b | 1.12 ± 0.16 b | 0.91 ± 0.07 b | |
| 100 | 80.82 ± 17.27 c | 10.48 ± 1.48 b | 30.83 ± 4.65 b | 0.69 ± 0.11 b | 0.70 ± 0.18 b | |
| 8361 | CK | 162.02 ± 17.16 a | 24.66 ± 2.86 a | 77.46 ± 8.98 a | 3.01 ± 1.45 a | 4.32 ± 0.3 a |
| 50 | 135.61 ± 1.13 b | 16.26 ± 3.53 b | 51.08 ± 11.09 b | 1.45 ± 0.2 ab | 2.19 ± 0.06 b | |
| 100 | 89.52 ± 11.7 c | 7.46 ± 2.41 c | 24.43 ± 7.58 c | 0.86 ± 0.21 b | 0.94 ± 0.07 c |
| Varieties | Treatment | Pn | Cond | Ci | Trm |
|---|---|---|---|---|---|
| 79−79 | CK | 10.45 ± 0.033 a | 0.65 ± 0.03 a | 151.7 ± 2.65 a | 13.3 ± 0.06 b |
| 50 | 8.14 ± 0.06 b | 0.46 ± 0.005 b | 143.34 ± 0.19 b | 14.3 ± 0.15 a | |
| 100 | 6.17 ± 0.19 c | 0.36 ± 0.01 c | 143.5 ± 0.5 c | 12.7 ± 0.31 c | |
| ADT | CK | 9.53± 0.11 a | 0.43 ± 0.01 b | 132.5 ± 1.6 b | 13.9 ± 0.21 a |
| 50 | 7.88 ± 0.53 b | 0.79 ± 0.03 a | 155.5 ± 2.5 a | 13.1 ± 0.33 b | |
| 100 | 4.43 ± 0.2 c | 0.18 ± 0.01 c | 149.6 ± 4.9 a | 4.73 ± 0.01 c | |
| 363 | CK | 8.64 ± 0.087 a | 0.32 ± 0.02 a | 141.5 ± 1.42 a | 9.08 ± 0.13 c |
| 50 | 6.35 ± 0.219 b | 0.33 ± 0.011 a | 135.5 ± 1.31 b | 9.69 ± 0.24 b | |
| 100 | 4.28 ± 0.105 c | 0.43 ± 0.015 b | 128.6 ± 0.049 c | 4.45 ± 0.21 c | |
| 8361 | CK | 9.39 ± 0.021 a | 0.43 ± 0.01 a | 139.9 ± 0.65 c | 14 ± 0.07 a |
| 50 | 7.25 ± 0.077 b | 0.3 ± 0.01 b | 135.3 ± 1.33 b | 7.1 ± 0.1 b | |
| 100 | 4.25 ± 0.123 c | 0.22 ± 0.005 c | 143.5 ± 0.58 a | 4.45 ± 0.21 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, D.; Zhang, L.; Zhang, S.; Cui, B. Physiological Response of Four Widely Cultivated Sunflower Cultivars to Cadmium Stress. Horticulturae 2023, 9, 320. https://doi.org/10.3390/horticulturae9030320
Tan D, Zhang L, Zhang S, Cui B. Physiological Response of Four Widely Cultivated Sunflower Cultivars to Cadmium Stress. Horticulturae. 2023; 9(3):320. https://doi.org/10.3390/horticulturae9030320
Chicago/Turabian StyleTan, Dingquan, Lingling Zhang, Sheng Zhang, and Bei Cui. 2023. "Physiological Response of Four Widely Cultivated Sunflower Cultivars to Cadmium Stress" Horticulturae 9, no. 3: 320. https://doi.org/10.3390/horticulturae9030320