
Identification of Causal Gene-Specific SNP Markers for the Development of Gynoecious Hybrids in Cucumber (Cucumis sativa L.) Using the PathoLogic Algorithm

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Genotyping-by-Sequencing
2.3. Reference-Based SNP Calling and Identification of SNPs
2.4. Sex-Specific SNP Discovery/Chromosome Strolling
2.5. Statistical Analysis
2.6. Data Deposition
3. Results
3.1. Field Evaluation of Parents and Hybrids
3.2. Statistics of Genotyping-by-Sequencing (GBS) Data
3.3. SNP Detection and Annotation
3.4. InDel Detection and Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berry, P.E. Cucurbitales. In Encyclopedia Britannica; Encyclopsedia Britannica: Chicago, IL, USA, 2019; Available online: https://www.britannica.com/plant/Cucurbitales (accessed on 19 January 2023).
- Sebastian, P.; Schaefer, H.; Telford, I.R.H.; Renner, S.S. Cucumber (Cucumis sativus L.) and melon (C. melo) have numerous wild relatives in Asia and Australia, and the sister species of melon is from Australia. Proc. Natl. Acad. Sci. USA 2010, 10, 14269–14273. [Google Scholar] [CrossRef] [Green Version]
- Meglic, V.; Serquen, F.; Staub, J.E. Genetic diversity in cucumber (Cucumis sativus L.): I. A re-evaluation of the U.S. germplasm collection. Genet. Resour. Crop Evol. 1996, 43, 533–546. [Google Scholar] [CrossRef]
- Staub, J.E.; Serquen, F.C.; Horejsi, T.; Chen, J. Genetic diversity in cucumber (Cucumis sativus L.): IV. An evaluation of Chinese germplasm. Genet. Resour. Crop Evol. 1999, 46, 297–310. [Google Scholar] [CrossRef]
- Staub, J.E.; Robbins, M.D.; Wehner, T.C. Cucumber. In Vegetables I. Handbook of Plant Breeding; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; Volume 1, pp. 241–282. [Google Scholar] [CrossRef]
- Naegele, R.P.; Wehner, T.C. Genetic resources of cucumber. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 61–86. [Google Scholar] [CrossRef]
- Rolnik, A.; Olas, B. Vegetables from the Cucurbitaceae family and their products: Positive effect on human health. Food Sci. Nutr. 2020, 78, 110788. [Google Scholar] [CrossRef]
- Li, H.; Wang, S.; Chai, S.; Yang, Z.; Zhang, Q.; Xin, H.; Xu, Y.; Lin, X.; Chen, X.; Yao, Z.; et al. Graph-based pangenome reveals structural and sequence variations related to agronomic traits and domestication in cucumber. Nat. Commun. 2022, 13, 682. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Sheng, Y.; Niu, H.; Li, Z. Gene interactions regulating sex determination in Cucurbits. Front. Plant Sci. 2019, 10, 1231. [Google Scholar] [CrossRef] [Green Version]
- Galun, E. Study of the inheritance of sex expression in the cucumber. The interaction of major genes with modifying genetic and non-genetic factors. Genetica 1961, 32, 134–163. [Google Scholar] [CrossRef]
- Kubicki, B. Investigations on sex determination in cucumbers (Cucumis sativus L.). V. Genes controlling intensity of femaleness. Genet. Pol. 1969, 10, 69–86. [Google Scholar]
- Kubicki, B. Investigations on sex determination in cucumber (Cucumis sativus L.). VII. Andromonoecism and hermaphroditism. Genet. Pol. 1969, 10, 101–120. [Google Scholar]
- Kubicki, B. Investigations on sex determination in cucumbers (Cucumis sativus L.). VI. Androecism. Genet. Pol. 1969, 10, 87–99. [Google Scholar]
- Coherent Market Insights. Available online: https://www.coherentmarketinsights.com/ (accessed on 30 June 2022).
- Jat, G.S.; Munshi, A.D.; Behera, T.K.; Choudhary, H.; Dash, P.; Ravindran, A.; Kumari, S. Genetics and molecular mapping of gynoecious (F) locus in cucumber (Cucumis sativus L.). J. Hort. Sci. Biot. 2019, 94, 24–32. [Google Scholar] [CrossRef]
- Boopathi, N.M. Genetic Mapping and Marker Assisted Selection: Basics, Practice and Benefits; Springer: Singapore, 2021. [Google Scholar]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, G.; Sim, S.C.; Jung, J.K.; Shim, E.J.; Chung, S.M.; Lee, G.P.; et al. Development of genome-wide single nucleotide polymorphism markers for variety identification of F1 hybrids in cucumber (Cucumis sativus L.). Sci. Hortic. 2021, 285, 110173. [Google Scholar] [CrossRef]
- Qi, J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, M.J.; Díaz, A.; Dhillon, N.P.S.; Reddy, U.K.; Picó, B.; Monforte, A.J. Re-evaluation of the role of Indian germplasm as center of melon diversification based on genotyping-by-sequencing analysis. BMC Genom. 2019, 20, 448. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Marcotrigiano, A.R.; Ciani, E.; Mazzeo, R.; Zonno, V.; Ruggieri, V.; Lotti, C.; Ricciardi, L. Genotyping-by-sequencing of a melon (Cucumis melo L.) germplasm collection from a secondary center of diversity highlights patterns of genetic variation and genomic features of different gene pools. BMC Genom. 2017, 18, 59. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Miller, A.N.; McGrath, M.; Babadoost, M. Genotyping-by-sequencing for analysis of the genetic variation of Podosphaera xanthii, incitant of cucurbit powdery mildew. Plant Dis. 2020, 104, 951–957. [Google Scholar] [CrossRef]
- Wang, C.; Xin, M.; Zhou, X.; Liu, W.; Liu, D.; Qin, Z. Transcriptome profiling reveals candidate genes associated with sex differentiation induced by night temperature in cucumber. Sci. Hortic. 2018, 232, 162–169. [Google Scholar] [CrossRef]
- Wang, Y.; VandenLangenberg, K.; Wen, C.; Wehner, T.C.; Weng, Y. QTL mapping of downy and powdery mildew resistances in PI 197088 cucumber with genotyping-by-sequencing in RIL population. Theor. Appl. Genet. 2018, 131, 597–611. [Google Scholar] [CrossRef]
- Pujol, M.; Alexiou, K.G.; Fontaine, A.-S.; Mayor, P.; Miras, M.; Jahrmann, T.; Garcia-Mas, J.; Aranda, M.A. Mapping cucumber vein yellowing virus resistance in cucumber (Cucumis sativus L.) by using BSA-seq analysis. Front. Plant Sci. 2019, 10, 1583. [Google Scholar] [CrossRef]
- Win, K.T.; Vegas, J.; Zhang, C.; Song, K.; Lee, S. QTL mapping for downy mildew resistance in cucumber via bulked segregant analysis using next-generation sequencing and conventional methods. Theor. Appl. Genet. 2017, 130, 199–211. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Huang, W.; Xu, Y.; Zhou, Q.; Wang, S.; Ruan, J.; Huang, S.; Zhang, Z. A chromosome scale genome assembly of cucumber (Cucumis sativus L.). Giga Sci. 2019, 8, giz072. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Karp, P.D.; Paley, S.M.; Krummenacker, M.; Latendresse, M.; Dale, J.M.; Lee, T.J.; Kaipa, P.; Gilham, F.; Spaulding, A.; Popescu, L.; et al. Pathway Tools version 13.0: Integrated software for pathway/genome informatics and systems biology. Brief Bioinform 2010, 11, 40–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, J.M.; Popescu, L.; Karp, P.D. Machine learning methods for metabolic pathway prediction. BMC Bioinform. 2010, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Wu, S.; Bai, Y.; Sun, H.; Jiao, C.; Guo, S.; Zhao, K.; Blanca, J.; Zhang, Z.; Huang, S.; et al. Cucurbit Genomics Database (CuGenDB): A central portal for comparative and functional genomics of cucurbit crops. Nucleic Acids Res. 2019, 47, D1128–D1136. [Google Scholar] [CrossRef] [Green Version]
- Nyirahabimana, F.; Shimira, F.; Zahid, G.; Solmaz, I. Recent status of Genotyping by Sequencing (GBS) Technology in cucumber (Cucumis sativus L.): A review. Mol. Biol. Rep. 2022, 49, 5547–5554. [Google Scholar] [CrossRef]
- Malepszy, S.; Niemirowicz-Szczytt, K. Sex determination in cucumber (Cucumis sativus L.) as a model system for molecular biology. Plant Sci. 1991, 80, 39–47. [Google Scholar] [CrossRef]
- MacMurray, A.L.; Iller, C.M. Cucumber sex expression modified by 2-chloroethanephosphonic acid. Science 1968, 162, 1397–1398. [Google Scholar] [CrossRef]
- Iwahori, S.; Lyons, J.M.; William, L.S. Induced femaleness in cucumber by 2-chloroethanephosphonic acid. Nature 1969, 222, 271–272. [Google Scholar] [CrossRef]
- Yamasaki, S.; Fujii, N.; Matsuura, S.; Mizusawa, H.; Takahashi, H. The M locus and ethylene-controlled sex determination in andromonoecious cucumber plants. Plant Cell Physiol. 2001, 42, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Fujii, N.; Takahashi, H. Photoperiodic regulation of CSACS2, CS-ACS4, and CS-ERS gene expression contributes to the femaleness of cucumber flowers through diurnal ethylene production under short-day conditions. Plant Cell Environ. 2003, 26, 537–546. [Google Scholar] [CrossRef]
- Yamasaki, S.; Fujii, N.; Takahashi, H. Characterization of ethylene effects on sex determination in cucumber plants. Sex. Plant Reprod. 2003, 16, 103–111. [Google Scholar] [CrossRef]
- Mibus, H.; Tatlioglu, T. Molecular characterization and isolation of the F/f gene for femaleness in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2004, 109, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Knopf, R.; Trebitsh, T. The female-specific Cs-ACS1G gene of cucumber. A case of gene duplication and recombination between the non-sex-specific 1-aminocyclopropane-1-carboxylate synthase gene and a branched-chain amino acid transaminase gene. Plant Cell. Physiol. 2006, 47, 1217–1228. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Fujii, N.; Miyazawa, Y.; Yamasaki, S.; Matsuura, S.; Mizusawa, H.; Fujita, Y.; Takahashi, H. Correlation between development of female flower buds and expression of the Cs-ACS2 gene in cucumber plants. J. Exp. Bot. 2007, 58, 2897–2907. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, S.; Liu, S.; Pan, J.; Zhang, Z.; Tao, Q.; Shi, Q.; Jia, Z.; Zhang, W.; Chen, H.; et al. Molecular isolation of the M gene suggests that a conserved-residue conversion induces the formation of bisexual flowers in cucumber plants. Genetics 2009, 182, 1381–1385. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, S.; Tao, Q.; Pan, J.; Si, L.; Gong, Z.; Cai, R. A putative positive feedback regulation mechanism in CsACS2 expression suggests a modified model for sex determination in cucumber (Cucumis sativus L.). J. Exp. Bot. 2012, 63, 4475–4484. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.H.; Li, F.; Duan, Q.-H.; Han, T.; Xu, Z.-H.; Bai, S.-N. Ethylene perception is involved in female cucumber flower development. Plant J. 2010, 61, 862–872. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, G.; Li, Y.; Mo, N.; Zhang, J.; Liang, Y. Transcriptomic analysis implies that GA regulates sex expression via ethylene-dependent and ethylene-independent pathways in cucumber (Cucumis sativus L.). Front. Plant Sci. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Trebitsh, T.; Staub, J.E.; O’Neill, S.D. Identification of a 1-aminocyclopropane-1-carboxylic acid synthase gene linked to the female (F) locus that enhances female sex expression in cucumber. Plant Physiol. 1997, 113, 987–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boualem, A.; Troadec, C.; Kovalski, I.; Sari, M.-A.; Perl-Treves, R.; Bendahmane, A. A conserved ethylene biosynthesis enzyme leads to andromonoecy in two Cucumis species. PLoS ONE 2009, 4, e6144. [Google Scholar] [CrossRef]
- Boualem, A.; Troadec, C.; Camps, C.; Lemhemdi, A.; Morin, H.; Sari, M.-A.; Fraenkel-Zagouri, R.; Kovalski, I.; Dogimont, C.; Perl-Treves, R.; et al. A cucurbit androecy gene reveals how unisexual flowers develop and dioecy emerges. Science 2015, 350, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sun, J.; Li, S.; Cui, Q.; Zhang, H.; Xin, F.; Wang, H.; Lin, T.; Gao, D.; Wang, S.; et al. An ACC oxidase gene essential for cucumber carpel development. Mol. Plant 2016, 9, 1315–1327. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.; Niu, H.; Wang, Z.; Zhang, W.; Wang, H.; Wang, S.; Li, Z. Ethylene responsive factor ERF110 mediates ethylene-regulated transcription of a sex determination-related orthologous gene in two Cucumis species. J. Exp. Bot. 2018, 69, 2953–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Xu, L.; Jia, Z.; Xu, Y.; Yang, Q.; Fei, Z.; Lu, X.; Chen, H.; Huang, S. Genetic association of ETHYLENE-INSENSITIVE3-like sequence with the sex determining M locus in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2008, 117, 927–933. [Google Scholar] [CrossRef]
- Yin, T.; Quinn, J.A. Tests of a mechanistic model of one hormone regulating both sexes in Cucumis sativus (Cucurbitaceae). Am. J. Bot. 1995, 82, 1537–1546. [Google Scholar] [CrossRef]
- Pimenta Lange, M.J.; Lange, T. Gibberellin biosynthesis and the regulation of plant development. Plant Biol. 2006, 8, 281–290. [Google Scholar] [CrossRef]
- Paley, S.; Karp, P. Evaluation of computational metabolic-pathway predictions for H. pylori. Bioinformatics 2002, 18, 715–724. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.; Foerster, H.; Fulcher, C.; Kaipa, P.; Krummenacker, M.; Latendresse, M.; Paley, S.; Rhee, S.Y.; Shearer, A.; Tissier, C.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nuc. Acids Res. 2008, 36, D623–D631. [Google Scholar] [CrossRef]
- Chandler, J.W.; Cole, M.; Filer, A.; Werr, W. BIM1, a bHLH protein involved in brassinosteroid signalling controls Arabidopsis embryonic patterning via interaction with dornroschen and dornroschen-like. Plant Mol. Biol. 2009, 69, 57–68. [Google Scholar] [CrossRef]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The basic helix-loop-helix transcription factor family in plants: A genome-wide study of protein structure and functional diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pater, S.; Pham, K.; Memelink, J.; Kijne, J. RAP-1 is an Arabidopsis MYC-like R protein homologue that binds to G-box sequence motifs. Plant Mol. Biol. 1997, 34, 169–174. [Google Scholar] [CrossRef]
- Smolen, G.A.; Pawlowski, L.; Wilensky, S.E.; Bender, J. Dominant alleles of the basic helix-loop-helix transcription factor ATR2 activate stress responsive genes in Arabidopsis. Genetics 2002, 161, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.-H.; Hong, X.; Agarwal, M.; Zhu, J.-K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiribuchi, K.; Sugimori, M.; Takeda, M.; Otani, T.; Okada, K.; Onodera, H.; Ugaki, M.; Tanaka, Y.; Tomiyama-Akimoto, C.; Yamaguchi, T.; et al. RERJ1, a jasmonic acid-responsive gene from rice, encodes a basic helix-loop-helix protein. Biochem. Biophys. Res. Commun. 2004, 325, 857–863. [Google Scholar] [CrossRef]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB homologs in drought and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar]
- Friedrichsen, D.M.; Nemhauser, J.; Muramitsu, T.; Maloof, J.N.; Alonso, J.; Ecker, J.R.; Furuya, M.; Chory, J. Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics 2002, 162, 1445–1456. [Google Scholar] [CrossRef]
- Rajani, S.; Sundaresan, V. The Arabidopsis myc/bHLH gene ALCATRAZ enables cell separation in fruit dehiscence. Curr. Biol. 2001, 11, 1914–1922. [Google Scholar] [CrossRef] [Green Version]
- Liljegren, S.J.; Roeder, A.H.K.; Kempin, S.A.; Gremski, K.; Ostergaard, L.; Guimil, S.; Reyes, D.K.; Yanofsky, M.F. Control of fruit patterning in Arabidopsis by indehiscent. Cell 2004, 116, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szécsi, J.; Joly, C.; Bordji, K.; Varaud, E.; Cock, J.M.; Dumas, C. BIGPETALp, a bHLH transcription factor is involved in the control of Arabidopsis petal size. EMBO J. 2006, 25, 3912–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gremski, K.; Ditta, G.; Yanofsky, M.F. The HECATE genes regulate female reproductive tract development in Arabidopsis thaliana. Development 2007, 134, 3593–3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Yang, C.; Yuan, Z.; Zhang, D.; Gondwe, M.Y.; Ding, Z.; Liang, W.; Zhang, D.; Wilson, Z.A. The aborted microspores regulatory network is required for post-meiotic male reproductive development in Arabidopsis thaliana. Plant Cell 2010, 22, 91–107. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yang, C.; Yuan, Z.; Zhang, D.; Gondwe, M.Y.; Ding, Z.; Liang, W.; Zhang, D.; Wilson, Z.A. Comparative transcriptomic analysis of male and female flowers of monoecious Quercus suber. Front. Plant Sci. 2014, 5, 599. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Olalde, J.I.; Zúñiga-Mayo, V.M.; Serwatowska, J.; Chavez Montes, R.A.; Lozano-Sotomayor, P.; Herrera-Ubaldo, H.; Gonzalez-Aguilera, K.L.; Ballester, P.; Ripoll, J.J.; Ezquer, I.; et al. The bHLH transcription factor SPATULA enables cytokinin signaling, and both activate auxin biosynthesis and transport genes at the medial domain of the gynoecium. PLoS Genet. 2017, 13, e1006726. [Google Scholar] [CrossRef] [Green Version]
- Sharples, S.C.; Nguyen-Phan, T.C.; Fry, S.C. Xyloglucan endotransglucosylase/hydrolases (XTHs) are inactivated by binding to glass and cellulosic surfaces, and released in active form by a heat-stable polymer from cauliflower florets. J. Plant Physiol. 2017, 218, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Colf, L.A.; Karren, M.A.; Sandrin, V.; Rodesch, C.K.; Sundquist, W.I. Human ESCRT-III and VPS4 proteins are required for centrosome and spindle maintenance. Proc. Natl. Acad. Sci. USA 2010, 107, 12889–12894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Hu, W.; Vierstra, R.D. The vacuolar protein sorting-38 subunit of the arabidopsis phosphatidylinositol-3-kinase complex plays critical roles in autophagy, endosome sorting, and gravitropism. Front. Plant Sci. 2018, 9, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Yan, S.; Yang, W.; Li, Y.; Xia, M.; Chen, Z.; Wang, Q.; Yan, L.; Song, X.; Liu, R.; et al. Transcriptomic analysis reveals the roles of microtubule-related genes and transcription factors in fruit length regulation in cucumber (Cucumis sativus L.). Sci Rep. 2015, 26, 8031. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.Q.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Asami, T.; Yu, J.Q. A role of brassinosteroids in early fruit development in cucumber. J. Exp. Bot. 2008, 59, 2299–2308. [Google Scholar] [CrossRef]
- Sabag, M.; Ben Ari, G.; Zviran, T.; Biton, I.; Goren, M.; Dahan, Y.; Sadka, A.; Irihimovitch, V. PaKRP, a cyclin-dependent kinase inhibitor from avocado, may facilitate exit from the cell cycle during fruit growth. Plant Sci. 2013, 213, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, K.; Chory, J. Brassinosteroid signal transduction: Still casting the actors. Curr. Opin. Plant Biol. 2000, 3, 79–84. [Google Scholar] [CrossRef]
- Zhou, Y.; Hu, L.; Jiang, L.; Liu, S. Genome-wide identification, characterization, and transcriptional analysis of the metacaspase gene family in cucumber (Cucumis sativus L.). Genome 2018, 61, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Takatsuji, H. Zinc-finger proteins: The classical zinc finger emerges in contemporary plant science. Plant Mol. Biol. 1999, 39, 1073–1078. [Google Scholar] [CrossRef]
- Colasanti, J.; Yuan, Z.; Sundaresan, V. The indeterminate gene encodes a zinc finger protein and regulates a leaf-generated signal required for the transition to flowering in maize. Cell 1998, 93, 593–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizhsky, L.; Davletova, S.; Liang, H.; Mittler, R. The zinc finger protein Zat12 is required for cytosolic ascorbate peroxidase 1 expression during oxidative stress in Arabidopsis. J. Biol. Chem. 2004, 279, 11736–11743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, H.; Maruyama, K.; Sakuma, Y.; Meshi, T.; Iwabuchi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Cys2/His2-type zinc-finger proteins function as transcription repressors under drought, cold, and high-salinity stress conditions. Plant Physiol. 2004, 136, 2734–2746. [Google Scholar] [CrossRef] [Green Version]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Radkova, M.; Revalska, M.; Kertikova, D.; Iantcheva, A. Zinc finger CCHC-type protein related with seed size in model legume species Medicago truncatula. Biotechnol. Biotechnol. Equip. 2019, 33, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Marchal, C.; Zhang, J.; Zhang, P.; Fenwick, P.; Steuernagel, B.; Adamski, N.M.; Boyd, L.; McIntosh, R.; Wulff, B.B.H.; Berry, S.; et al. BED-domain-containing immune receptors confer diverse resistance spectra to yellow rust. Nat. Plants 2018, 4, 662–668. [Google Scholar] [CrossRef]
- Knip, M.; de Pater, S.; Hooykaas, J.J. The Sleeper genes: A transposase-derived angiosperm-specific gene family. BMC Plant Biol. 2012, 12, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smaczniak, C.; Immink, R.G.H.; Muiño, J.M.; Blanvillain, R.; Busscher, M.; Busscher-Lange, J.; Dinh, Q.D.; Liu, S.; Westphal, A.H.; Boeren, S.; et al. Characterization of MADS-domain transcription factor complexes in Arabidopsis flower development. Proc. Natl. Acad. Sci. USA 2012, 109, 1560–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theissen, G.; Melzer, R. Molecular mechanisms underlying origin and diversification of the angiosperm flower. Ann. Bot. 2007, 100, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Urasaki, N.; Tarora, K.; Shudo, A.; Ueno, H.; Tamaki, M.; Miyagi, N.; Adaniya, S.; Matsumura, H. Digital transcriptome analysis of putative sex-determination genes in papaya (Carica papaya). PLoS ONE 2012, 7, e40904. [Google Scholar] [CrossRef]
- Boualem, A.; Fergany, M.; Fernandez, R.; Troadec, C.; Martin, A.; Morin, H.; Sari, M.-A.; Collin, F.; Flowers, J.M.; Pitrat, M.; et al. A conserved mutation in an ethylene biosynthesis enzyme leads to andromonoecy in melons. Science 2008, 321, 836–838. [Google Scholar] [CrossRef]
- Jones, M.L. Ethylene biosynthetic genes are differentially regulated by ethylene and ACC in carnation styles. Plant Growth Regul. 2003, 40, 129–138. [Google Scholar] [CrossRef]
- Lin, Z.; Zhong, S.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.O.; Yang, S.F. Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene. Proc. Natl. Acad. Sci. USA 1979, 76, 170–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, W.; Chen, F.; Cheng, Y.; Huang, X.; Zou, B.; Wang, Y.; Xu, W.; Qu, S. Genome-Wide Identification and Characterization of Members of the ACS Gene Family in Cucurbita maxima and Their Transcriptional Responses to the Specific Treatments. Int. J. Mol. Sci. 2022, 23, 8476. [Google Scholar] [CrossRef]
- García, A.; Aguado, E.; Garrido, D.; Martínez, C.; Jamilena, M. Two androecious mutations reveal the crucial role of ethylene receptors in the initiation of female flower development in Cucurbita pepo. Plant J. 2020, 103, 1548–1560. [Google Scholar] [CrossRef]
- Ahmadizadeh1, M.; Chen, J.-T.; Hasanzadeh, S.; Ahmar, S.; Heidari, P. Insights into the genes involved in the ethylene biosynthesis pathway in Arabidopsis thaliana and Oryza sativa. J. Genet. Eng. Biotechnol. 2020, 18, 62. [Google Scholar] [CrossRef]
- O’Neill, S.D. Pollination regulation of flower development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 547–574. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, K.; Yoshioka, T.; Hashiba, T.; Satoh, S. Role of the gynoecium in natural senescence of carnation (Dianthus caryophyllus L.) flowers. J. Exp. Bot. 2000, 51, 2067–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Evensen, K.B.; Kao, T.-H. Ethylene synthesis and floral senescence following compatible and incompatible pollinations in Petunia inflata. Plant Physiol. 1992, 99, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Trebitsh, T.; Rudich, J.; Riov, J. Auxin, biosynthesis of ethylene and sex expression in cucumber (Cucumis sativus). Plant Growth Regul. 1987, 5, 105–113. [Google Scholar] [CrossRef]
- Xue, J.Q.; Li, Y.H.; Tan, H.; Yang, F.; Ma, N.; Gao, J.P. Expression of ethylene biosynthetic and receptor genes in rose floral tissues during ethylene-enhanced flower opening. J. Exp. Bot. 2008, 59, 2161–2169. [Google Scholar] [CrossRef] [Green Version]
- Ha, S.T.T.; Lim, J.H.; In, B.C. Differential expression of ethylene signalling and biosynthesis genes in floral organs between ethylene sensitive and insensitive rose cultivars. Hortic. Sci. Technol. 2019, 37, 227–237. [Google Scholar]
- Aguado, E.; García, A.; Iglesias-Moya, J.; Romero, J.; Wehner, T.C.; Gómez-Guillamón, M.L.; Picó, B.; Garcés-Claver, A.; Martínez, C.; Jamilena, M. Mapping a partial andromonoecy locus in Citrullus lanatus using BSA-Seq and GWAS approaches. Front. Plant Sci. 2020, 11, 1243. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.-L.; Peng, Y.-B.; Cui, J.-X.; Gu, H.-T.; Xu, L.-Y.; Li, Y.-Q.; Xu, Z.-H.; Bai, S.-N. Developmental analyses reveal early arrests of the spore-bearing parts of reproductive organs in unisexual flowers of cucumber (Cucumis sativus L.). Planta 2004, 220, 230–240. [Google Scholar] [CrossRef]
- Byers, R.E.; Baker, L.R.; Sell, H.M.; Herner, R.C.; Dilley, D.R. Ethylene: A natural regulator of sex expression of Cucumis melo L. Proc. Natl. Acad. Sci. USA 1972, 69, 717–720. [Google Scholar] [CrossRef] [Green Version]
- Rudich, J.; Halevy, A.H.; Kedar, N. Increase in femaleness of three cucurbits by treatment with Ethrel, an ethylene releasing compound. Planta 1969, 86, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.T.; Lu, Q.; Liu, Z.; Lv, T.X.; Li, X.Y.; Bu, H.D.; Liu, W.T.; Xu, Y.X.; Yuan, H.; Wang, A.D. Auxin-activated MdARF5 induces the expression of ethylene biosynthetic genes to initiate apple fruit ripening. New Phytol. 2020, 226, 1781–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.; Aguado, E.; Martínez, C.; Loska, D.; Beltrán, S.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. The ethylene receptors CpETR1A and CpETR2B cooperate in the control of sex determination in Cucurbita pepo. J. Exp. Bot. 2020, 71, 154–167. [Google Scholar] [CrossRef] [Green Version]
- García, A.; Aguado, E.; Parra, G.; Manzano, S.; Martínez, C.; Megías, Z.; Cebrián, G.; Romero, J.; Beltrán, S.; Garrido, D.; et al. Phenomic and genomic characterization of a mutant platform in Cucurbita pepo. Front. Plant Sci. 2018, 9, 1049. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Zhang, J.; Zhang, H.; Sun, H.; Gong, G.; Shi, J.; Tian, S.; Guo, S.; Ren, Y.; Shen, H.; et al. Mutation in the gene encoding 1-aminocyclopropane-1-carboxylate synthase 4 (CitACS4) led to andromonoecy in watermelon. J. Integr. Plant Biol. 2016, 58, 762–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, S.; Aguado, E.; Martínez, C.; Megías, Z.; García, A.; Jamilena, M. The ethylene biosynthesis gene CitACS4 regulates monoecy/andromonoecy in watermelon (Citrullus lanatus). PLoS ONE 2016, 11, e0154362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, S.; Martínez, C.; García, J.M.; Megías, Z.; Jamilena, M. Involvement of ethylene in sex expression and female flower development in watermelon (Citrullus lanatus). Plant Physiol. Biochem. 2014, 85, 96–104. [Google Scholar] [CrossRef]
- Manzano, S.; Martínez, C.; Megías, Z.; Garrido, D.; Jamilena, M. Involvement of ethylene biosynthesis and signalling in the transition from male to female flowering in the monoecious Cucurbita pepo. J. Plant Growth Regul. 2013, 32, 789–798. [Google Scholar] [CrossRef]
- Martin, A.; Troadec, C.; Boualem, A.; Rajab, M.; Fernandez, R.; Morin, H.; Pitrat, M.; Dogimont, C.; Bendahmane, A. A transposon-induced epigenetic change leads to sex determination in melon. Nature 2009, 461, 1135–1138. [Google Scholar] [CrossRef]
- Martínez, C.; Jamilena, M. To be a male or a female flower, a question of ethylene in cucurbits. Curr. Opin. Plant Biol. 2021, 59, 101981. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.; Manzano, S.; Megías, Z.; Barrera, A.; Boualem, A.; Garrido, D.; Bendahmane, A.; Jamilena, M. Molecular and functional characterization of CpACS27A gene reveals its involvement in monoecy instability and other associated traits in squash (Cucurbita pepo L.). Planta 2014, 239, 1201–1215. [Google Scholar] [CrossRef] [PubMed]


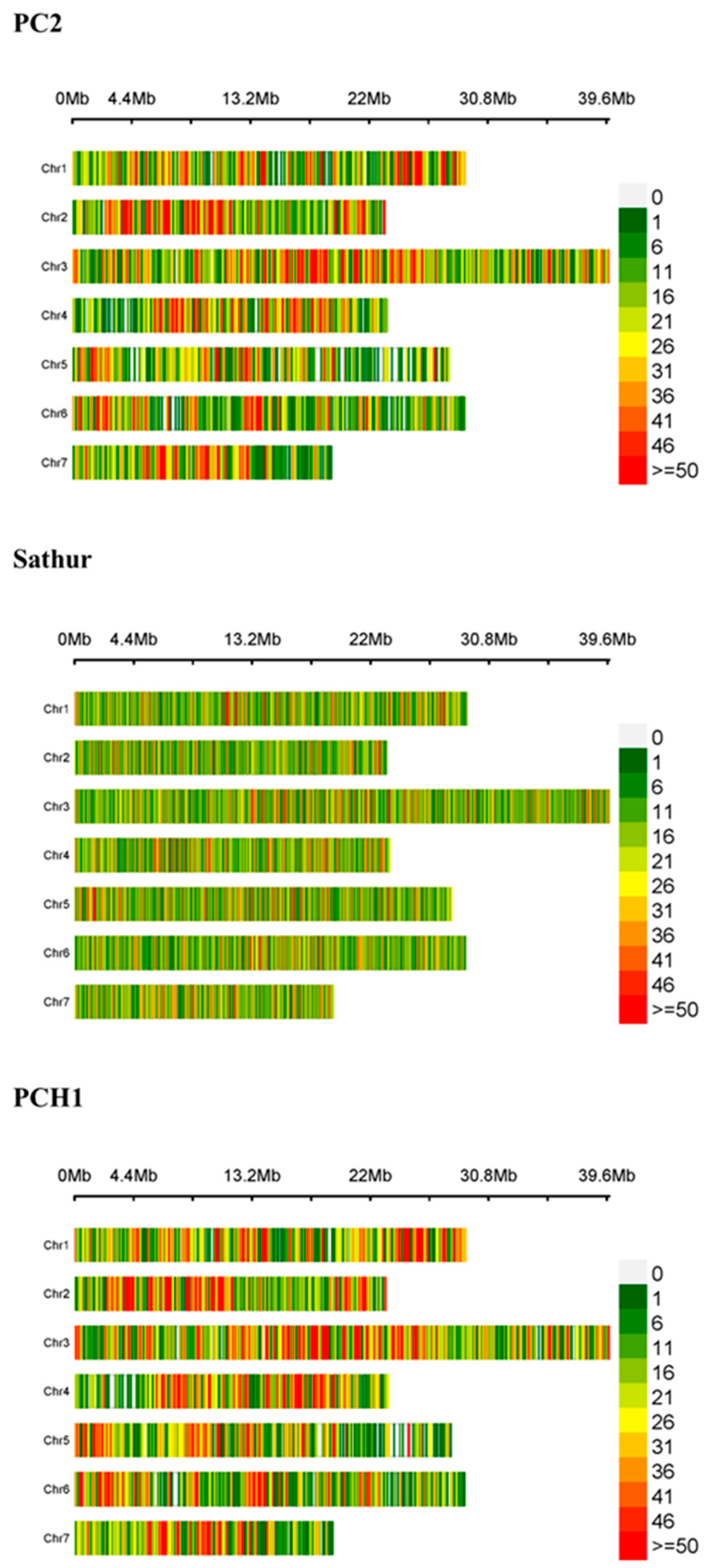{kind=link}
| Name of the Regional Genotypes | Days to First Female Flower Opening | Number of Male Flowers | Number of Female Flowers | Number of Fruits per Plant | Length of Fruit (cm) | Average Fruit Weight (g) | Yield per Plant (kg) |
|---|---|---|---|---|---|---|---|
| CBE-CS- 06 | 39.80 | 127.00 D | 26.60 | 11.00 | 21.12 | 272.27 | 3.00 |
| CBE-CS- 16 | 37.80 | 128.20 D | 28.60 | 10.60 | 23.82 | 278.84 | 3.02 |
| CBE-CS- 24 | 37.20 | 126.00 D | 30.00 | 8.60 | 19.26 | 253.42 | 2.26 |
| CBE-CS- 32 | 38.40 | 129.40 D | 29.80 | 11.20 | 22.02 | 298.80 | 3.48 |
| CBE-CS- 33 | 37.40 | 125.80 D | 26.40 | 10.40 | 20.42 | 286.48 | 3.00 |
| CBE-CS- 35 | 40.20 | 120.40 D | 29.60 | 9.00 | 12.62 | 215.44 | 1.06 |
| CBE-CS- 36 | 38.40 | 126.80 D | 28.00 | 9.40 | 16.68 | 258.68 | 1.54 |
| AVCU 1202 | 33.40 | 9.40 | 43.80 D | 5.60 | 14.44 | 250.66 | 1.38 |
| AVCU 1203 | 34.20 | 13.40 | 51.40 D | 6.60 | 15.78 | 242.16 | 1.62 |
| AVCU 1205 | 37.60 | 12.60 | 54.20 D | 5.40 | 13.70 | 188.34 | 1.00 |
| AVCU 1206 | 30.00 | 14.00 | 64.00 D | 8.20 | 17.38 | 225.00 | 1.87 |
| AVCU 1302 | 43.80 | 3.80 | 31.60 D | 4.00 | 11.94 | 182.00 | 1.68 |
| AVCU 1303 | 32.20 | 11.99 | 45.40 D | 7.60 | 12.04 | 192.66 | 1.50 |
| Pant C2 | 42.80 | 3.20 | 33.50 D | 8.00 | 10.40 | 236.00 | 1.82 |
| CD (5%) | 2.87 | 1.56 | 4.47 | 0.63 | 1.87 | 14.98 | 0.73 |
| S. No | Parents and Hybrids | No. of Primary Branches | No. of Male Flowers/Plant | No. of Female Flowers/Plant | Days to First Female Flower Opening | Node at Which First Male Flower Opens | Node at Which First Female Flower Opens | Fruit Length (cm) | Fruit Girth (cm) | Single Fruit Weight (kg) | No. of Fruits/Plant | Yield/Plant (kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male parents | ||||||||||||
| 1. | CBE-CS-33 | 4.78 | 85.29 | 26.42 | 35.46 | 2.56 | 3.65 | 40.85 | 19.42 | 0.141 | 11.29 | 1.591 |
| 2. | CBE-CS-16 | 6.13 | 84.36 | 27.52 | 36.63 | 2.29 | 3.57 | 43.52 | 19.64 | 0.139 | 11.22 | 1.559 |
| Female parent | ||||||||||||
| 3. | Pant C 2 | 6.38 | 8.53 | 28.34 | 31.32 | 2.42 | 3.86 | 15.32 | 9.16 | 0.172 | 16.43 | 2.83 |
| Hybrids | ||||||||||||
| 4. | TNAU PCH 1 | 6.65 | 5.16 | 31.96 | 23.26 | 2.13 | 3.22 | 16.96 | 9.22 | 0.158 | 22.68 | 3.58 |
| 5. | TNAU PCH 2 | 6.54 | 5.28 | 30.53 | 25.58 | 2.34 | 3.61 | 16.53 | 9.34 | 0.146 | 21.92 | 3.20 |
| CD | 1.23 | 6.46 | 2.26 | 2.18 | 1.10 | 1.04 | 4.62 | 0.93 | 0.69 | 2.47 | 1.68 | |
| Sample | Raw Reads | Mapped Reads | Q30 (%) | Mapping Rate (%) | Average Depth (X) | GC Content (%) |
|---|---|---|---|---|---|---|
| PCH1 | 7,318,584 | 6,074,701 | 86.91 | 83.00 | 30.80 | 34.84 |
| PC2 | 7,239,920 | 5,954,078 | 84.86 | 82.24 | 32.13 | 35.80 |
| Sathur | 7,515,922 | 3,458,720 | 83.50 | 46.02 | 25.61 | 34.98 |
| S. No. | Category | Sub-Category | PC2 | Sathur | PCH1 |
|---|---|---|---|---|---|
| 1 | Non-exonic | Upstream | 3494 | 9950 | 3841 |
| 2 | Non-exonic | Missense | 0 | 0 | 0 |
| 3 | Exonic | Stop gain | 25 | 208 | 26 |
| Stop loss | 5 | 26 | 4 | ||
| Synonymous | 876 | 17,107 | 870 | ||
| Non-synonymous | 905 | 13,739 | 890 | ||
| Unknowns | 0 | 0 | 0 | ||
| 4 | Non-exonic | Intronic | 5674 | 65,725 | 5991 |
| 5 | Non-exonic | Splicing | 15 | 93 | 10 |
| 6 | Non-exonic | Downstream | 3397 | 10,134 | 3705 |
| 7 | Non-exonic | Upstream/downstream | 516 | 1511 | 583 |
| 8 | Non-exonic | Intergenic | 24,344 | 50,281 | 26,815 |
| 9 | Non-exonic | ts | 25,208 | 109,443 | 27,432 |
| 10 | Non-exonic | tv | 14,935 | 71,565 | 16,180 |
| 11 | Non-exonic | ts/tv | 1.687 | 1.529 | 1.695 |
| 12 | Non-exonic | Het rate | 0.029 | 0.096 | 0.024 |
| Total | 40,143 | 181,008 | 43,612 |
| Chromosome | Direct Functional Gene Hits Related to Female Flowering/Fruiting Traits |
|---|---|
| 1 |
|
| 3 |
|
| 6 |
|
| 7 |
|
| Chr | Position | Reference | Allele | Gene | Annotated Position | Female | Male | Hybrid |
|---|---|---|---|---|---|---|---|---|
| 4 | 17433658 | C | C T | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | C C | T T | T T |
| 4 | 17433667 | A | A C | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | A A | C C | C C |
| 4 | 17433668 | T | T C | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | T T | C C | C C |
| 4 | 17433722 | G | G A | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | G G | A A | A A |
| 4 | 17433723 | A | A C | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | A A | C C | C C |
| 4 | 17433727 | G | G C | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | G G | C C | C C |
| 4 | 17433812 | A | A G | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | A A | G G | G G |
| 4 | 17433857 | A | A G | gene:Csa_4G499310,gene:Csa_4G499320 | intergenic | A A | G G | G G |
| 6 | 11413220 | T | T C | gene:Csa_6G168270,gene:Csa_6G169270 | intergenic | T C | T C | T C |
| 6 | 11413245 | G | G A | gene:Csa_6G168270,gene:Csa_6G169270 | intergenic | G A | G G | G G |
| 6 | 11413259 | T | T A | gene:Csa_6G168270,gene:Csa_6G169270 | intergenic | T A | T T | T T |
| 6 | 11413283 | T | T C | gene:Csa_6G168270,gene:Csa_6G169270 | intergenic | T T | T C | T C |
| 6 | 11413317 | A | A G | gene:Csa_6G168270,gene:Csa_6G169270 | intergenic | A G | A G | A G |
| 6 | 11907031 | A | A G | gene | nonsynonymous SNV | A G | A A | A A |
| 6 | 29069874 | T | T C | gene:Csa_6G538800,NONE | intergenic | T C | T T | T T |
| 6 | 29069890 | A | A G | gene:Csa_6G538800,NONE | intergenic | A G | A A | A A |
| S. No. | Category | PC2 | Sathur | PCH1 | |
|---|---|---|---|---|---|
| 1. | Exonic | Upstream | 583 | 623 | 652 |
| 2. | Stop gain | 0 | 4 | 0 | |
| 3. | Stop loss | 0 | 1 | 0 | |
| 4. | Frameshift deletion | 10 | 47 | 9 | |
| 5. | Frameshift insertion | 11 | 53 | 11 | |
| 6. | Non-frameshift deletion | 17 | 102 | 18 | |
| 7. | Non-frameshift insertion | 17 | 104 | 17 | |
| 8. | Intronic | 805 | 2616 | 850 | |
| 9. | Splicing | 2 | 14 | 4 | |
| 10. | Downstream | 544 | 501 | 610 | |
| 11. | Upstream/downstream | 81 | 80 | 92 | |
| 12. | Intergenic | 2459 | 1275 | 2760 | |
| 13. | Insertion | 2361 | 3115 | 2620 | |
| 14. | Deletion | 2374 | 3103 | 2626 | |
| Het Rate | 0.002 | 0.001 | 0.001 | ||
| Total | 4735 | 6218 | 5246 | ||
| Chr | Position 1 | Position 2 | Reference | Altered | Genotype | Annotated Position | Gene ID | Function | Location | Hit |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2,668,764 | 2,668,764 | - | GT | Homozygous | Frameshift insertion | CsaV3_1G004260 G3I-29448 | Mitogen activated protein kinase–Cell proliferation, hormonal signaling | 2,662,730–2,668,666 | Nearby |
| 2 | 4,929,433 | 4,929,441 | TTAGTAGTA | AAG | Homozygous | Non frameshift deletion | CsaV3_2G008570 G3I-43185 | Villin protein–structural protein in microvilli | 4,915,559–4,927,778 | Nearby |
| 4,929,451 | 4,929,456 | GTAGTA | - | Homozygous | ||||||
| 3 | 33,422,942 | 33,422,942 | - | AAG | Homozygous | Non frameshift insertion | CsaV3_3G040770 G3I-28088 | Nucleotide sugar transporter family protein–glycosyltransferases substrates | 33,416,748–33,422,008 | Nearby |
| 6 | 18,785,551 | 18,785,552 | TC | - | Homozygous | Frameshift deletion | CsaV3_6G304050 G3I-37940 | Histone lysine N-methyl transferase–Zinc ion binding, transferase activity protein methylation | 18,865,436–18,887,620 | Direct |
| 18,885,449 | 18,885,449 | - | TC | Frameshift insertion | ||||||
| 18,885,492 | 18,885,501 | CATTCTCCAT | - | Frameshift deletion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narayanan, M.B.; Mohan, S.V.G.; Subramanyam, P.; Venkatachalam, R.; Markkandan, K. Identification of Causal Gene-Specific SNP Markers for the Development of Gynoecious Hybrids in Cucumber (Cucumis sativa L.) Using the PathoLogic Algorithm. Horticulturae 2023, 9, 389. https://doi.org/10.3390/horticulturae9030389
Narayanan MB, Mohan SVG, Subramanyam P, Venkatachalam R, Markkandan K. Identification of Causal Gene-Specific SNP Markers for the Development of Gynoecious Hybrids in Cucumber (Cucumis sativa L.) Using the PathoLogic Algorithm. Horticulturae. 2023; 9(3):389. https://doi.org/10.3390/horticulturae9030389
Chicago/Turabian StyleNarayanan, Manikanda Boopathi, Shobhana V. Gnanapanditha Mohan, Praneetha Subramanyam, Rajasree Venkatachalam, and Kesavan Markkandan. 2023. "Identification of Causal Gene-Specific SNP Markers for the Development of Gynoecious Hybrids in Cucumber (Cucumis sativa L.) Using the PathoLogic Algorithm" Horticulturae 9, no. 3: 389. https://doi.org/10.3390/horticulturae9030389
APA StyleNarayanan, M. B., Mohan, S. V. G., Subramanyam, P., Venkatachalam, R., & Markkandan, K. (2023). Identification of Causal Gene-Specific SNP Markers for the Development of Gynoecious Hybrids in Cucumber (Cucumis sativa L.) Using the PathoLogic Algorithm. Horticulturae, 9(3), 389. https://doi.org/10.3390/horticulturae9030389