
Effects of Applying Nitrogen and Potassium on Lilium lancifolium Growth and Accumulation of Secondary Metabolites in Bulbs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Plant Materials and Treatments
2.3. Determination of Nutrient Contents
2.3.1. Sample Digestion
2.3.2. Determination of N and K Contents
2.4. Determination of Active Component Contents
2.5. Statistical Analysis
3. Results
3.1. Effects of N and K on Agronomic Traits
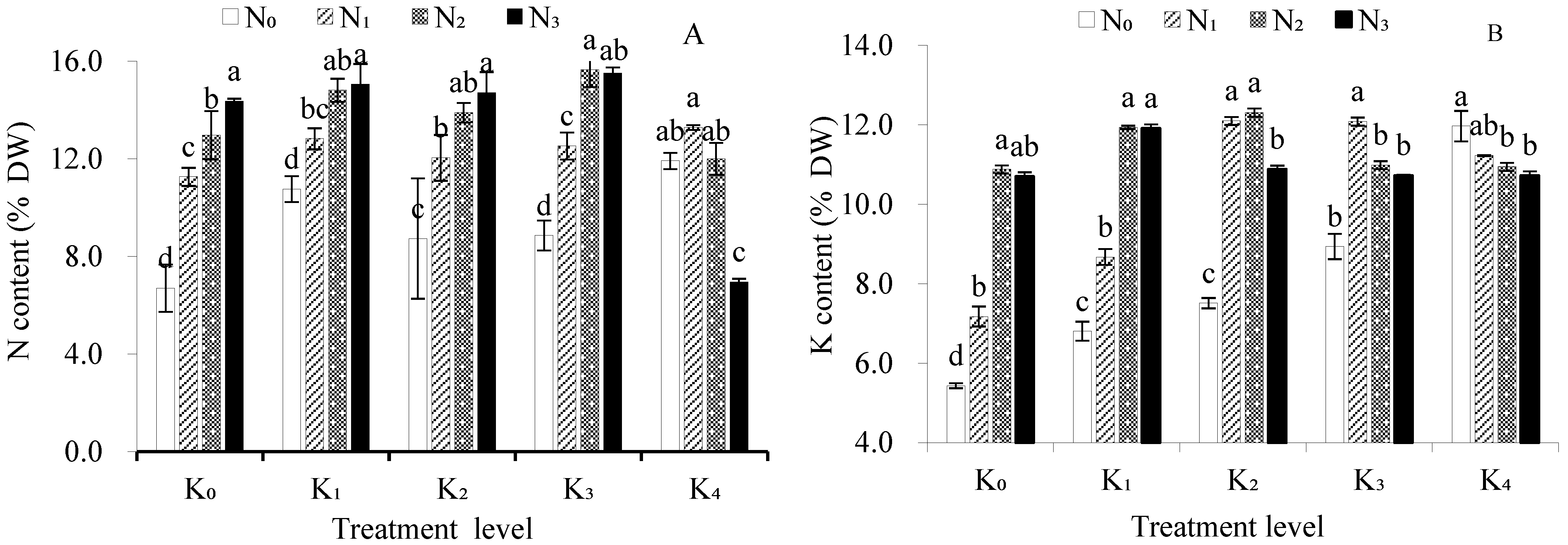
3.2. Effects of Nitrogen (N) and Potassium (K) on Total N and K Contents of Bulbs
3.3. Effects of N and K on Active Components of Bulbs
3.3.1. Effects of Individual Factors
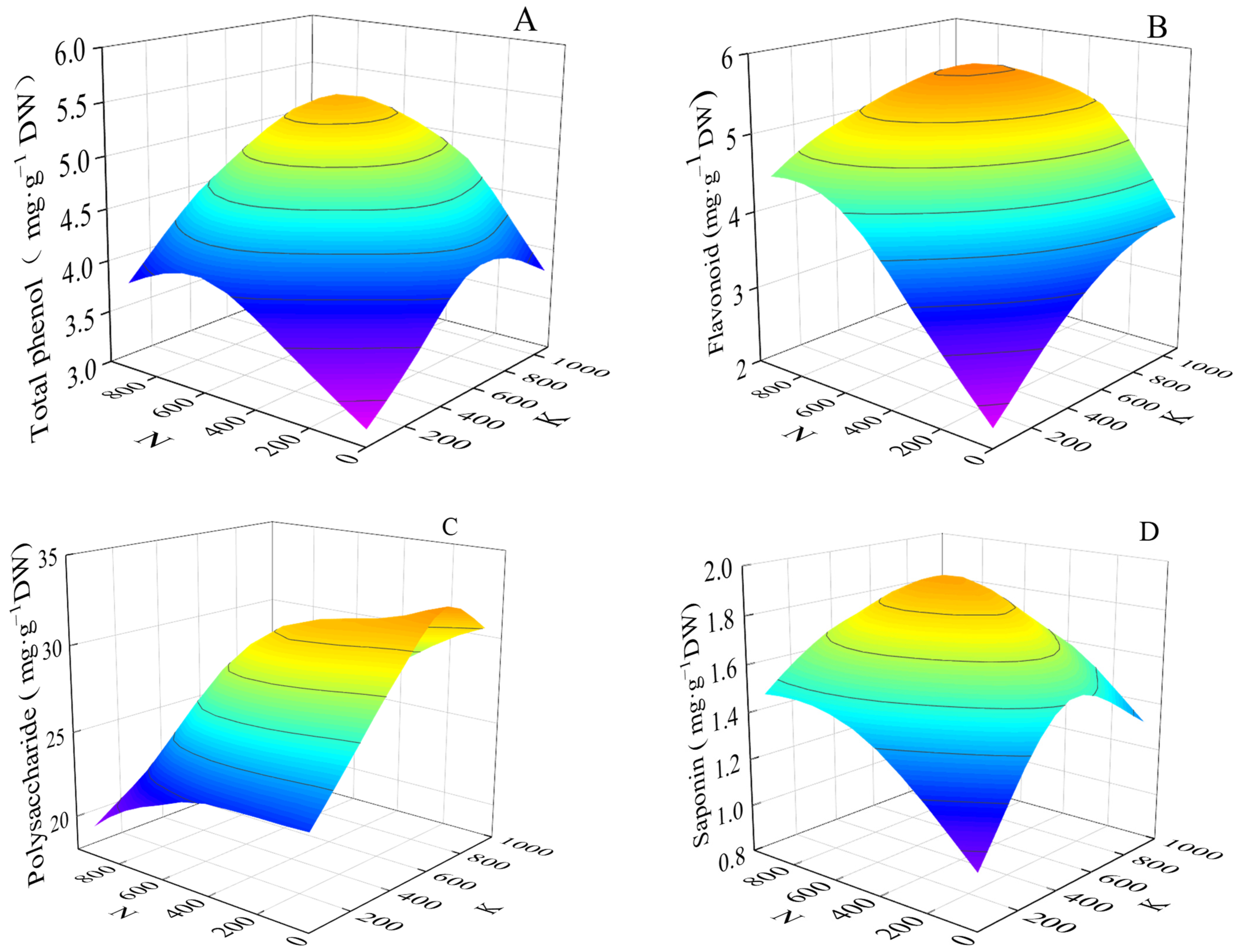
3.3.2. Effects of Two Factors
3.3.3. Optimized Combination of Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rong, L.; Lei, J.; Wang, C. Collection and evaluation of the genus Lilium resources in Northeast China. Genet. Resour. Crop Evol. 2011, 58, 115–123. [Google Scholar] [CrossRef]
- Luo, J.; Li, L.; Kong, L. Preparative separation of phenylpropenoid glycerides from the bulbs of Lilium lancifolium by high-speed counter-current chromatography and evaluation of their antioxidant activities. Food Chem. 2012, 131, 1056–1062. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, Y.; Yan, L.; Guo, Y.; Niu, L. Phenolic compounds and antioxidant activity of bulb extracts of six Lilium species native to China. Molecules 2012, 17, 9361–9378. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Wei, H.; Zhang, Y.; Guo, Z.; Qiu, Y.; Wen, L.; Xie, Z. Structural characterization of Lanzhou lily (Lilium davidii var. unicolor) polysaccharides and determination of their associated antioxidant activity. J. Sci. Food Agric. 2020, 100, 5603–5616. [Google Scholar]
- Wang, T.; Huang, H.; Zhang, Y.; Li, X.; Li, H.; Jiang, Q.; Gao, W. Role of Effective Composition on Antioxidant, Anti-inflammatory, Sedative-Hypnotic Capacities of 6 Common Edible Lilium Varieties. J. Food Sci. 2015, 80, H857–H868. [Google Scholar] [CrossRef] [PubMed]
- Sim, W.S.; Choi, S.I.; Jung, T.D.; Cho, B.Y.; Choi, S.H.; Park, S.M.; Lee, O.H. Antioxidant and anti-inflammatory effects of Lilium lancifolium bulbs extract. J. Food Biochem. 2020, 44, e13176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Y.; Luo, L.-M.; Wang, Y.-X.; Zhu, N.; Zhao, T.-J.; Qin, L. Total saponins from Lilium lancifolium: A promising alternative to inhibit the growth of gastric carcinoma cells. J. Cancer 2020, 11, 4261. [Google Scholar] [CrossRef]
- Wang, X.; Yang, L.; Liu, J.; Wang, R.; Zhang, Q.; Shan, Y.; Ding, S. Comparison of the biochemical properties and thermal inactivation of polyphenol oxidase from three lily bulb cultivars. J. Food Biochem. 2020, 44, e13431. [Google Scholar] [CrossRef]
- Chau, C.-F.; Wu, S.-H. The development of regulations of Chinese herbal medicines for both medicinal and food uses. Trends Food Sci. Technol. 2006, 17, 313–323. [Google Scholar] [CrossRef]
- You, X.; Xie, C.; Liu, K.; Gu, Z. Isolation of non-starch polysaccharides from bulb of tiger lily (Lilium lancifolium Thunb.) with fermentation of Saccharomyces cerevisiae. Carbohydr. Polym. 2010, 81, 35–40. [Google Scholar] [CrossRef]
- Barzegar, T.; Mohammadi, S.; Ghahremani, Z. Effect of nitrogen and potassium fertilizer on growth, yield and chemical composition of sweet fennel. J. Plant Nutr. 2020, 43, 1189–1204. [Google Scholar] [CrossRef]
- Martin, M. The Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 1988; p. 1250. [Google Scholar]
- Haukioja, E.; Ossipov, V.; Koricheva, J.; Honkanen, T.; Larsson, S.; Lempa, K. Biosynthetic origin of carbon-based secondary compounds: Cause of variable responses of woody plants to fertilization? Chemoecology 1998, 8, 133–139. [Google Scholar] [CrossRef]
- Raven, J.A.; Handley, L.L.; Andrews, M. Global aspects of C/N interactions determining plant–environment interactions. J. Exp. Bot. 2004, 55, 11–25. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 135–189. [Google Scholar]
- Andrews, M.; Raven, J.; Lea, P. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Jacobson, T.K.B.; da Cunha Bustamante, M.M. Effects of nutrient addition on polyphenol and nutrient concentrations in leaves of woody species of a savanna woodland in Central Brazil. J. Trop. Ecol. 2019, 35, 288–296. [Google Scholar] [CrossRef]
- Coria-Cayupán, Y.S.; Sánchez de Pinto, M.I.; Nazareno, M.A. Variations in bioactive substance contents and crop yields of lettuce (Lactuca sativa L.) cultivated in soils with different fertilization treatments. J. Agric. Food Chem. 2009, 57, 10122–10129. [Google Scholar] [CrossRef]
- Tavallali, V.; Esmaili, S.; Karimi, S. Nitrogen and potassium requirements of tomato plants for the optimization of fruit quality and antioxidative capacity during storage. J. Food Meas. Charact. 2018, 12, 755–762. [Google Scholar] [CrossRef]
- Walsh, O.S. Nitrogen and potassium fertilization for improved dryland spring wheat grain yield and quality. Crops Soils 2019, 52, 10–16. [Google Scholar] [CrossRef]
- Ma, L.; Yang, X.; Shi, Y.; Yi, X.; Ji, L.; Cheng, Y.; Ni, K.; Ruan, J. Response of tea yield, quality and soil bacterial characteristics to long-term nitrogen fertilization in an eleven-year field experiment. Appl. Soil Ecol. 2021, 166, 103976. [Google Scholar] [CrossRef]
- Schlegel, A.J.; Havlin, J.L. Irrigated grain sorghum response to 55 years of nitrogen, phosphorus, and potassium fertilization. Agron. J. 2021, 113, 464–477. [Google Scholar] [CrossRef]
- Guillén-Román, C.J.; Guevara-González, R.G.; Rocha-Guzmán, N.E.; Mercado-Luna, A.; Perez-Perez, M.C.I. Effect of nitrogen privation on the phenolics contents, antioxidant and antibacterial activities in Moringa oleifera leaves. Ind. Crops Prod. 2018, 114, 45–51. [Google Scholar] [CrossRef]
- Chung, R.-S.; Chen, C.-C.; Ng, L.-T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crops Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Jackson, M. Soil Chemical Analysis; Wisconsin Prentice Hall of India Pvt. Ltd: New Delhi, India, 1967; p. 46. [Google Scholar]
- Black, C. Methods of Soil Analysis; Part 2, Series 9; American Society of Agronomy institute publications: Madison, WI, USA, 1965; pp. 894–1372. [Google Scholar]
- Chen, F.; Huang, G. Antioxidant activity of polysaccharides from different sources of ginseng. Int. J. Biol. Macromol. 2019, 125, 906–908. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, M.; Ran, J.; Zhang, T.; Sun, H.; Dong, M.; Zhang, Z.; Zheng, H. Variation in phenolic compounds and antioxidant activity in apple seeds of seven cultivars. Saudi J. Biol. Sci. 2016, 23, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, R.-H.; Hou, D.-Y.; Li, T.-C.; Liu, X.-Y. Simultaneous determination of total flavone and total saponin in Gynostemma pentaphyllum by signal multiplier spectrophotometry. Spectrosc Spect Anal. 2006, 26, 1753–1756. [Google Scholar]
- Zhang, Y.; Chen, Y.; Lu, H.; Kong, X.; Dai, J.; Li, Z.; Dong, H. Growth, lint yield and changes in physiological attributes of cotton under temporal waterlogging. Field Crops Res. 2016, 194, 83–93. [Google Scholar] [CrossRef]
- Xia, Y.; Jiang, C.; Wang, X.; Chen, F. Studies on potassium uptake and use efficiency of different cotton (Gossypium hirsutum L.) genotypes by grafting. J. Food Agric. Environ. 2013, 11, 472–476. [Google Scholar]
- Abou El-Magd, M.; Zaki, M.; Eldewiny, C. Effect of planting dates and different levels of potassium fertilizer on growth, yield and chemical composition of sweet fennel cultivars under newly reclaimed sandy soil conditions. J. Am. Sci. 2010, 6, 89–105. [Google Scholar]
- Salvagiotti, F.; Miralles, D.J. Radiation interception, biomass production and grain yield as affected by the interaction of nitrogen and sulfur fertilization in wheat. Eur. J. Agron. 2008, 28, 282–290. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, G.; Jiao, X.; Zhou, G. Effects of nitrogen application and planting density on growth and yield of Sesbania pea grown in saline soil. Curr. Sci. 2019, 116, 113891. [Google Scholar] [CrossRef]
- Hamdi, W.; Rihab, B.; Quertatani, S.; Gharbi, A. Effect of nitrogen and potassium levels on the potatoes growth in the Chott Mariem region of Tunisia. Aust. J. Exp. Agric. 2018, 28, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Lin, W. Effects of nitrogen fertilizer rates on the growth and nutrient utilization of calla lily intercropped with rubber trees. Soil Tillage Res. 2021, 211, 105031. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; El-Nakhel, C.; Mori, M. Nitrogen use and uptake efficiency and crop performance of baby spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianella locusta L.) grown under variable sub-optimal N regimes combined with plant-based biostimulant application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Wang, Y.; Ye, Y.; Zhen, S.; Zhou, B.; Wang, Y.; Hu, Y.; Zhao, Y.; Huang, Y. Grain yields and nitrogen use efficiencies in different types of stay-green maize in response to nitrogen fertilizer. Plants 2020, 9, 474. [Google Scholar] [CrossRef] [PubMed]
- Sui, N.; Wang, L.; Sun, J.; Wu, J.; Chen, M.; Zhang, S.; Yuan, Q.; Kai, G. Estimating Fritillaria thunbergii Miq. yield, quality, and potassium use efficiency in response to potassium application rate. Ind. Crops Prod. 2021, 164, 113409. [Google Scholar] [CrossRef]
- Hu, A.; Tang, T.; Liu, Q. Nitrogen use efficiency in different rice-based rotations in southern China. Nutr. Cycl. Agroecosyst. 2018, 112, 75–86. [Google Scholar] [CrossRef]
- Olde Venterink, H. Legumes have a higher root phosphatase activity than other forbs, particularly under low inorganic P and N supply. Plant Soil 2011, 347, 137–146. [Google Scholar] [CrossRef]
- Pelinganga, O.; Mphosi, M. Optimum NPK fertilizer requirement for Amaranthus hybridus leafy vegetable under greenhouse conditions. Res. Crops 2019, 20, 353–356. [Google Scholar]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Alhasan, A.S.; Abbas, M.K.; Al-Ameri, M.; Al-Ameri, D.T. Growth and Yield Response of Basil (Ocimum basilicum L.) to Different Rates of Urea Fertilizer under Field Conditions; IOP Publishing: Bristol, UK, 2020; p. 12044. [Google Scholar]
- Hu, W.; Ren, T.; Meng, F.; Cong, R.; Li, X.; White, P.J.; Lu, J. Leaf photosynthetic capacity is regulated by the interaction of nitrogen and potassium through coordination of CO2 diffusion and carboxylation. Physiol. Plant. 2019, 167, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Wakeel, A. Potassium–sodium interactions in soil and plant under saline-sodic conditions. J. Plant Nutr. Soil Sci. 2013, 176, 344–354. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Factors | Constant Interval | Levels | ||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||
| N | 304.9 | 0 | 304.90 | 609.80 | 914.70 | – |
| K | 261.67 | 0 | 261.67 | 523.34 | 785.01 | 1046.68 |
| N | K | Plant Height (cm) | Stem Diameter (mm) | Leaf Number (No.) | Total Chlorophyll (mg g−1) | Bulb Circumference (cm) |
|---|---|---|---|---|---|---|
| 0 | 0 | 127.78 ± 1.52 gh | 6.43 ± 0.67 def | 107.17 ± 2.46 d | 46.50 ± 2.07 h | 12.12 ± 0.58 de |
| 1 | 128.47 ± 2.46 fgh | 6.86 ± 0.30 bcdef | 109.83 ± 2.37 cd | 46.00 ± 3.14 h | 12.97 ± 0.84 bcd | |
| 2 | 132.45 ± 1.16 cdefg | 7.08 ± 0.73 abcdef | 112.50 ± 2.29 bcd | 51.85 ± 3.44 g | 12.88 ± 0.82 bcd | |
| 3 | 135.57 ± 1.92 bcde | 6.74 ± 0.81 bcdef | 114.67 ± 1.59 abcd | 52.98 ± 2.34 fg | 12.90 ± 1.14 bcd | |
| 4 | 133.45 ± 1.97 cdef | 6.31 ± 0.60 f | 120.83 ± 0.97 ab | 58.16 ± 3.49 ab | 12.87 ± 0.42 bcd | |
| 1 | 0 | 132.17 ± 1.28 defgh | 6.77 ± 0.48 bcdef | 117.33 ± 1.85 abc | 55.43 ± 2.85 cde | 11.95 ± 0.40 de |
| 1 | 134.60 ± 1.56 cdef | 6.81 ± 0.32 bcdef | 118.67 ± 1.77 abc | 56.25 ± 3.40 abcd | 12.43 ± 0.72 cde | |
| 2 | 133.55 ± 1.00 cdefg | 7.30 ± 0.42 abcd | 117.00 ± 2.16 abc | 56.61 ± 3.45 abcd | 12.98 ± 0.42 bcd | |
| 3 | 135.68 ± 2.10 bcde | 7.05 ± 0.35 bcdef | 114.00 ± 0.89 abcd | 55.97 ± 3.13 bcde | 13.78 ± 0.94 ab | |
| 4 | 132.50 ± 2.13 cdefg | 7.11 ± 0.60 abcdef | 114.67 ± 1.28 abcd | 57.76 ± 3.80 abc | 12.92 ± 0.30 bcd | |
| 2 | 0 | 134.53 ± 0.99 cdef | 7.18 ± 0.36 abcdef | 119.17 ± 1.05 ab | 53.94 ± 2.76 efg | 12.55 ± 1.02 bcd |
| 1 | 136.78 ± 1.68 bcd | 7.25 ± 0.51 abcde | 122.17 ± 1.60 a | 56.57 ± 2.91 abcd | 13.05 ± 0.85 bcd | |
| 2 | 138.78 ± 1.92 abc | 7.35 ± 0.91 abc | 118.33 ± 1.34 abc | 58.35 ± 3.34 a | 14.23 ± 0.68 a | |
| 3 | 143.17 ± 3.17 a | 7.26 ± 0.44 abcde | 117.33 ± 3.03 abc | 52.56 ± 2.37 g | 13.60 ± 0.32 abc | |
| 4 | 141.55 ± 2.23 ab | 7.92 ± 0.81 a | 112.50 ± 1.28 bcd | 53.19 ± 4.18 fg | 13.48 ± 1.02 abc | |
| 3 | 0 | 129.00 ± 2.14 efgh | 7.44 ± 0.62 ab | 121.33 ± 2.63 ab | 56.87 ± 2.26 abcd | 11.37 ± 0.57 e |
| 1 | 130.83 ± 1.61 defg | 7.32 ± 0.34 abc | 118.00 ± 3.08 abc | 56.78 ± 3.01 abcd | 12.87 ± 0.72 bcd | |
| 2 | 128.17 ± 2.86 fgh | 6.74 ± 0.27 bcde | 117.00 ± 2.87 abc | 54.89 ± 2.57 def | 13.75 ± 0.56 ab | |
| 3 | 126.50 ± 2.83 h | 6.49 ± 0.45 cdef | 117.67 ± 1.52 abc | 52.58 ± 2.12 g | 13.43 ± 0.29 abc | |
| 4 | 126.67 ± 1.98 h | 6.40 ± 0.31 ef | 114.17 ± 1.66 abcd | 53.18 ± 3.66 fg | 12.70 ± 0.42 bcd |
| Traits | Plant Height | Stem Diameter | Leaf Number | Total Chlorophyll | Bulb Circumference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Factors | F | P | F | P | F | P | F | P | F | P |
| N | 1.79 | 0.07 | 3.77 * | 0.04 | 2.47 | 0.07 | 3.12 * | 0.04 | 1.79 | 0.14 |
| K | 3.25 * | 0.03 | 1.61 | 0.12 | 0.96 | 0.38 | 4.29 * | 0.03 | 2.97 * | 0.04 |
| N * K | 3.47 * | 0.03 | 0.72 | 0.18 | 0.54 | 0.36 | 1.03 | 0.04 | 2.91 * | 0.04 |
| N | K | Plant Height | Stem Diameter | Leaf Number | Total Chlorophyll | Bulb Circumference | |
|---|---|---|---|---|---|---|---|
| N | 1 | ||||||
| K | 0.00 | 1 | |||||
| Plant Height | 0.50 * | 0.31 | 1 | ||||
| Stem Diameter | 0.27 | −0.07 | 0.62 ** | 1 | |||
| Leaf Number | 0.47 * | −0.10 | 0.59 ** | 0.17 | 1 | ||
| Chlorophyll | 0.33 | 0.20 | 0.52 * | 0.28 | 0.78 ** | 1 | |
| Bulb Circumference | 0.14 | 0.56 * | 0.43 | 0.08 | −0.05 | 0.13 | 1 |
| N | K | N Content | K Content | |
|---|---|---|---|---|
| N | 1 | |||
| K | 0.00 | 1 | ||
| N content | 0.53 * | 0.55 * | 1 | |
| K content | 0.44 | 0.55 * | 0.42 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Bu, H.; Wang, H.; Zhang, P.; Jin, L. Effects of Applying Nitrogen and Potassium on Lilium lancifolium Growth and Accumulation of Secondary Metabolites in Bulbs. Horticulturae 2023, 9, 396. https://doi.org/10.3390/horticulturae9030396
Wang Y, Bu H, Wang H, Zhang P, Jin L. Effects of Applying Nitrogen and Potassium on Lilium lancifolium Growth and Accumulation of Secondary Metabolites in Bulbs. Horticulturae. 2023; 9(3):396. https://doi.org/10.3390/horticulturae9030396
Chicago/Turabian StyleWang, Yunxia, Hubai Bu, Han Wang, Ping Zhang, and Lei Jin. 2023. "Effects of Applying Nitrogen and Potassium on Lilium lancifolium Growth and Accumulation of Secondary Metabolites in Bulbs" Horticulturae 9, no. 3: 396. https://doi.org/10.3390/horticulturae9030396
APA StyleWang, Y., Bu, H., Wang, H., Zhang, P., & Jin, L. (2023). Effects of Applying Nitrogen and Potassium on Lilium lancifolium Growth and Accumulation of Secondary Metabolites in Bulbs. Horticulturae, 9(3), 396. https://doi.org/10.3390/horticulturae9030396