
Comparative Analysis of Gene Expression between Early Maturation Mutant ‘Beni Shogun’ and ‘Fuji’ Cultivars during Fruit Development and Ripening


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Fruit Quality Analysis and Internal Ethylene Measurement
2.3. RNA Extraction
2.4. Reverse Transcription
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
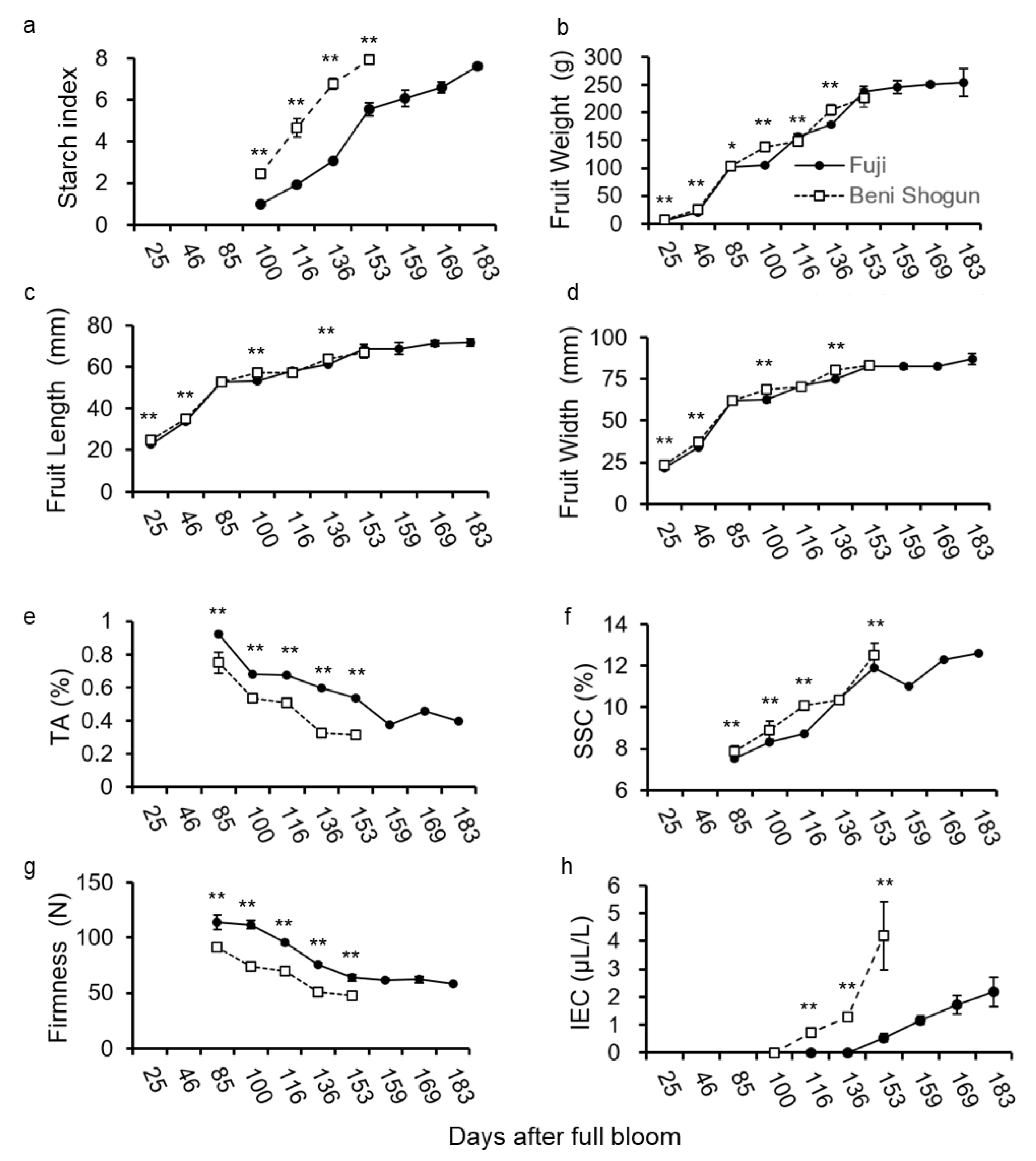
3.1. Fruit Quality Analysis during Maturation of ‘Beni Shogun’ and ‘Fuji’
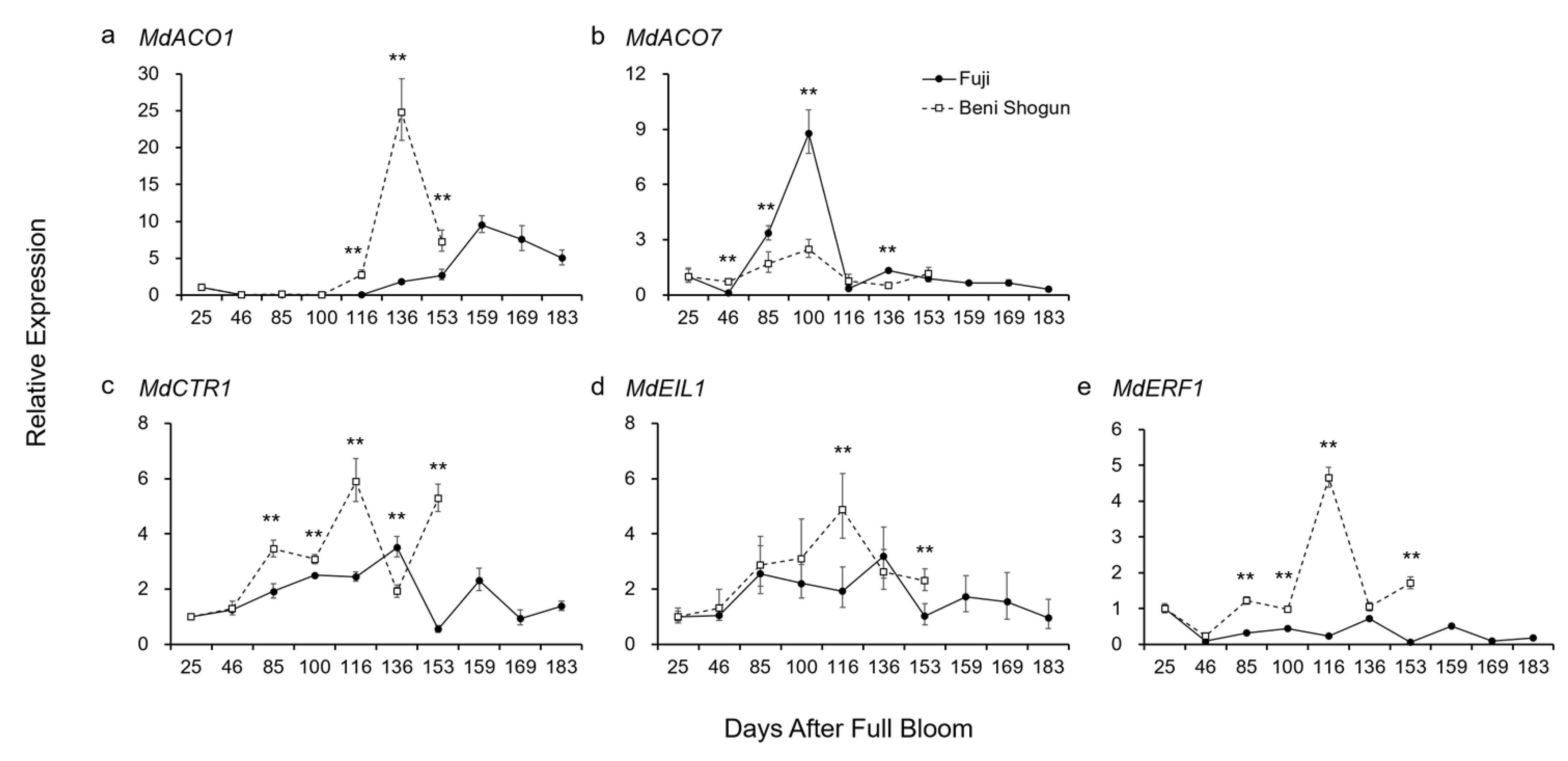
3.2. Comparison of Gene Expression Associated with Ethylene
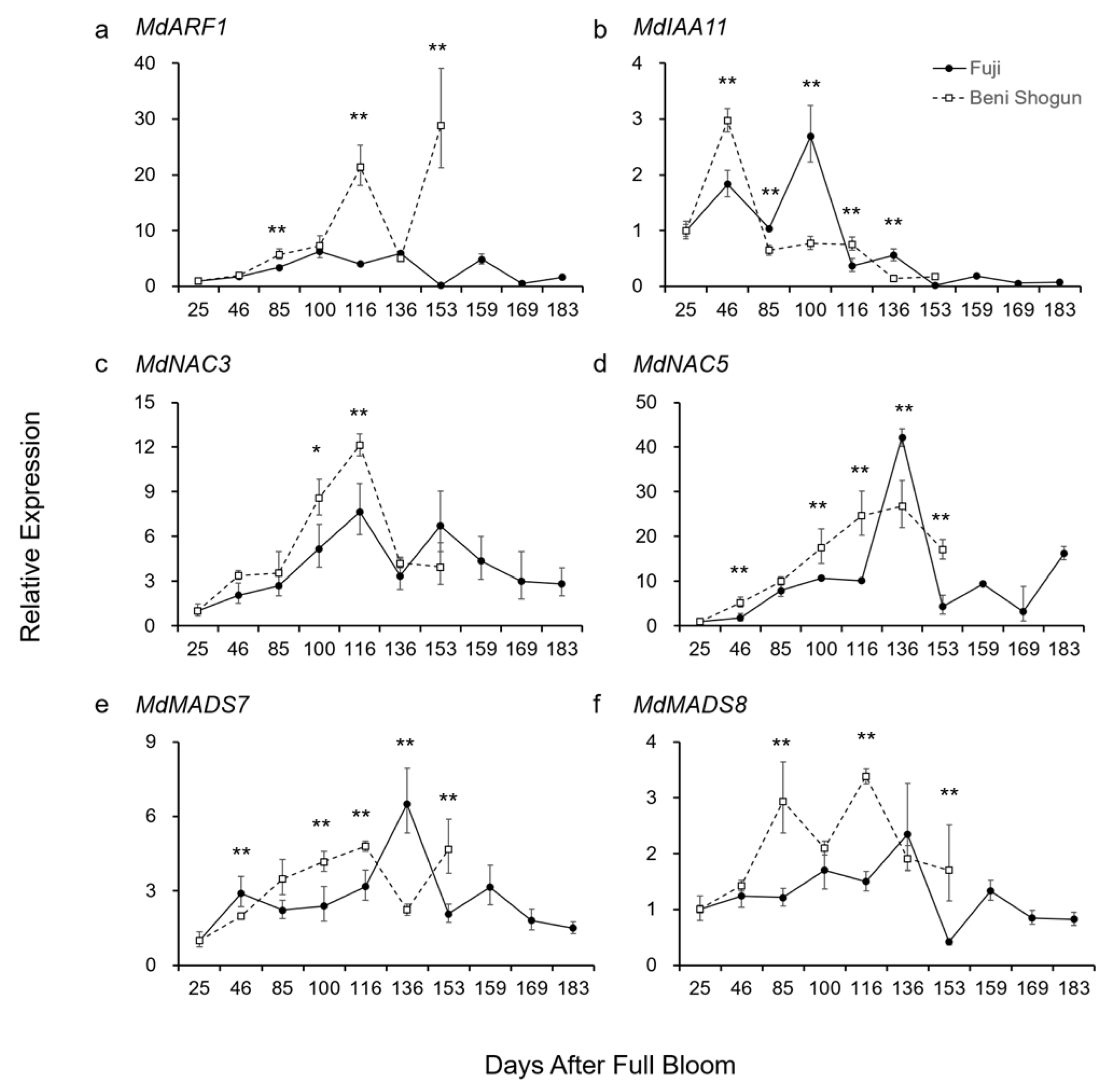
3.3. Comparison of Gene Expression Linked to Auxin and Developmental Regulation
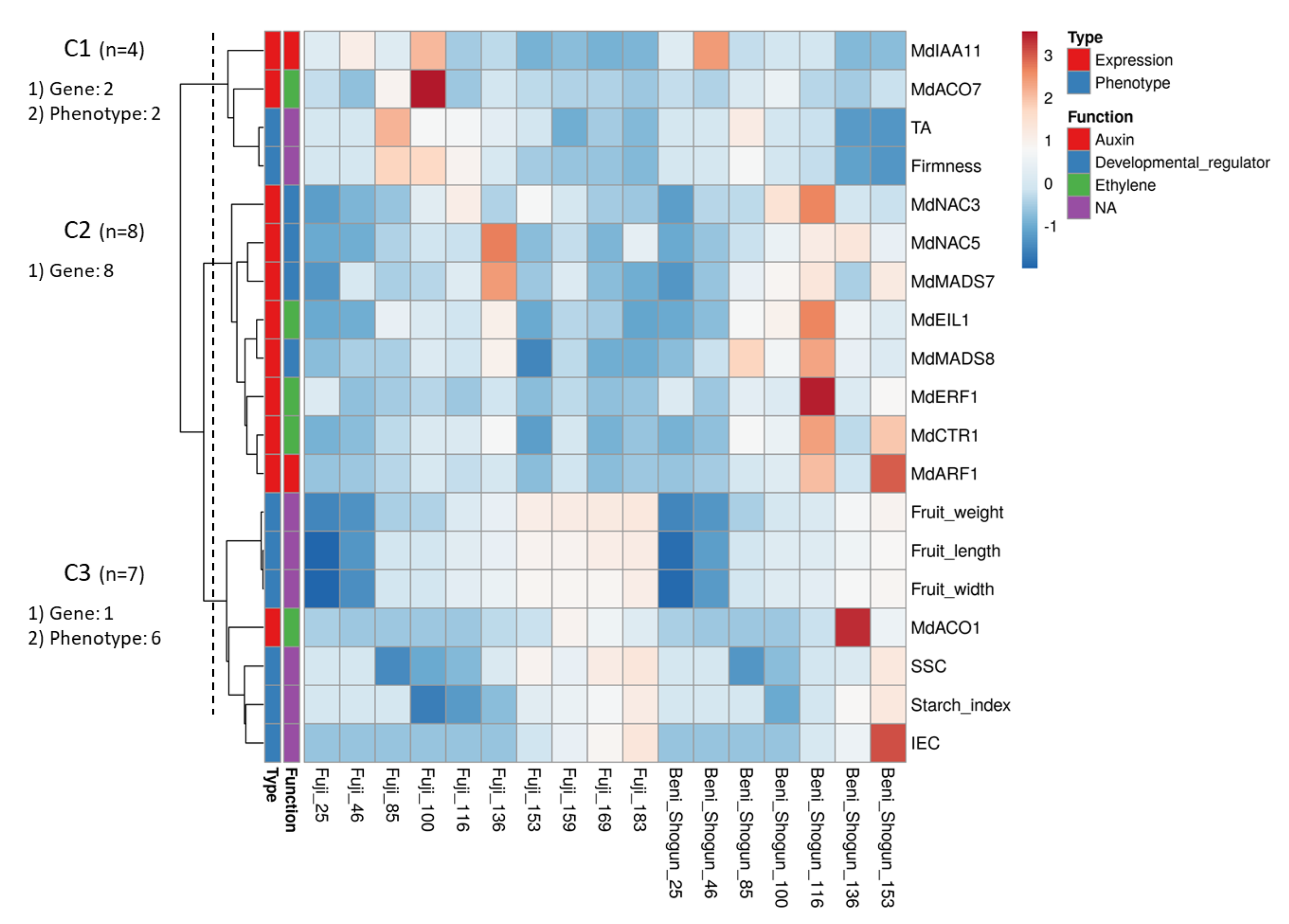
3.4. Clustering Analysis of Phenotyping Trait and Expression Data
4. Discussion
4.1. Fruit Quality and Ripening Behavior Difference between Two Cultivars
4.2. Putative Molecular Mechanism behind Distinct Fruit Development, Maturation, and Ripening
4.3. Clustering Analysis Reveals Hidden Relationships between Gene Expression Patterns and Fruit Phenotypes in Two Apple Varieties
4.4. A Working Model for How ‘Beni Shogun’ and ‘Fuji’ Showed Different Fruit Development, Maturation, and Ripening Behaviors
- (1)
- Multiple signaling pathways, hormones, and transcription factors regulate fruit development and ripening in apples.
- (2)
- Ethylene biosynthesis and signal transduction play an essential role in the ripening process of both ‘Beni Shogun’ and ‘Fuji’ apples.
- (3)
- The higher expression of MdACO1 in ‘Beni Shogun’ suggests that this variety produces more ethylene than ‘Fuji’, which could explain the difference in ethylene production between the two varieties.
- (4)
- Auxin also plays a significant role in fruit maturation, and its interplay with ethylene is crucial for fruit development.
- (5)
- The regulation of auxin transport and homeostasis-related genes including MdARF1 and MdIAA11 affects the ripening time in apples by potentially activating the ethylene biosynthesis pathway.
- (6)
- The higher expression levels of MdNAC3, MdNAC5, MdMADS7, and MdMADS8 in ‘Beni Shogun’ and MdNAC5 in ‘Fuji’ suggest that these transcription factors may play a role in the differential regulation of fruit ripening between the two varieties.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schoen, D.J.; Schultz, S.T. Somatic Mutation and Evolution in Plants. Ann. Rev. Ecol. Evol. Syst. 2019, 50, 49–73. [Google Scholar] [CrossRef]
- Foster, T.M.; Aranzana, M.J. Attention sports fans! The far-reaching contributions of bud sport mutants to horticulture and plant biology. Hortic. Res. 2018, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, I.; Liang, D.; Xu, K. Transcriptome analysis of an apple (Malus x domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.S.; Jiang, N.; Zhang, Y.G.; Wang, M.; Ren, J.P.; Tao, J.M. A SNP in the promoter region of theVvmybA1 gene is responsible for differences in grape berry color between two related bud sports of grape. Plant Growth Regul. 2017, 82, 457–465. [Google Scholar] [CrossRef]
- Nashima, K.; Terakami, S.; Nishitani, C.; Yamamoto, T.; Habu, T.; Takahashi, H.; Nakazono, M.; Isuzugawa, K.; Hanada, T.; Takashina, T.; et al. Transcriptome analysis of flower receptacles of the European pear (Pyrus communis L.) ‘La France’ and its giant fruit sport using next-generation sequencing technology. J. Hortic. Sci. Biotechnol. 2014, 89, 293–300. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, G.; Li, Y.; Lyu, M.; Zhang, H.; Zhang, N.; Chen, R. Integrative genomic and transcriptomic analyses of a bud sport mutant ‘Jinzao Wuhe’ with the phenotype of large berries in grapevines. PeerJ 2023, 11, e14617. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Girona, E.; Zhang, Y.; Eduardo, I.; Mora, J.R.H.; Alexiou, K.G.; Arus, P.; Aranzana, M.J. A deletion affecting an LRR-RLK gene co-segregates with the fruit flat shape trait in peach. Sci. Rep.-UK 2017, 7, 6714. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Liu, X.; Gao, H.; Xiao, W.; Chen, X.; Fu, X.; Li, L.; Li, D.; Gao, D. Comparison Between Flat and Round Peaches, Genomic Evidences of Heterozygosity Events. Front. Plant Sci. 2019, 10, 592. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Liu, Q.; Xiong, J.J.; Deng, X.X. Difference of a citrus late-ripening mutant (Citrus sinensis) from its parental line in sugar and acid metabolism at the fruit ripening stage. Sci. China Ser. C-Life Sci. 2007, 50, 511–517. [Google Scholar] [CrossRef]
- Wang, A.D.; Tan, D.M.; Tatsuki, M.; Kasai, A.; Li, T.Z.; Saito, H.; Harada, T. Molecular mechanism of distinct ripening profiles in ‘Fuji’ apple fruit and its early maturing sports. Postharvest. Biol. Technol. 2009, 52, 38–43. [Google Scholar] [CrossRef]
- Ban, S.; El-Sharkawy, I.; Zhao, J.; Fei, Z.; Xu, K. An apple somatic mutation of delayed fruit maturation date is primarily caused by a retrotransposon insertion-associated large deletion. Plant J. 2022, 111, 1609–1625. [Google Scholar] [CrossRef]
- Zhang, M.; Du, T.T.; Yin, Y.R.; Cao, H.Y.; Song, Z.H.; Ye, M.; Liu, Y.T.; Shen, Y.H.; Zhang, L.B.; Yang, Q.; et al. Synergistic effects of plant hormones on spontaneous late-ripening mutant of ‘Jinghong’ peach detected by transcriptome analysis. Food Qual. Saf. 2022, 6. [Google Scholar] [CrossRef]
- Du, X.; Wang, Y.; Liu, M.; Liu, X.; Jiang, Z.; Zhao, L.; Tang, Y.; Sun, Y.; Zhang, X.; Liu, D.; et al. The assessment of epigenetic diversity, differentiation, and structure in the ‘Fuji’ mutation line implicates roles of epigenetic modification in the occurrence of different mutant groups as well as spontaneous mutants. PLoS ONE 2020, 15, e0235073. [Google Scholar] [CrossRef]
- Dong, Q.L.; Yan, Z.Y.; Liu, Z.; Yao, Y.X. Early ripening events caused by bud mutation in Beni Shogun apple. Russ. J. Plant Physl+ 2011, 58, 439–447. [Google Scholar] [CrossRef]
- Cho, H.J.; Kim, G.H.; Choi, C. Differential gene expression and epigenetic analyses between striped and blushed skinned sports of ‘Fuji’ apple. Sci. Hortic.-Amsterdam. 2020, 261, 108944. [Google Scholar] [CrossRef]
- Hubbard, M.; Kelly, J.; Rajapakse, S.; Abbott, A.; Ballard, R. Restriction-Fragment-Length-Polymorphisms in Rose and Their Use for Cultivar Identification. Hortscience 1992, 27, 172–173. [Google Scholar] [CrossRef] [Green Version]
- Aranzana, M.J.; Carbo, J.; Arus, P. Microsatellite variability in peach [Prunus persica (L.) Batsch]: Cultivar identification, marker mutation, pedigree inferences and population structure. Appl. Genet. 2003, 106, 1341–1352. [Google Scholar] [CrossRef]
- Oliveira, E.J.D.; Costa, J.L.; Santos, L.F.D.; Carvalho, F.M.d.; Silva, A.d.S.; Dantas, J.L.L. Molecular characterization of papaya genotypes using AFLP markers. Rev. Bras. Frutic. 2011, 33, 849–858. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, G.H.; Kwon, S.I.; Kim, J.H.; Kwon, Y.S.; Choi, C. Analysis of ‘Fuji’ apple somatic variants from next-generation sequencing. Genet. Mol. Res. 2016, 15, gmr.15038185. [Google Scholar] [CrossRef]
- Fernandez, I.M.A.; Saski, C.A.; Manganaris, G.A.; Gasic, K.; Crisosto, C.H. Genomic Sequencing of Japanese Plum (Prunus salicina Lindl.) Mutants Provides a New Model for Rosaceae Fruit Ripening Studies. Front. Plant Sci. 2018, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.Z.; Cao, Y.H.; Cheng, J.H.; Xiang, J.; Shen, B.W.; Wu, J. Comparative transcriptome analyses of a table grape ‘Summer Black’ and its early-ripening mutant ‘Tiangong Moyu’ identify candidate genes potentially involved in berry development and ripening. J. Plant Interact. 2020, 15, 213–222. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlani, S.; Masiero, S.; Mizzotti, C. Fruit ripening: The role of hormones, cell wall modifications, and their relationship with pathogens. J. Exp. Bot. 2019, 70, 2993–3006. [Google Scholar] [CrossRef]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. Plant J. 2021, 105, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Stella, S.; Van de Weg, W.E.; Guerra, W.; Cecchinel, M.; Dalla Via, J.; Koller, B.; Sansavini, S. Role of the genes Md-ACO1 and Md-ACS1 in ethylene production and shelf life of apple (Malus domestica Borkh). Euphytica 2005, 141, 181–190. [Google Scholar] [CrossRef]
- Yang, X.T.; Song, J.; Campbell-Palmer, L.; Fillmore, S.; Zhang, Z.Q. Effect of ethylene and 1-MCP on expression of genes involved in ethylene biosynthesis and perception during ripening of apple fruit. Postharvest. Biol. Technol. 2013, 78, 55–66. [Google Scholar] [CrossRef]
- Busatto, N.; Tadiello, A.; Trainotti, L.; Costa, F. Climacteric ripening of apple fruit is regulated by transcriptional circuits stimulated by cross-talks between ethylene and auxin. Plant Signal. Behav. 2017, 12, e1268312. [Google Scholar] [CrossRef] [Green Version]
- An, J.P.; Wang, X.F.; Li, Y.Y.; Song, L.Q.; Zhao, L.L.; You, C.X.; Hao, Y.J. EIN3-LIKE1, MYB1, and ETHYLENE RESPONSE FACTOR3 Act in a Regulatory Loop That Synergistically Modulates Ethylene Biosynthesis and Anthocyanin Accumulation. Plant Physiol. 2018, 178, 808–823. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.X.; Liu, D.; Pan, Y.; Gong, W.; Ma, L.G.; Luo, J.C.; Deng, X.W.; Zhu, Y.X. An annotation update via cDNA sequence analysis and comprehensive profiling of developmental, hormonal or environmental responsiveness of the Arabidopsis AP2/EREBP transcription factor gene family. Plant Mol. Biol. 2005, 59, 853–868. [Google Scholar] [CrossRef]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Ireland, H.S.; Yao, J.L.; Tomes, S.; Sutherland, P.W.; Nieuwenhuizen, N.; Gunaseelan, K.; Winz, R.A.; David, K.M.; Schaffer, R.J. Apple SEPALLATA1/2-like genes control fruit flesh development and ripening. Plant J. 2013, 73, 1044–1056. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, R.J.; Ireland, H.S.; Ross, J.J.; Ling, T.J.; David, K.M. SEPALLATA1/2-suppressedmature apples have lowethylene, high auxin and reduced transcription of ripening-related genes. Aob. Plants 2013, 5, pls047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Li, M.; Liu, B.; Yan, M.; Yu, X.; Zi, H.; Liu, R.; Yamamuro, C. Interlinked regulatory loops of ABA catabolism and biosynthesis coordinate fruit growth and ripening in woodland strawberry. Proc. Natl. Acad. Sci. USA 2018, 115, E11542–E11550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlani, S.; Mizzotti, C.; Masiero, S. The NAC side of the fruit: Tuning of fruit development and maturation. BMC Plant Biol. 2021, 21, 238. [Google Scholar] [CrossRef]
- Wang, B.; Hu, W.; Fang, Y.; Feng, X.; Fang, J.; Zou, T.; Zheng, S.; Ming, R.; Zhang, J. Comparative Analysis of the MADS-Box Genes Revealed Their Potential Functions for Flower and Fruit Development in Longan (Dimocarpus longan). Front. Plant Sci. 2021, 12, 813798. [Google Scholar] [CrossRef]
- Dong, T.; Chen, G.; Tian, S.; Xie, Q.; Yin, W.; Zhang, Y.; Hu, Z. A non-climacteric fruit gene CaMADS-RIN regulates fruit ripening and ethylene biosynthesis in climacteric fruit. PLoS ONE 2014, 9, e95559. [Google Scholar] [CrossRef]
- Blanpied GD, S.K. Predicting harvest date windows for apples. In Information Bulletin 221, Cornell Cooperative Extension; Cornell University: Ithaca, NY, USA, 1992. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Singh, V.; Weksler, A.; Friedman, H. Different Preclimacteric Events in Apple Cultivars with Modified Ripening Physiology. Front. Plant Sci. 2017, 8, 1502. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Lee, J.; Rudell, D.; Evans, K.; Zhu, Y.M. Transcriptional Regulation of Auxin Metabolism and Ethylene Biosynthesis Activation During Apple (Malus x domestica) Fruit Maturation. J. Plant Growth Regul. 2016, 35, 655–666. [Google Scholar] [CrossRef]
- Yue, P.; Lu, Q.; Liu, Z.; Lv, T.; Li, X.; Bu, H.; Liu, W.; Xu, Y.; Yuan, H.; Wang, A. Auxin-activated MdARF5 induces the expression of ethylene biosynthetic genes to initiate apple fruit ripening. New Phytol. 2020, 226, 1781–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Ch. | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Description |
|---|---|---|---|---|---|
| Ethylene synthesis and signal transduction | |||||
| MdACO1 | MDP0000195885 | 10 | CAGAATGTCGATAGCCTCGT | GCAGTCCAGAATACAGCTTC | 1-aminocyclopropane-1-carboxylate oxidase 1 |
| MdACO7 | MDP0000200896 | 15 | GGTGGAAAGGGTCCTTCTGT | CACCAGCATCAGTGTGCTCT | 1-aminocyclopropane-1-carboxylate oxidase 1 |
| MdCTR1 | MDP0000230308 | 12 | ACAAGATTTTCATGCCGAAC | TATGGACAAGTTTGGAGGCT | Serine/threonine-protein kinase CTR1 |
| MdEIL1 | MDP0000423881 | 15 | GTTCGATGCTTCGGGACTT | ACCTGACTGGTTCACTGGTTG | ETHYLENE INSENSITIVE 3-like 1 protein |
| MdERF1 | MDP0000128979 | 13 | TCAGATCTTGACACCATCTCT | CACTTGTCACTACTTTGGTGATAG | Ethylene-responsive transcription factor RAP2-3-like |
| Auxin signal transduction | |||||
| MdARF1 | MDP0000194603 | 7 | AAAGATTGGTTGCTGGTGAC | TCACTGACGAGGGTATCTGA | ADP-ribosylation factor 1 |
| MdIAA11 | MDP0000164095 | 9 | TTCGTAAGTGCAGTTCCTCC | TCCATCTCAGTGGCCATATCT | Auxin-responsive protein IAA26-like |
| Transcription factors | |||||
| MdNAC5 | MDP0000868419 | 3 | CATGCAGTTCTGGGGTCACT | TCAAGCGCTAAATGATACGTGC | NAC domain-containing protein 18 |
| MdNAC3 | MDP0000173636 | 5 | GACCACTAGGAGATGGGGTT | ACGTTACCCGTATATCGTTGCT | NAC transcription factor 25-like |
| MdMADS8 | MDP0000366022 | 17 | GCAAAGGAACTTGAGAGCAGC | AATGGACCCAAGTCCTCACC | Developmental protein SEPALLATA 1 |
| MdMADS7 | MDP0000326390 | 14 | AACCTACCAGCCAACGAGAC | CCTTTGTGTTCAGGTGGGAC | MADS-box protein CMB1-like |
| Housekeeping gene | |||||
| MdACTIN | MDP0000170174 | 7 | GGCTCTATTCCAACCATCCA | TAGAAGCAGTGCCACCACAC | Actin-related protein 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Ban, S.; Cho, H.J.; Han, A.R.; Choi, C. Comparative Analysis of Gene Expression between Early Maturation Mutant ‘Beni Shogun’ and ‘Fuji’ Cultivars during Fruit Development and Ripening. Horticulturae 2023, 9, 430. https://doi.org/10.3390/horticulturae9040430
Kim YJ, Ban S, Cho HJ, Han AR, Choi C. Comparative Analysis of Gene Expression between Early Maturation Mutant ‘Beni Shogun’ and ‘Fuji’ Cultivars during Fruit Development and Ripening. Horticulturae. 2023; 9(4):430. https://doi.org/10.3390/horticulturae9040430
Chicago/Turabian StyleKim, Yu Jin, Seunghyun Ban, Hye Jeong Cho, A Reum Han, and Cheol Choi. 2023. "Comparative Analysis of Gene Expression between Early Maturation Mutant ‘Beni Shogun’ and ‘Fuji’ Cultivars during Fruit Development and Ripening" Horticulturae 9, no. 4: 430. https://doi.org/10.3390/horticulturae9040430
APA StyleKim, Y. J., Ban, S., Cho, H. J., Han, A. R., & Choi, C. (2023). Comparative Analysis of Gene Expression between Early Maturation Mutant ‘Beni Shogun’ and ‘Fuji’ Cultivars during Fruit Development and Ripening. Horticulturae, 9(4), 430. https://doi.org/10.3390/horticulturae9040430