
Correlation between Destructive and Non-Destructive Measurements of Highbush Blueberry (Vaccinium corymbosum L.) Fruit during Maturation

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Material
2.2. Chlorophyll Absorbance Measurements (IAD) Using DA-Meter
2.3. Maturity Measurements
2.4. Extraction of Sugars, Organic Acids and Phenolics
2.5. Analytical Methods
2.6. RNA Extraction and Analysis
2.7. Quantitative Reverse Transcription PCR (RT-qPCR)
2.8. Statistics
3. Results
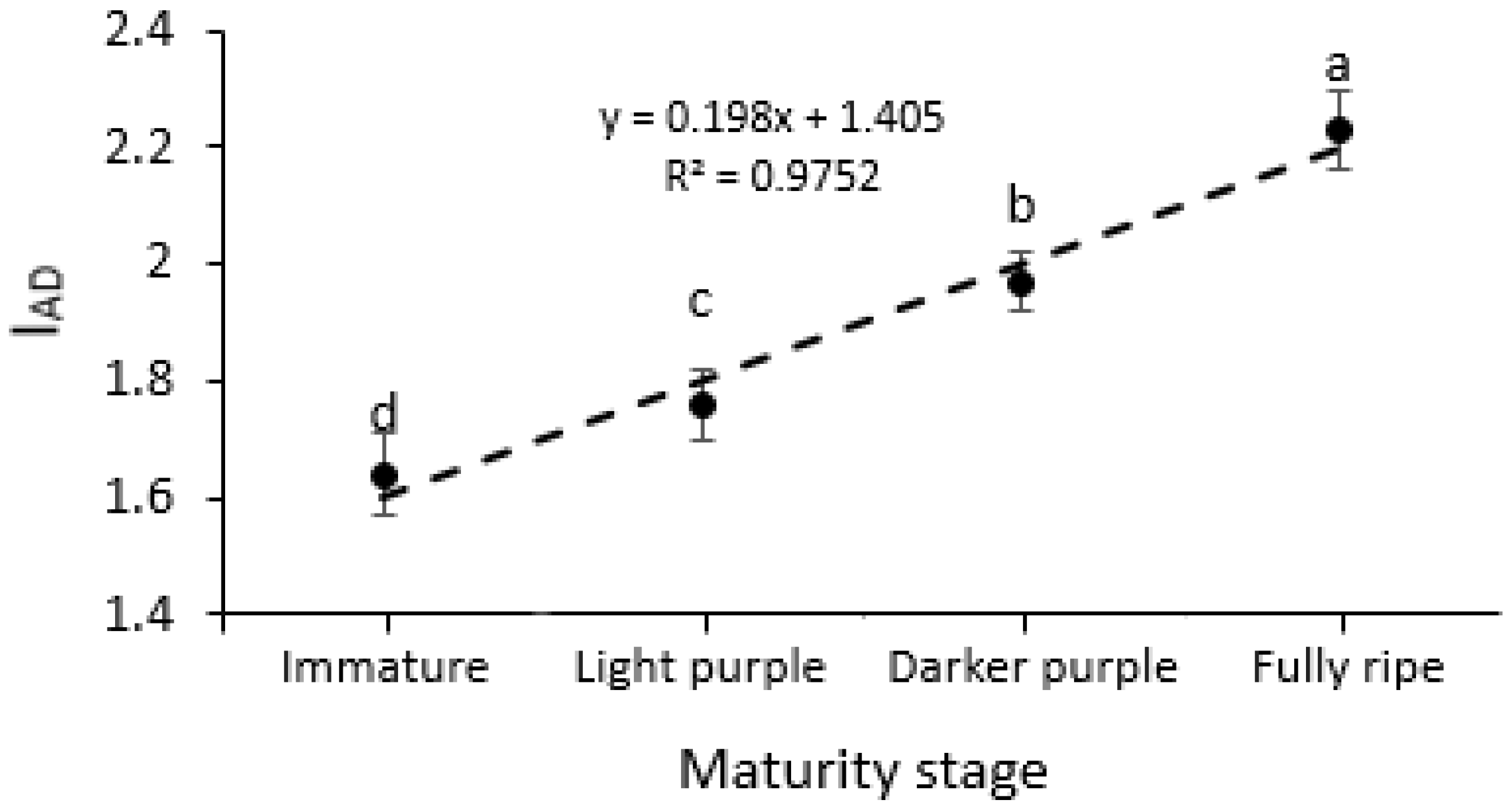
3.1. Chlorophyll Absorbance (IAD) Measurements
3.2. Peel Color and Fruit Firmness
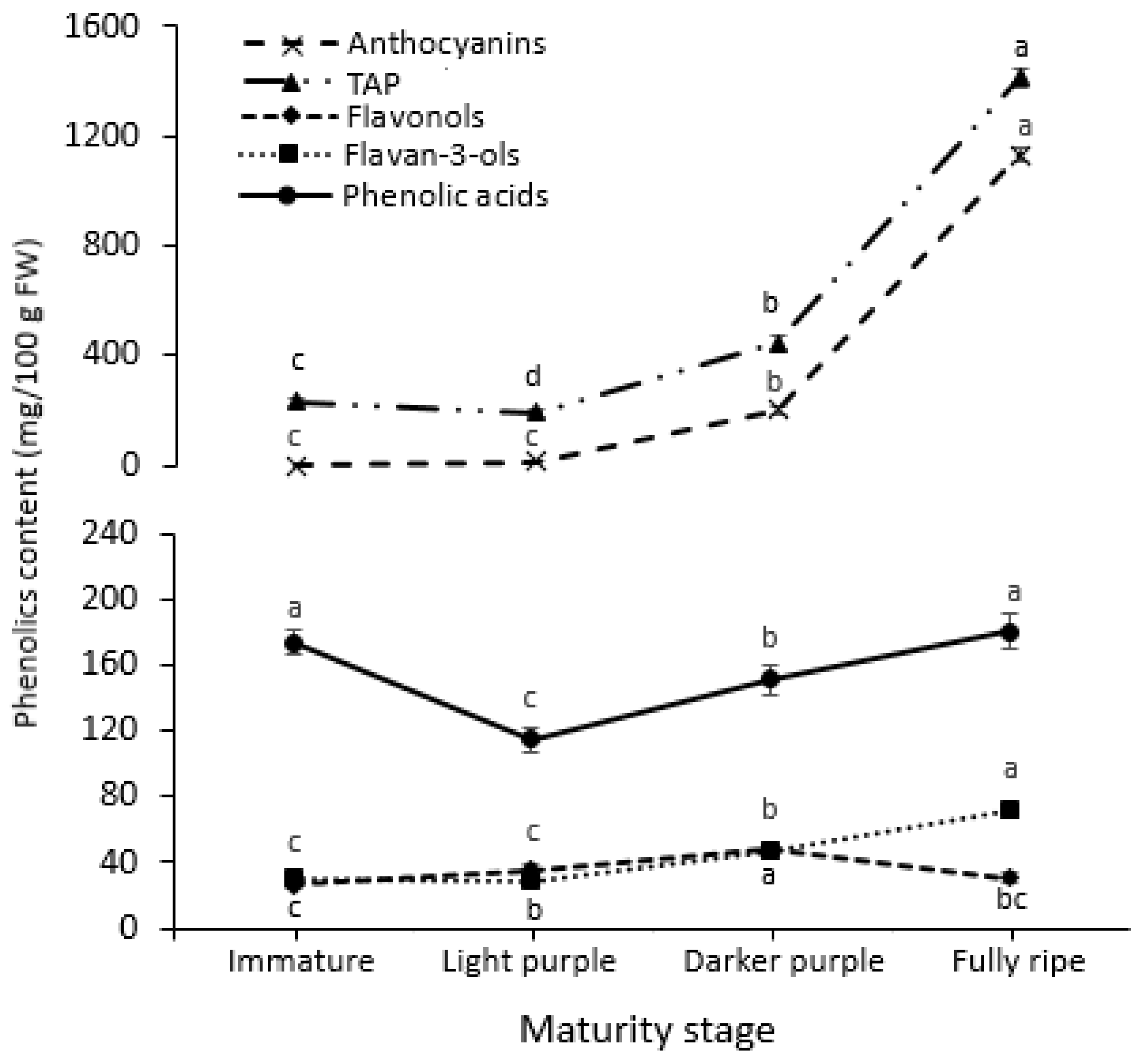
3.3. Fruit Chemical Composition
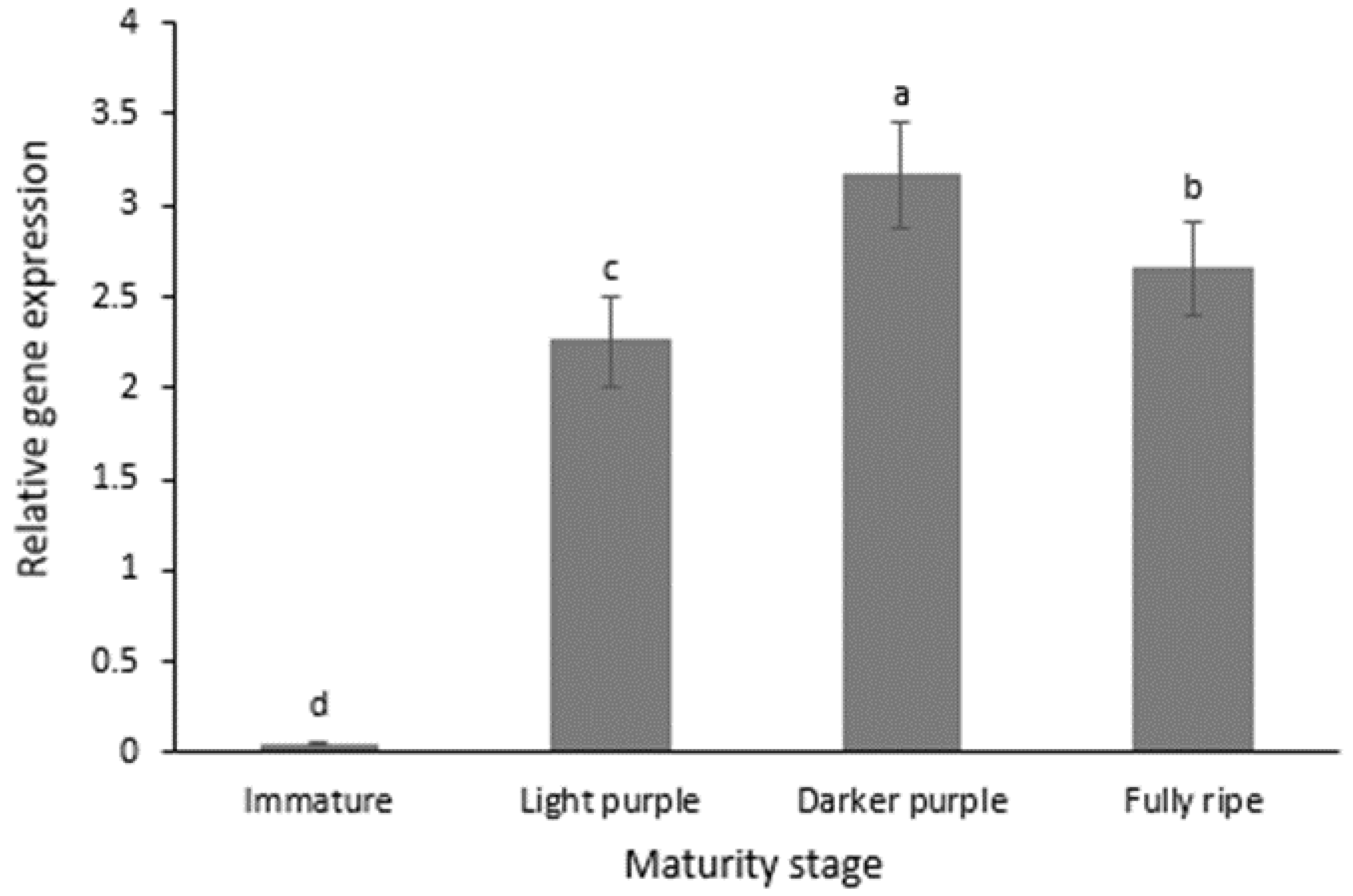
3.4. Relative Expression of Anthocyanidin Synthase (ANS)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Retamales, J.B.; Hancock, J.F. Blueberries, 2nd ed.; Cabi: Boston, MA, USA, 2018. [Google Scholar]
- Grace, M.H.; Xiong, J.; Esposito, D.; Ehlenfeldt, M.; Lila, M.A. Simultaneous LC-MS Quantification of Anthocyanins and Non-Anthocyanin Phenolics from Blueberries with Widely Divergent Profiles and Biological Activities. Food Chem. 2019, 277, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Y.; Zhang, H.C.; Liu, W.X.; Li, C.Y. Survey of Antioxidant Capacity and Phenolic Composition of Blueberry, Blackberry, and Strawberry in Nanjing. J. Zhejiang Univ. Sci. B 2012, 13, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Riihinen, K.; Jaakola, L.; Kärenlampi, S.; Hohtola, A. Organ-Specific Distribution of Phenolic Compounds in Bilberry (Vaccinium Myrtillus) and “northblue” Blueberry (Vaccinium Corymbosum x V. Angustifolium). Food Chem. 2008, 110, 156–160. [Google Scholar] [CrossRef]
- Castrejón, A.D.R.; Eichholz, I.; Rohn, S.; Kroh, L.W.; Huyskens-Keil, S. Phenolic Profile and Antioxidant Activity of Highbush Blueberry (Vaccinium Corymbosum L.) during Fruit Maturation and Ripening. Food Chem. 2008, 109, 564–572. [Google Scholar] [CrossRef]
- Milivojević, J.; Radivojević, D.; Nikolić, M.; Maksimović, J.D. Changes in Fruit Quality of Highbush Blueberries (Vaccinium Corymbosum) during the Ripening Season. Acta Hortic. 2016, 1139, 657–664. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Sun, J.; Jackson, A. Dynamic Changes of Enzymes Involved in Sugar and Organic Acid Level Modification during Blueberry Fruit Maturation. Food Chem. 2020, 309, 125617. [Google Scholar] [CrossRef]
- Woodruff, R.E. Chemical and physical changes of blueberry fruit associated with ripening and detrioration. 1960. Michigan State University of Agriculture and Applied Science. Department of Horticulture. Available online: https://scholar.google.com/scholar?hl=sl&as_sdt=0%2C5&q=chemical+and+physical+changes+of+blueberry+fruit&btnG= (accessed on 15 January 2022).
- Howard, L.R.; Clark, J.R.; Brownmiller, C. Antioxidant Capacity and Phenolic Content in Blueberries as Affected by Genotype and Growing Season. J. Sci. Food Agric. 2003, 83, 1238–1247. [Google Scholar] [CrossRef]
- Zorenc, Z.; Veberic, R.; Stampar, F.; Koron, D.; Mikulic-Petkovsek, M. Changes in Berry Quality of Northern Highbush Blueberry (Vaccinium Corymbosum L.) during the Harvest Season. Turkish J. Agric. For. 2016, 40, 855–864. [Google Scholar] [CrossRef]
- Infante, R. Harvest Maturity Indicators in the Stone Fruit Industry. Stewart Postharvest Rev. 2012, 8, 1–6. [Google Scholar] [CrossRef]
- Ribera-Fonseca, A.; Noferini, M.; Rombolá, A.D. Non-Destructive Assessment of Highbush Blueberry Fruit Maturity Parameters and Anthocyanins by Using a Visible/ near Infrared (Vis/NIR) Spectroscopy Device: A Preliminary Approach. J. Soil Sci. Plant Nutr. 2016, 16, 174–186. [Google Scholar] [CrossRef]
- Smrke, T.; Veberic, R.; Hudina, M.; Stamic, D.; Jakopic, J. Comparison of Highbush Blueberry (Vaccinium Corymbosum L.) under Ridge and Pot Production. Agriculture 2021, 11, 929. [Google Scholar] [CrossRef]
- Smrke, T.; Veberic, R.; Hudina, M.; Zitko, V.; Ferlan, M.; Jakopic, J. Fruit Quality and Yield of Three Highbush Blueberry (Vaccinium Corymbosum L.) Cultivars Grown in Two Planting Systems under Different Protected Environments. Horticulturae 2021, 7, 591. [Google Scholar] [CrossRef]
- Chung, S.W.; Yu, D.J.; Lee, H.J. Changes in Anthocyanidin and Anthocyanin Pigments in Highbush Blueberry (Vaccinium Corymbosum Cv. Bluecrop) Fruits during Ripening. Hortic. Environ. Biotechnol. 2016, 57, 424–430. [Google Scholar] [CrossRef]
- Carbone, F.; Preuss, A.; De Vos, R.C.H.; D’Amico, E.; Perrotta, G.; Bovy, A.G.; Martens, S.; Rosati, C. Developmental, Genetic and Environmental Factors Affect the Expression of Flavonoid Genes, Enzymes and Metabolites in Strawberry Fruits. Plant, Cell Environ. 2009, 32, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New Insights into the Regulation of Anthocyanin Biosynthesis in Fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Liu, J.; Osbourn, A.; Ma, P. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef]
- Yang, J.; Li, B.; Shi, W.; Gong, Z.; Chen, L.; Hou, Z. Transcriptional Activation of Anthocyanin Biosynthesis in Developing Fruit of Blueberries (Vaccinium Corymbosum L.) by Preharvest and Postharvest UV Irradiation. J. Agric. Food Chem. 2018, 66, 10931–10942. [Google Scholar] [CrossRef]
- Zorenc, Z.; Veberic, R.; Slatnar, A.; Koron, D.; Miosic, S.; Chen, M.H.; Haselmair-Gosch, C.; Halbwirth, H.; Mikulic-Petkovsek, M. A Wild ‘Albino’ Bilberry (Vaccinium Myrtillus L.) from Slovenia Shows Three Bottlenecks in the Anthocyanin Pathway and Significant Differences in the Expression of Several Regulatory Genes Compared to the Common Blue Berry Type. PLoS ONE 2017, 12, 1–14. [Google Scholar] [CrossRef]
- Leiva-Valenzuela, G.A.; Lu, R.; Aguilera, J.M. Prediction of Firmness and Soluble Solids Content of Blueberries Using Hyperspectral Reflectance Imaging. J. Food Eng. 2013, 115, 91–98. [Google Scholar] [CrossRef]
- Infante, R.; Contador, L.; Rubio, P.; Mesa, K.; Meneses, C. Non-Destructive Monitoring of Flesh Softening in the Black-Skinned Japanese Plums “Angeleno” and “Autumn Beaut” on-Tree and Postharvest. Postharvest Biol. Technol. 2011, 61, 35–40. [Google Scholar] [CrossRef]
- Ziosi, V.; Noferini, M.; Fiori, G.; Tadiello, A.; Trainotti, L.; Casadoro, G.; Costa, G. A New Index Based on Vis Spectroscopy to Characterize the Progression of Ripening in Peach Fruit. Postharvest Biol. Technol. 2008, 49, 319–329. [Google Scholar] [CrossRef]
- Bonora, E.; Noferini, M.; Vidoni, S.; Costa, G. Modeling Fruit Ripening for Improving Peach Homogeneity in Planta. Sci. Hortic. 2013, 159, 166–171. [Google Scholar] [CrossRef]
- Bonora, E.; Stefanelli, D.; Costa, G. Nectarine Fruit Ripening and Quality Assessed Using the Index of Absorbance Difference (IAD). Int. J. Agron. 2013, 2013. [Google Scholar] [CrossRef]
- Calzarano, F.; Di Marco, S. Further Evidence That Calcium, Magnesium and Seaweed Mixtures Reduce Grapevine Leaf Stripe Symptoms and Increase Grape Yields. Phytopathol. Mediterr. 2018, 57, 459–471. [Google Scholar] [CrossRef]
- Ribera-Fonseca, A.; Noferini, M.; Jorquera-Fontena, E.; Rombolà, A.D. Assessment of Technological Maturity Parameters and Anthocyanins in Berries of Cv. Sangiovese (Vitis Vinifera L.) by a Portable Vis/NIR Device. Sci. Hortic. 2016, 209, 229–235. [Google Scholar] [CrossRef]
- DeLong, J.; Prange, R.; Harrison, P.; Nichols, D.; Wright, H. Determination of Optimal Harvest Boundaries for HoneycrispTM Fruit Using a New Chlorophyll Meter. Can. J. Plant Sci. 2014, 94, 361–369. [Google Scholar] [CrossRef]
- Nyasordzi, J.; Friedman, H.; Schmilovitch, Z.; Ignat, T.; Weksler, A.; Rot, I.; Lurie, S. Utilizing the IAD Index to Determine Internal Quality Attributes of Apples at Harvest and after Storage. Postharvest Biol. Technol. 2013, 77, 80–86. [Google Scholar] [CrossRef]
- Peifer, L.; Ottnad, S.; Kunz, A.; Damerow, L.; Blanke, M. Effect of Non-Chemical Crop Load Regulation on Apple Fruit Quality, Assessed by the DA-Meter. Sci. Hortic. 2018, 233, 526–531. [Google Scholar] [CrossRef]
- Nagpala, E.G.L.; Noferini, M.; Farneti, B.; Piccinini, L.; Costa, G. Cherry-Meter: An Innovative Non-Destructive Device for Cherry Fruit Ripening and Quality Assessment. In Proceedings of the VII International Cherry Symposium, Plasencia, Spain, 23–27 June 2013; pp. 23–27. [Google Scholar]
- Gudenschwager, O.; González-Agüero, M.; Defilippi, B.G. A General Method for High-Quality RNA Isolation from Metabolite-Rich Fruits. S. Afr. J. Bot. 2012, 83, 186–192. [Google Scholar] [CrossRef]
- Harb, J.; Saleh, O.; Kittemann, D.; Neuwald, D.A.; Hoffmann, T.; Reski, R.; Schwab, W. Changes in Polyphenols and Expression Levels of Related Genes in “Duke” Blueberries Stored under High CO2 Levels. J. Agric. Food Chem. 2014, 62, 7460–7467. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing 2021. Available online: https://www.r-project.org (accessed on 15 January 2023).
- Bonora, E.; Noferini, M.; Stefanelli, D.; Costa, G. A New Simple Modeling Approach for the Early Prediction of Harvest Date and Yield in Nectarines. Sci. Hortic. 2014, 172, 1–9. [Google Scholar] [CrossRef]
- Curtis, P.J.; Van Der Velpen, V.; Berends, L.; Jennings, A.; Feelisch, M.; Umpleby, A.M.; Evans, M.; Fernandez, B.O.; Meiss, M.S.; Minnion, M.; et al. Blueberries Improve Biomarkers of Cardiometabolic Function in Participants with Metabolic Syndrome-Results from a 6-Month, Double-Blind, Randomized Controlled Trial. Am. J. Clin. Nutr. 2019, 109, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Jennings, A.; Welch, A.A.; Spector, T.; Macgregor, A.; Cassidy, A. Intakes of Anthocyanins and Flavones Are Associated with Biomarkers of Insulin Resistance and Inflammation in Women. J. Nutr. 2014, 144, 202–208. [Google Scholar] [CrossRef]
- Li, X.; Jin, L.; Pan, X.; Yang, L.; Guo, W. Proteins Expression and Metabolite Profile Insight into Phenolic Biosynthesis during Highbush Blueberry Fruit Maturation. Food Chem. 2019, 290, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Ribera, A.E.; Reyes-Díaz, M.; Alberdi, M.; Zuñiga, G.E.; Mora, M.L. Antioxidant Compounds in Skin and Pulp of Fruits Change among Genotypes and Maturity Stages in Highbush Blueberry (Vaccinium Corymbosum L.) Grown in Southern Chile. J. Soil Sci. Plant Nutr. 2010, 10, 509–536. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Törrönen, R.; Kärenlampi, S.; Hohtola, A. Expression of Genes Involved in Anthocyanin Biosynthesis in Relation to Anthocyanin, Proanthocyanidin, and Flavonol Levels during Bilberry Fruit Development. Plant Physiol. 2002, 130, 729–739. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maturity Stage | IAD |
|---|---|
| Immature | 1.51–1.70 |
| Light purple | 1.71–1.90 |
| Darker purple | 1.91–2.10 |
| Fully ripe | 2.11–2.30 |
| Immature | Light Purple | Darker Purple | Fully Ripe | Sign. | |
|---|---|---|---|---|---|
| L* (lightness) | 58.03 ± 1.52 a | 38.76 ± 1.44 b | 35.50 ± 1.23 c | 32.45 ± 0.98 d | *** |
| C* (chroma) | 34.10 ± 1.23 a | 16.03 ± 1.91 b | 6.73 ± 0.79 c | 3.49 ± 0.34 d | *** |
| h° (hue angle) | 94.94 ± 1.59 c | 14.31 ± 2.13 d | 350.5 ± 2.95 a | 262.1 ± 3.99 b | *** |
| Firmness (N) | 2.13 ± 0.26 a | 0.47 ± 0.09 b | 0.42 ± 0.07 b | 0.23 ± 0.04 c | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smrke, T.; Stajner, N.; Cesar, T.; Veberic, R.; Hudina, M.; Jakopic, J. Correlation between Destructive and Non-Destructive Measurements of Highbush Blueberry (Vaccinium corymbosum L.) Fruit during Maturation. Horticulturae 2023, 9, 501. https://doi.org/10.3390/horticulturae9040501
Smrke T, Stajner N, Cesar T, Veberic R, Hudina M, Jakopic J. Correlation between Destructive and Non-Destructive Measurements of Highbush Blueberry (Vaccinium corymbosum L.) Fruit during Maturation. Horticulturae. 2023; 9(4):501. https://doi.org/10.3390/horticulturae9040501
Chicago/Turabian StyleSmrke, Tina, Natasa Stajner, Tjasa Cesar, Robert Veberic, Metka Hudina, and Jerneja Jakopic. 2023. "Correlation between Destructive and Non-Destructive Measurements of Highbush Blueberry (Vaccinium corymbosum L.) Fruit during Maturation" Horticulturae 9, no. 4: 501. https://doi.org/10.3390/horticulturae9040501
APA StyleSmrke, T., Stajner, N., Cesar, T., Veberic, R., Hudina, M., & Jakopic, J. (2023). Correlation between Destructive and Non-Destructive Measurements of Highbush Blueberry (Vaccinium corymbosum L.) Fruit during Maturation. Horticulturae, 9(4), 501. https://doi.org/10.3390/horticulturae9040501