Abstract
Paraphalaenopsis labukensis Shim, A. Lamb & C.L. Chan (P. labukensis) is an endangered monopodial epiphytic orchid threatened due to habitat fragmentation and overharvesting. Consequently, this research aimed to achieve in vitro propagation of P. labukensis through asymbiotic seed germination and seedling development. This study focused on identification and optimizing capsule maturity, basal media and nutrient requirements. In addition, after hand pollinating the plant, their capsules’ flowers, length, girth, and colour variations were recorded weekly to ascertain morphological maturity characters of this species. Murashige and Skoog (MS), Knudson C (KC), and Vaccin and Went (VW) basal media were used to determine seed vegetation at 60, 90, and 120 days. Subsequently, for the proliferation and development of protocorms, the best basal media were those supplemented with different organic additives (coconut water (CW) and banana homogenate (BH)) and plant growth regulators (PGRs). As a result, 120 days after pollination (DAP) was chosen as the best capsule age for germination since the capsules had reached maturity. The highest germination rate was attained after 90 days of culture in Knudson C (KC) medium, with 98.78 ± 0.89%, followed by Murashige and Skoog (MS) medium, with 92.80 ± 3.26%. The KC supplemented with 0.5 mg/L NAA media documented a maximum percentage of 17.25 ± 0.96%. The 5 g/L (w/v) BH recorded a higher protocorm proliferation percentage than CW. Additionally, after 150 days of culture (DOC), this medium combination produced the most leaves, averaging four or five. The result of this present study has successfully established an effective in vitro propagation protocol for this species.
1. Introduction
The Orchidaceae family is one of the most prominent flowering plant families, and its cut flowers and potted plants are economically valuable in the floriculture industry [1]. The highest mountain in the Malay Archipelago, Mount Kinabalu, has been linked to the species richness in Borneo [2]. Borneo is also among one of many orchid biodiversity hotspots, even though the estimated orchid species count in the region varies significantly. Numerous species of high horticultural importance inhabit some parts of Mount Kinabalu, including Paphiopedilum rothschildianum, Paraphalaenopsis labukensis (P. labukensis), and Renanthera bella [3].
P. labukensis is a leafless tropical monopodial epiphytic species only found in restricted locations in the Labuk River Basin, Sabah. The plant is also known as the ‘rat-tailed’ orchid as its leaves, which hang downwards, could grow up to 3 m long [4]. In the wild, the terete leaves of P. labukensis generally do not exceed 120 cm but could reportedly exceed 2 m in cultivation. The plant could reach approximately 6–9 mm thick, and its inflorescence typically bears five or six beautiful cinnamon-scented flowers (Figure S1), which bloom twice a year in February and October.
P. labukensis is a distinctive orchid species as it is one of the four genera endemic to Sabah, Borneo, while the other three (P. denevei, P. serpentilingua, P. laycockii) are found in Kalimantan Timur Barat [5]. Nevertheless, the IUCN classified the orchid as endangered [4,6,7]. P. labukensis is endangered due to its considerable collection by orchid traders in the wild. Moreover, the orchid is frequently poached due to its high public profile and unique leaf characteristics, which could grow up to 3 m long and hang onwards. Most P. labukensis also grow within hill ridges, suspended from small trees, or above ground, making it easily accessible. Large-scale in vitro propagation could reduce the commercial value of the plant, fulfilling commercial demands, and reintroducing the threatened species into the wild, which could be an option for diminishing pressure from illegal collections of the orchids. It is also critical to utilise efficient techniques to increase the number of orchids for restoration.
Typically, orchids are seed propagated. Nonetheless, orchid seeds germinate at a 2–5% rate, while seedlings take 12 years to mature naturally [8]. Furthermore, orchid seeds require particular conditions to germinate in their habitats, which leads to their symbiotic relationship with a mycorrhizal fungus [9]. Consequently, asymbiotic seed germination is a highly effective tool for conservation efforts as it offers a practical option for the fast propagation and development of orchid seedlings. Nevertheless, understanding seed viability is still crucial when determining ideal germination circumstances.
Successful asymbiotic seed productions of several wild orchids have been reported, including Eulophia alta [10], Bletilla striata [11], Cyrtopodium paludicolum [12], (Spathoglottis plicata [13], Cremastra appendiculata (D. Don) Makino [14], and Paphiopedilum tigrinum [15]. Other essential elements influencing the success of in vitro asymbiotic seed cultivations and protocorm development include macro and micro element compositions, vitamins, sucrose, nitrogen, carbohydrates, and organic nutrients [16,17,18]. Several authors reported successful seed germinations by employing various basal mediums supplemented with plant growth regulators (PGRs) [19,20,21,22,23,24]. The beneficial effects of plant growth regulators, such as benzyl aminopurine (BAP) and naphthalene acetic acid (NAA), on the growth and development of orchids have also been widely documented [8,25,26,27]. Moreover, numerous studies also have reported improving the culture medium via organic additives supplementation [28,29].
Benzylaminopurine (BAP) and naphthalene acetic acid (NAA) play a crucial role in regulating various physiological process, including enhanced in cell division and shoot regeneration. These plant hormones have a wide-ranging influence on various plant process, including seed germination, and they remain active throughout all stages of germination processes [1,30,31]. In addition to plant growth regulators, the supplementation of organic additives, such as coconut water (CW) and banana homogenate (BH) contain diverse range of nutrients, such as carbohydrates, vitamins, minerals, amino acids, which support cell growth and development [32,33].
The unrestricted procurement of P. labukensis from its native habitat and damage caused to its indigenous habitation may result in its extinction. Moreover, the micropropagation of P. labukensis has never been documented, hence requiring a methodology for this commercially valuable orchid. The maturity level of the orchid seed has a significant impact on the success of seed germination. Therefore, the current study was conducted to procure a systematic mass P. labukensis cultivation procedure. As a result, this study was carried out to determine the maturity level of the seeds that are adequate for seed germination. The influences of different basal media, coconut water (CW), banana homogenate (BH), and plant growth regulators (PGRs) on P. labukensis seed germination and plantlet growth were also identified.
2. Materials and Methods
2.1. Plant Material
Paraphalaeonopsis labukensis is a long-leaved epiphytic orchid endemic to Sabah, Malaysia, which flowers from October to February. This study obtained P. labukensis capsules through hand pollination at the greenhouse in the Institute for Tropical Biology and Conservation, Universiti Malaysia Sabah (06°003′33″ N, 116°012′29″ E). The capsules were harvested 60, 90, and 120 days after pollination (DAP) and were preserved in a tiny bottle containing silica gel upon being transferred and stored at 4 °C before cultivated.
2.2. Hand Pollination Technique
The current study employed a hand pollination method according to the procedure reported previously [34] with slight alterations. The pollination was conducted two to three days after all the flowers had completely bloomed.
The first step started with identifying fully open flowers and targeting the columns and anther caps. The process was performed early morning when the flowers were fresher. Next, the anther caps and pollinia were gently removed with a pin from the gynostemium. A slight upward pressure was administered to the bottom of the anther caps, which adhered to the toothpick upon contact. The anther caps were then carefully removed before depositing the pollens to the same or another flower by gently depositing them onto the stigmatic surfaces. The step was repeated a few times to ensure the maximum amount of pollen was transferred. Once hand pollinating was completed, the capsules were labelled with the parent plants and the pollination date. Subsequently, flower senescence, capsule development, and capsule dehiscence were observed.
2.3. Capsule Formation and Development
The growth and maturation of the capsule were monitored on a daily basis, starting from pollination until reaching full maturity, to document any observed changes. Measurements of the capsule length and diameter were conducted and recorded. The length of the capsule was measured using a ruler (Figure S2), while the diameter was measured using a Vernier calliper (white arrow) (Figure S2). To ensure the capsules were harvested at the appropriate stage, mature capsules were collected before natural dehiscence by carefully cutting them from the inflorescence. In the case of P. labukensis, the capsules were considered mature and ready for harvesting 120 days after pollination.
2.4. Surface Sterilisation
The P. labukensis capsules were first brushed with a soft brush under running tap water for the initial sterilisation procedure. The capsules were then immersed in a 30% antifungal fungicide solution (Anchor Thiram) for 15 min before rinsing with running tap water. Subsequently, the capsules were subjected to surface sterilisation post-drying with tissue paper.
Next, the surface sterilisation of the capsule surfaces was performed under a laminar flow to avoid contamination. This reduces the risk of dirt and small particles in open the air. The seed capsules were first immersed in a mixture of 20% sodium hypochlorite (NaOCl) solution and two drops of Tween 20 for 15 min. The capsules were then thoroughly rinsed three times with sterile distilled water. Subsequently, the capsules were dipped in 70% ethanol for 10 s before being passed under a flame rapidly for a few seconds. The capsules were cut into halves (longitudinally) and aseptically cultured on three different basal media, KC, MS, and VW. The cultures were then incubated at 25 ± 2 °C under a 16/8 h light photoperiod.
2.5. Tetrazolium Viability Test (TZ)
A few seeds were stained with 2, 3, 5-triphenyl tetrazolium chloride (TTC) (Thermo Fisher Scientific® United Kingdom (UK), Loughborough, UK) staining test to determine their viability. Around 3 mg seeds were incubated in 1% TTC solution in the dark for 40–48 h at 28 °C and then incubated in a water bath at 40 °C. The procedure employed is followed according to the report previous by Hosomi et al. [35] with slight modifications.
The colour development of intact viable seed embryos was then observed under a stereomicroscope (Motic, Hong Kong, China). Before observation, the seeds were washed thrice with distilled water, and around 200 seeds were observed in five replicates. For the result, pink or dark red seeds were considered viable, while colourless or without embryos were deemed unviable. Viable seed percentages were obtained by dividing the number of viable embryos by the total number of embryos analysed [36].
2.6. Culture Initiation, Seed Germination on Different Basal Media
Three basal media, Murashige and Skoog (MS) (Table S1) [37], Knudson C (KC) (Table S2) [38], and Vacin and Went (VW) (Table S3) [39], were utilised in the preliminary experiments in the present study to assess the optimal seed germination and protocorm formation. The pH of the medium was adjusted according to their respective pH value, such as [KC (pH 5.2–5.3); VW (pH 5.2); and MS (pH 5.6–5.7)] and autoclaved at 121 °C for 20 min. Subsequently, the sterilised media were aliquoted equally (50 mL) in 9 cm diameter Petri dishes. The seed of 150 DAP were added to the dishes. Subsequently, the seeds were monitored weekly up to 150 days of culture with a total of 200 seed per culture to evaluate the developmental stages of the seeds and protocorms. All culture media were incubated at 25 ± 2 °C under continuous 16/8 h of light and dark conditions from a cool white inflorescent. Seed germination percentage were recorded after 30 DOC. Each treatment conducted was performed in five replicates and each replicate consisted of 200 seeds. Resultantly, the KC medium shows the most optimum results and thus employed in the following stages of the study.
Individual seeds were observed with a stereomicroscope (Zeiss) and digital microscope before an image analyser was employed to count and evaluate them. When the testa seeds ruptured due to the enlarging of embryos (stage 2), the seeds were considered germinating. The percentage of seed germination (%) is calculated based on the formula below:
Percentage of seed germination (%) = [total of seed germinated/total seeds (n = 200)] × 100
The seed developmental stage identification was modified and adopted from [10,40]. The results obtained in the current study are summarised in Table 1.

Table 1.
P. labukensis seed development stages.
2.7. Protocorm Proliferation and Development
The KC medium was incorporated with varying CW and BH concentrations, 5, 10, 15, and 20%, to test optimal protocorm proliferation. The CW was procured from young coconuts directly after filtration. Meanwhile, the banana peels (homogenate) were weighed according to the desired amount. The protocorms (1–2 mm long) from 60 days old cultures cultivated from germinated seeds are employed as explants. The growing protocorms were then subcultured every four weeks to boost seedling maturation and development rates.
The shoot and root induction and further growth were performed on KC media supplemented with various NAA (0.5, 1.0, 1.5, and 2 mg/L) and BAP (0.5, 1.0, 1.5, and 2 mg/L) concentrations and combinations of both. All culture media were incubated at 25 ± 2 °C under continuous 16/8 h of light and dark conditions for a cool white inflorescence. All the experiments were performed in five replicates, and 10 cultures were performed for each protocorm evaluation for each treatment. The observation of protocorm proliferate, protocorms forming leaves or roots, the average length of leaves and roots, and percentage of necrosis was done every ten days.
2.8. Statistical Analysis
The experiments were conducted using a completely randomized design (CRD). Each treatment was carried out in five replicates and repeated twice, resulting in a total of 200 to 300 seeds being observed per culture. The germination period and development were monitored every 20 days for up to 150 days of culture. The data were statistically analysed using a one-way analysis of variance (ANOVA) to determine any significant differences between the means. The results are presented as mean values ± standard deviation (SD). To further compare the means, Duncan’s multiple range test was applied at a significance level of p = 0.05 and p < 0.05 for protocorm proliferation and development.
The statistical analysis of the data was performed using the SPSS software (version 20), IBM Corporation, Armonk, New York.
3. Results
3.1. Morphology Indicator for Capsule Maturity
Three days post-bloom, this study’s P. labukensis flower samples were manually pollinated to procure reliable and adequate seed sources for the seed germination stage. Capsule development in length and diameter after 15 days of hand pollination increased. Subsequently, 7–10 days post-pollination, the diameter of the capsules rose from 0.5 to 0.8 ± 0.3 cm. In the following 15 days, the diameter and length of the capsules reached 1.15 ± 0.53 cm and 10 ± 0.5 cm, which remained constant until maturation. On average, the capsules were 4–10 cm on the longest axis and 0.8–1 cm long on the widest axis (Figure 1).
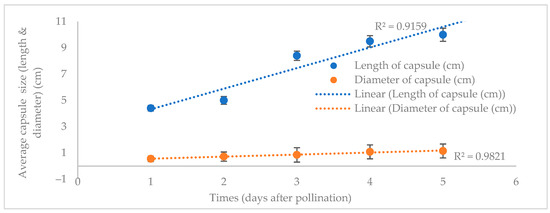
Figure 1.
The change in P. labukensis average capsule length and diameter (cm).
When the P. labukensis flower samples in the current study dried, they were still linked to the tips of their ovaries. The orchid seed capsules were pubescent, upright, and possessed thick, papery walls. The longitudinal sides of the capsules begin to fracture when they fully mature, releasing the seeds inside. The capsules observed changed from dark green to pinkish red five days post-pollination (Figure 2A). The level of development on the capsules was more apparent at 40 DAP when they turned dark crimson of pollination (Figure 2B).

Figure 2.
The capsule development pattern and maturation post-DAP. (A) The swollen immature capsule in diameter (arrow). Scale bar = 2 cm. (B) The immature capsules after 30 DAP. Scale bar = 3 cm. (C) The dark red mature capsules after 120 DAP. Scale bar = 3 cm.
The mature P. labukensis capsules in this study were tight and soft throughout the observation, and at 120 DAP, they dehisced. The capsule gradually changed colour from dark crimson to dark red (Figure 2C). The matured capsules were dry, powdery, dark brown, and were readily distributed on the culture media. The mature P. labukensis capsule seeds were single-spindled with paper-thin seed coats enclosing an extremely small and simplified embryo in globular clusters of cells, while the seeds were oblong or slightly fusiform.
3.2. Seed Viability
In this study, seed viability assessments were conducted to determine the quality of the seeds obtained from three maturity levels; 60, 90, and 120 DAP. The capsules responded differently to the tetrazolium solutions test (Table 2).
Table 2.
The effects of different capsule maturity levels on P. labukensis seed viability.
The 60 DAP capsules demonstrated no response towards the viability test (Figure 3A), while the seeds were non-viable as none of the seeds was stained red. Moreover, the embryos were absent and possessed undeveloped zygotic embryos. The mature P. labukensis seeds at 120 DAP recorded the most viable seeds, where most of the embryos were stained light-rose-red with the highest seed viability at 88.42 ± 2.15 (Figure 3C). Seed analysis using scanned pictures subjected to the tetrazolium test also proved effective, where the viable (light-rose-red), non-viable (white), and empty seed coats (no embryo) were distinguishable.
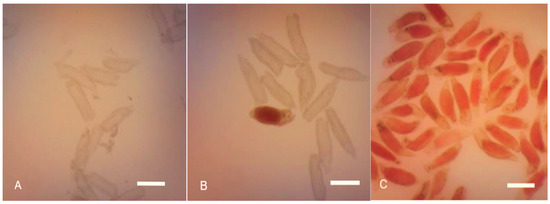
Figure 3.
The effects of 1% tetrazolium chloride solution on P. labukensis seed staining. (A) The stainless embryos from 40 DAP capsule seeds. Scale bar = 0.2 mm. (B) Unstained and red-stained embryos from 90 DAP capsule seeds. Scale bar = 0.2 mm. (C) Red-stained embryos from 120 DAP capsule seeds. Scale bar = 0.2 mm.
3.3. The Effects of Basal Media on In Vitro Seed Germination
Three basal media, MS, KC, and VW, were utilised in the present study for in vitro seed cultivation, which resulted in varied responses (Table 3). Over 70% of the seeds collected at 120 DAP germinated in all media after 50 days of culture (DOC). The KC medium documented the highest seed germination rate within 50 DAC, with mean ± standard deviation (SD) of five replicates of 42.90 ± 3.98% of the seeds growing to stage 3.
Table 3.
The effects of varying basal media on P. labukensis seed germination.
The MS and VW media recorded no discernible differences. After 150 DAC, the seeds cultivated in KC and MS basal media recorded germination rates of over 90%. Nevertheless, only 75.66 ± 11.42% of the seeds in VW germinated. The KC medium produced optimum P. labukensis seed germination compared to the MS and VW media. Consequently, the KC medium was employed in further experiments.
3.4. Observation of Protocorm Developmental Stages
The seed-to-seedling morphological development stages of the P. labukensis in this study were recorded (Figure 4). First, curves on the surfaces of the protocorms were formed at 35 DOC. The curves then developed apical shoots that further grew into new single leaves (Figure 4E). The second leaves were recorded after 43 DOC (Figure 4F). At 60 DOC, the third leaf began to rise between the two leaves, resulting in further development (Figure 4G). The roots of P. labukensis samples observed in the present study started to form after the formation of two leaves, which was at 50 DOC. Some protocorms formed leaves but did not develop roots, whereas the others started to form roots after 50 DOC. Complete plantlets were documented after 150 DOC with complete root systems.

Figure 4.
The P. labukensis protocorm development at stages (A) Stage 0: the ungerminated seed under SEM. Scale bar = 1 mm. (B) Stage 1: a swollen seed that formed a yellow protocorm. Scale bar = 2 mm. (C) Stage 2: the appearance of rhizoids where embryo breaks out from testa coat. Scale bar = 2 mm. (D) Stage 3: the formation of protruding curves on the surface of a protocorm. Scale bar = 2 cm. (E) Stage 4: the apical shoot is present. Scale bar = 3 cm. (F) Stage 5: the second leaf is present. Scale bar = 5 cm. (G) Stage 6: there is the presence of two or three leaves with roots (seedling). Scale bar = 5 cm.
3.5. The Effects of KC Basal Medium Supplemented with Organic Additive on Protocorm Development and Proliferation
In the current study, BH recorded a higher P. labukensis proliferation percentage than CW (Table 4). The 5 g/L (w/v) BH employed in this study documented 15.75 ± 0.96% protocorm proliferation percentage (Figure 5A), which was the highest, while the lowest was observed in the media supplemented with 20% (v/v) CW at 1.00 ± 0.52% (Figure 5C,D). The enhanced protocorm proliferation and development in the media added with BH and CW suggested that some organic substances positively affected the germination and growth of the orchid seed samples in this study. Furthermore, the presence of molecules similar to cytokinin has been linked to the beneficial effects of CW.
Table 4.
The effects of natural additives on P. labukensis protocorm proliferation under 16 h photoperiod at 25 ± 2 °C after 150 days of culture.

Figure 5.
P. labukensis in vitro protocorm proliferation and seedling development in KC medium supplement with (A) 5 g/L (w/v) BH, (B) 5% (v/v) CW, (C,D) 20% (v/v) CW (stage 6), (E) 0.5 mg/L NAA, (F) 2.0 mg/L BAP, (G) 0.5 mg/L NAA (stage 6), (H) 0.5 mg/L BAP + 1.0 mg/L NAA (stage 6), and (I) 1.0 mg/L BAP + 1.0 mg/L NAA (Stage 6) (Scale bar ((A,B) = 1 mm, (C,D) = 4 cm; (E,F) = 1 mm; (G) = 1 cm; (H,I) = 5 cm)).
3.6. The Effect of KC Basal Media Supplementd with Single PGRs on Protocorm Development and Proliferation under 16 h Photoperiod at 25 ± 2 °C after 150 Days of Culture
Most of the treatments employed in this study recorded varied protocorm proliferation percentages (Table 5). The 0.5 mg/L NAA media documented the maximum percentage at 17.25 ± 0.96% (Figure 5E). The fastest response was observed on day 30 after subculture. On the other hand, the media with 2.0 mg/L BAP recorded the lowest protocorm proliferation percentage of 5.00 ± 0.10%. The same media also recorded the highest new protocorm mean at 3.50 ± 0.57 (Figure 5F).
Table 5.
The effects of PGRs on P. labukensis protocorm proliferation under 16 h photoperiod at 25 ± 2 °C after 150 days of culture.
3.7. The Effects of KC Basal Medium Supplemented with Single PGRs on Seedling Formation
Depending on the concentrations, the plant growth regulators utilised in this study elicited varied orchid seed germination and seedling growth rates. The single PGR treatment considerably affected the P. labukensis protocorm growth and development (Table 6). On the other hand, the KC medium supplemented with NAA resulted in a better effect than BAP on the shoot and root formation of the orchid.
Table 6.
The influences of KC medium supplemented with single BAP and NAA on P. labukensis seedling formation after 240 days of culture (DOC).
The 0.5 mg/L NAA recorded the highest leaves produced at 8.00 ± 0.82% (Figure 5G), while 1.5 mg/L NAA recorded the highest shoot induction at 13.25 ± 0.50%. Moreover, the orchids grown in the media with NAA produced five more leaves compared to the seedlings cultivated in the BAP-added media.
The BAP-incorporated media did not produce a better response towards P. labukensis protocorm growth and development. The seeds in the 1.0 mg/L BAP recorded a higher protocorm development response of 8.50 ± 0.96%, whereas both 1.5 and 2.0 mg/L BAP documented 5.00 ± 0.57%, which was the lowest value. On the other hand, the P. labukensis in the control treatment recorded 10.00 ± 0.57% growth.
The highest mean number of leaves, recorded at 08.00 ± 0.82%, was procured with 5 mg/L NAA, with which four or five leaves were observed after 150 DOC. Meanwhile, 2.0 mg/L BAP documented the lowest mean number of leaves. Nonetheless, high concentrations of BAP and NAA (1.5–2.0 mg/L) did not contribute to forming more leaves. Instead, they inhibited the formation of leaves on the P. labukensis specimens. The 2.0 mg/L NAA and BAP treatments recorded 2.00 ± 0.82% and 1.00 ± 0.20% mean number of leaves, respectively.
The lowest concentration of NAA (0.5 mg/L) recorded the highest percentage and mean number of roots (12.65 ± 0.58%). The NAA hormone induced shoot and root formations in the P. labukensis samples employed in the present study. Conversely, the KC medium supplemented with 2.0 mg/L BAP recorded the lowest root formation percentage of 3.00 ± 0.43%.
3.8. The Effects of KC Basal Medium Supplemented with PGRs Combinations on Seedling Formation
The seedlings cultivated in the KC medium with 0.5 mg/L BAP + 1.0 mg/L NAA reached 34.75 ± 0.56% (shoot) and 18.50 ± 0.57 (root) formation in 215 days (Table 7 and Figure 5H). Furthermore, this medium also contributed to the highest mean number of leaves (5.75 ± 0.63) and roots (8.65 ± 0.86). Moreover, in 150 DOC, this medium combination also demonstrated the highest number of leaves produced, averaging four or five.
Table 7.
The influences of KC medium supplemented with combination of BAP and NAA on P. labukensis seedling formation after 240 days of culture (DOC).
The seedlings in the 1.0 mg/L BAP + 1.0 mg/L NAA required 235 days to develop (presence of leave and root), as demonstrated in Figure 5I. The 1.0 mg/L BAP + 1.5 mg/L NAA combination recorded the lowest shoot formation percentage with 10.00 ± 0.25%. Conversely, the 0.5 mg/L BAP + 1.5 mg/L treated NAA medium recorded the lowest mean number of roots. The results supported the findings suggesting that PGR combinations yield much higher growth and development rates of most orchid species. Furthermore, media with combinations of BAP and NAA were generally more effective than employing only NAA or BAP in inducing shoots.
4. Discussion
The hand pollination of orchid capsules can be a delicate and long process. Nonetheless, the technique is essential for producing healthy new generations of orchids. Hand pollinating orchid capsules is common in orchid cultivation, especially for species with limited natural pollination opportunities. The technique reportedly improved successful orchid capsule formation at a 95.47% success rate [41]. Nevertheless, some pollinated orchid flowers did not develop into capsules due to fungal infections or being eaten by predators, such as ants, before maturation [42].
The period for an orchid capsule to reach full maturity varies according to species [43]. Yet, the length and diameter of the capsule seed are not the most important parameters in evaluating seed maturity [44]. Moreover, orchid seeds differ in embryo morphometry, seed coat decoration, size, shape, and volume [45]. Consequently, knowledge of seed germination capabilities is necessary to determine the optimal germination conditions, which then could enhance the seed range that would most likely propagate [46]. The information also distinguishes the proportions of seeds not germinating due to unfavourable conditions from those lacking viability.
A report by Mayo-Mosqueda [47] suggested that the capsule age and maturity influenced orchid seed viability. Embryos would only be viable after the ripening of the capsules [41]. Furthermore, Lo et al. [48] reported that the walls of the carapace of a seed become cutinised and lignified upon maturity. Identifying viable seeds for species propagation and conservation is also crucial for creating a native orchid seed as it could be impacted even in ideal circumstances [49,50,51].
In most orchid seeds, the embryo is undifferentiated and without endosperm. On the other hand, the embryos in new seeds are a collection of living cells that could be dyed with TTC. Consequently, TTC staining viability evaluation is unsuitable for employment on immature seeds [52]. Nevertheless, the assessment has been reported as one of the most attested methods for evaluating seed quality and embryo maturation [35,53].
Tetrazolium is a quick assessment, and thus is crucial, especially when deciding how to maintain seed banks [54]. Dehydrogenases convert the colourless tetrazolium chloride to a red compound in living tissues, hence colouring viable seeds. The intensity of the tint ranges from pink to dark red [55]. The method has been successfully utilised to evaluate the viability of orchid seeds [56,57]. In this study, the seeds from 120 DAP capsules were chosen for germination assessments. At this stage, the seeds have completed their development and are ready for dispersal and germination under favourable condition. On the other hand, the seeds at 60 DAP showed no response towards the viability test, suggesting that they are still immature and have not yet developed the necessary structured or stored enough resources required for successful germination.
The basal medium could significantly affect epiphytic orchid seed germination, including the prevalent MS, KC, and VW media (Tables S1–S3). Each medium contains different essential nutrients and hormone compositions, which could influence the growth and development of orchid seedlings [58,59,60]. This study utilised three basal media, each with a unique chemical makeup.
The KC medium is a simple medium comprising only inorganic salts and sucrose. The medium possesses lower cytokinin levels than its MS counterpart, which could result in slower shoot growth. Nevertheless, the seeds cultivated in KC media demonstrated improved root growth, resulting in more robust seedlings with higher survival chances post-transplantation to soil or other growth media. Unfortunately, the medium might not be suitable for all orchid species, and some might require additional nutrients and growth regulators commonly used in MS medium for optimal growth.
The MS culture is a popular basal medium for orchid seed germination due to its balanced macronutrient, micronutrient, vitamin combination, and PGRs, including auxins and cytokinins. The high cytokinin levels in the medium can also promote shoot growth, leading to a higher seed germination percentage. Nonetheless, excessive cytokinin levels could result in hyperhydricity, a condition where the leaves of the seedlings become translucent and soft, and might eventually die. The phenomenon could impact some orchid species sensitive to high cytokinin levels, resulting in poor seedling growth [61]. Consequently, modifying nutrients for orchid propagation is essential.
In other report by Stewart, [62] classified orchids that respond to in vitro propagation into two groups based on their basic nutritional requirements. The first category includes orchids that germinate easily in a culture medium with a simple nutrient composition, such as KC and VW. The second group encompasses species that require a culture rich in macro- and micronutrients, such as the MS medium [63].
Organic additives are commonly employed to promote orchid growth. Previous studies have incorporated two types of organic nutrient supplements into the KC basal medium to improve protocorm proliferation, including CW and BH. Reports recorded a high proliferation rate in the medium incorporated with BH. It was also discovered that a medium with Hyponex BH added was the ideal seedling formation culture of P. wardii and P. spicerianum [64]
BH is rich in minerals, such as iron and potassium, vitamins and amino acids that play a critical role in promoting the growth of protocorm-like bodies (PLBs) [1]. The study also documented that the beneficial influences of BH were due to its pH buffering abilities, thus maintaining the pH of the culture medium and further encouraging PLB growth. In a report by Nambiar et al. [65], 10% Emas and Rastali (banana) were the ideal homogenates to replace sucrose in culture media among various banana cultivar homogenates. The study proved that lower BH concentrations increased orchid proliferation, but it was unsuitable for protocorm growth improvement in high concentrations. The PLBs were not healthy when supplemented with BH, especially at higher percentages.
CW is rich in cytokinins, auxins, and other growth-promoting hormones. Consequently, the substance is an ideal medium for orchid tissue culture. The hormones aid in regulating cell division and differentiation, leading to seedling formation [66,67]. Nevertheless, in this study, CW-supplemented KC medium failed to promote high protocorm proliferation; however, the advantages of CW in improving seed germination and development of Dendrobium orchid seeds were consistent with the Cymbidium protocorm growth stimulation in CW-containing media [68]. Research by Van Wikle and Pullman, [69] also demonstrated the abilities of CW to improve Doritaenopsis seedling growth. Furthermore, a modified KC nutrient solution with 10% v/v coconut water, 0.5 mg L−1 niacin, and 0.5 mg/L pyridoxine HCl added formed a compact callus, which then proliferated into Dendrobium fimbriatum PLBs [70].
High concentrations of PGR could lead to browning necrosis in protocorms. The browning phenomenon could be due to unsuitable culture conditions, such as inappropriate nutritional balance or insufficient growth-stimulating substances [71]. Combining BH and CW in tissue culture media could provide a balanced nutrient source and growth regulators to enhance protocorm proliferation and subsequent seedling formation. Nonetheless, the exact composition of the tissue culture media, including the concentrations of BH and CW, might vary depending on the specific orchid species and growth conditions.
The present study employed PGRs, such as BAP and NAA, for protocorm proliferation. The P. labukensis protocorm plantlets observed in the current study responded positively to all parameters evaluated. Another report by Parthibhan et al. [72] reported that the protocorm explants, which are composed of highly meristematic cells, elicited multiple new shoots in the presence of PGRs. PGRs are essential in promoting the growth of numerous plant species [73]. Moreover, protocorm-like bodies (PLBs) could be regenerated with single treatments of PGRs [74,75]. Modifying a certain amount of PGRs in culture media has also enhanced protocorm proliferation efficiency, with several reports advocating the success of secondary protocorm mass proliferation from primary protocorms, mainly for epiphytic orchids [27,76]. Furthermore, cytokinin and auxin in the right balance reportedly influenced the organogenesis process, which is the process of plant organ development, such as shoots and roots. In this study, the lowest auxin concentration in the NAA-containing medium elicited the best response regarding P. labukensis protocorm proliferation. The response towards the NAA hormone was also the most significant in most of the concentrations employed. The findings in the current study were supported by a report by Prada and Pant [77], in which the MS medium supplemented with 0.5 mg/L NAA recorded the maximum rate of Cymbidium elegans protocorm multiplication.
Exogenous auxins, such as NAA, have been reported to result in inhibitory effects in explants of Oncidium Gower Ramsey, a species in the Oncidium genus [78]. In another study, Tao et al. [79] discovered that a medium supplemented with 0.5 mg/L NAA was the best at turning Cymbidium faveri Rolfe PLBs green after 30 DOC. Nevertheless, the report documented a downward PLB multiplication trend at higher NAA of concentrations.
In other research, high protocorm proliferation percentages also successfully found in media with 1.0 and 2.0 mg/L BAP [8,80]. Roy et al. [27] recorded the maximum rate of Vanda coerulea protocorm proliferation when cultivated in a culture treated with 2.0 mg/L BAP. Conversely, the current study documented the lowest percentage of protocorm proliferation of seedlings in the 2.0 mg/L BAP culture due to necrosis. The results were supported by the findings of another study, where NAA at higher concentrations exhibited inhibitory effects on PLB formation and maximum necrosis in Vanda testacea [81].
The combinations, ratios and concentrations of PGRs are critical to determining efficient plantlet regenerations [82]. Numerous researchers have recognised that the influence of a single PGR on shoot multiplication is better than the effects of a combination of PGRs in orchids [71,83]. Nevertheless, this study found that the media fortified with BAP and NAA produced the maximum shoot number. Consequently, the current study evaluated varying concentrations of BAP, NAA, and its combinations to obtain the optimal response on P. labukensis shoot and root induction.
The advantages of BAP and NAA on the growth and development of orchids have been reported by several studies [8,25,26]. In other study by Bhattacharyya et al. [84], they also agreed with the findings, where the addition of NAA and BAP in the culture medium led to optimum micropropagation of Vanda tesselata. The combination was also successfully employed by orchid breeders. Furthermore, shoot multiplications were documented in various orchid species when combinations of BAP and NAA were employed [85].
Generally, auxin stimulates root formation, while cytokinins enhance shoot development and cell division [86]. Similarly, this study observed that the 0.5 mg/L NAA induced and enhanced P. labukensis root formation, hence more roots were formed. Another study by Paudel et al. [87] supported the results, where the highest number of roots were documented when Esmeralda clarkei shoots were cultured in media supplemented with 0.5 and 1.0 mg/L NAA.
A report found that higher concentrations of PGRs increased the total phenolic content in the seed embryonic cells, while the oxidised phenol products inhibited cellular plant growth [88]. Auxin is a plant hormone widely utilised to induce adventitious rooting and control morphogenesis in micropropagation. Moreover, Bektas et al. [89] reported that BAP treatment was favoured since it elicited more shoots from O. coriophora protocorms.
The auxin-to-cytokinin ratio for shoot formation varies from species to species [90]. Several reports suggested that a 2:1 cytokinin-to-auxin combination was appropriate for shoot multiplication in some orchid species [67,91]. Nonetheless, the P. labukensis samples in this study preferred a low cytokinin concentration and high auxin concentration combination, whereas high auxin concentrations were advantageous for shoot and root formations.
The longest and highest number of roots were recorded by plantlets with the longest shoot, leaf, and root and most leaves and roots. The Dendrobium aqueum Lindley orchid documented a maximum of 6.25 and 5.25 roots per explant when cultivated in 7 and 3 mg/L of KIN. The species also averaged 0.98 cm roots in a KIN 5 mg/L medium [72]. Several studies also reported the superiority of PGR combinations for in vitro orchid shoot proliferation and development [92,93,94,95]. In the present study, the growing seedlings with well-developed long roots were transplanted into pots containing sand, brick, and soil (4:4:2) for acclimatisation. Nevertheless, none of the 10 transplanted plants survived after a fortnight under greenhouse conditions.
5. Conclusions
Understanding the different stage of seed development and their viability is crucial for plant production. The observation on capsule formation, indicates that the mature P. labukensis seeds at 120 DAP have reached optimal stage for viability. Meanwhile, the seeds at 60 DAP showed no response towards the viability test.
Selecting a basal medium for orchid seed germination relies on the specific requirements of the species and desired outcomes. In this study, KC basal medium was the most effective treatment for P. labukensis seed germination. Nevertheless, no significant difference was recorded with the other media evaluated. The incorporation of BH (5 g/L and 10 g/L) elicited the highest number of protocorm proliferation. The CW also demonstrated a remarkable influence on protocorm growth, where 70% responded after 200 DOC.
Protocorm proliferation and shoot and root formation responded significantly to PGR, which contained low concentrations of auxin, 0.5 and 1.0 mg/L NAA. The media added with the PGR exhibited superior protocorm growth and development after 240 DOC. Moreover, seedling development demonstrated a better response towards the hormone combination 1.0 mg/L NAA + 0.5 mg/L BAP (4.75 ± 0.50%). The 1.0 mg/L NAA + 0.5 mg/L BAP media recorded the highest root formation percentage at 8.75 ± 0.96%. Consequently, NAA and BAP could enhance shoot and root induction of the P. labukensis orchids.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9060681/s1, Figure S1: The Paraphaleonopsis labukensis (A) Four elongation of flowers; (B) Capsules formation after 30 days after pollination. Scale bar = 1 cm; Figure S2: P. labukensis capsule length and diameter measurement; Table S1: Media formulation for MS basal medium (Murashige and Skoog, 1962) [37]; Table S2: Media formulation for KC basal media (Knudson, 1945) [38]; Table S3: Media formulation for VW basal media (Vacin and Went, 1949) [39].
Author Contributions
Conceptualization, N.A.R., J.A.G. and H.V.N.; methodology, H.V.N. and N.A.R.; validation, H.V.N., N.A.R., A.A.M. and J.A.G.; formal analysis, H.V.N., A.A.M. and N.N.M.; resources, N.A.R. and J.A.G.; writing—original draft preparation, H.V.N. and N.A.R.; writing—review and editing, H.V.N., N.A.R., N.A.S. and N.N.M.; supervision, N.A.R.; project administration, N.A.S. and J.A.; funding acquisition, N.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was fully funded by research university grants from Universiti Malaysia Sabah (SBK0316-2017 and GUG 0284-2018).
Data Availability Statement
Not applicable.
Acknowledgments
The author extends his gratitude to Sabah Biodiversity Center (SaBC) for providing access license ref. no JKM/MBS.100-2/2JLD10(6).
Conflicts of Interest
The authors report there are no competing interests to declare.
References
- Utami, E.S.W.; Hariyanto, S. In vitro seed germination and seedling development of a rare Indonesian native orchid Phalaenopsis amboinensis J.J. Sm. Scietifica 2019, 2019, 8105138. [Google Scholar]
- Juiling, S.; Leon, S.K.; Jumian, J.; Tsen, S.; Lee, Y.L.; Khoo, E.; Sugau, J.B.; Nilus, R.; Pereira, J.T.; Damit, A. Conservation assessment and spatial distribution of endemic orchids in Sabah. Borneo. Nat. Conserv. Res. 2020, 5, 136–144. [Google Scholar] [CrossRef]
- Wood, J.; Beaman, J.; Beaman, R. The Plants of Mount Kinabalu 2. Orchids; Royal Botanic Garden: Richmond, UK, 1993. [Google Scholar]
- Chan, C.L.; Lamb, A.; Shim, P.S.; Wood, J.J. Orchids of Borneo: Introduction and Selection of Species; Print and Co. Sdn Bhd: Kuala Lumpur, Malaysia, 1994; p. 402. [Google Scholar]
- Swarts, N.D.; Dixon, K.W. Perspectives on orchid conservation in botanic gardens. Trends Plant Sci. 2009, 14, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Lamb, A. The conservation of orchids in Sabah (Malaysian Borneo). In Tropical Botanic Gardens: Their Roles in Conservation and Development; Heywood, V.H., Wyse Jackson, P.S., Eds.; Academic Press: London, UK, 1991; pp. 308–327. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-I. 2020. Available online: https://www.iucnredlist.org/ (accessed on 3 March 2023).
- Basker, S.; Bai, V.N. In vitro propagation of an epiphytic and rare orchid Eria bambusifolia Lindl. Res. Biotechnol. 2010, 1, 15–20. [Google Scholar]
- Chen, Y.; Goodale, U.; Fan, X.; Gao, J. Asymbiotic seed germination and in vitro seedling development of Paphiopedilum spicerianum: An orchid with an extremely small population in China. Glob. Ecol. Conserv. 2015, 3, 367–378. [Google Scholar] [CrossRef]
- Johnson, T.R.; Kane, M.E. Asymbiotic germination of ornamental Vanda: In vitro germination and development of three hybrids. Plant Cell Tissue Organ Cult. 2007, 91, 251–261. [Google Scholar] [CrossRef]
- Kahraman, M.U.; Cullum, F.J. Asymbiotic germination and seedling development of terrestrial orchid Bletilla striata using in vitro and ex vitro Cultures. Hortic. Stud. 2010, 38, 1–14. [Google Scholar] [CrossRef]
- Ferreira, W.D.M.; Oliveira, A.M.D.; Viana, J.C.; Suzuki, R.M.; Oliveira, J.R.G.D. Asymbiotic germination, initial development in vitro and acclimatisation of Cyrtopodium paludicolum Hoehne, a Brazilian Savanna orchid species. Rodriguésia 2022, 73, 1–14. [Google Scholar] [CrossRef]
- Barrientos, B.A.B.; Fang, J.Y. Influence of photoperiod and culture medium on the speed of asymbiotic seed germination and seedling development in Spathoglottis plicata. HortScience 2019, 54, 1570–1575. [Google Scholar] [CrossRef]
- Yang, N.; Wang, D.; Gao, Y.; Hu, E.; Yu, X.; Peng, S.; Ji, J.; Zhang, M.S. An efficient micropropagation protocol, chemical components, and hypoglycemic activity for Cremastra appendiculata (D. Don) Makino pseudobulbs. In Vitro Cell. Dev. Biol.-Plant 2022, 58, 213–224. [Google Scholar] [CrossRef]
- Yao, L.; Huang, J.; Zhang, S. An Improved Protocol for Asymbiotic seed germination and seedling development of Paphiopedilum tigrinum. Horticulturae 2021, 7, 298. [Google Scholar] [CrossRef]
- Dutra, D.; Johnson, T.R.; Kauth, P.J.; Stewart, S.L.; Kane, M.E.; Richardson, L. Asymbiotic seed germination, in vitro seedling development, and greenhouse acclimatisation of the threatened terrestrial orchid Bletia purpurea. Plant Cell Tissue Organ Cult. 2008, 94, 11–21. [Google Scholar] [CrossRef]
- Parthibhan, S.; Benjamin, J.F.; Muthukumar, M.; Ahamed, N.; Sherif, T.; Senthi Kumar, M.V.R. Influence of nutritional media and photoperiods on in vitro asymbiotic seed germination and seedling development of Dendrobium aqueum Lindley. Afr. J. Plant Sci. 2012, 6, 383–393. [Google Scholar] [CrossRef]
- Paul, S.; Kumaria, S.; Tandon, P. An effective nutrient medium for asymbiotic seed germination and large-scale in vitro regeneration of Dendrobium hookerianum, a threatened orchid of northeast India. AoB Plants 2012, 2012, plr032. [Google Scholar] [CrossRef]
- Jawan, R.; Gansau, J.A.; Abdullah, J.O. In vitro culture of Borneo’s endemic orchid, Vanda dearei. Asia-Pac. J. Mol. Biol. Biotechnol. 2010, 18, 203–207. [Google Scholar]
- Islam, M.; Akter, M.; Prodhan, A. Effect of potato extract on in vitro seed germination and seedling growth of local Vanda roxburgii orchid. J. Bangladesh Agric. Univ. 2012, 9, 211–215. [Google Scholar] [CrossRef]
- Dwiyani, R.; Yuswanti, H.; Darmawati, I.A.P.; Suada, K.; Mayadewi, N.N.A. In vitro germination and its subsequent growth of an orchid of Vanda tricolor Lindl. var. suavis from Bali on complex additives enriched medium. J. Agric. Sci. 2015, 37, 144–150. [Google Scholar] [CrossRef]
- Jadhav, S.G.; Chavan, D.A.; Waghmare, Y.M. Effect of plant spacing, growth regulator and nutrient management on yield, quality and economics of Bt cotton. J. Cotton Res. Dev. 2015, 29, 48–52. [Google Scholar]
- Bhattarai, P. Effects of plant growth regulators on growth and yield of pre-basic seed potato production under glasshouse condition. SAARC J. Agric. 2017, 15, 149–160. [Google Scholar] [CrossRef][Green Version]
- Chin, C.K.; Lee, Z.H.; Mubbarakh, S.A.; Antony, J.J.J.; Chew, B.L.; Subramaniam, S. Effects of plant growth regulators and activated charcoal on somaclonal variations of protocorm-like bodies (PLBs) of Dendrobium Sabin Blue orchid. Biocatal. Agric. Biotechnol. 2019, 22, 101426. [Google Scholar] [CrossRef]
- Hossain, M.M. Asymbiotic seed germination and in vitro seedling development of Epidendrum ibaguense Kunth. (Orchidaceae). Afr. J. Biotechnol. 2008, 7, 3614–3619. Available online: http://www.academicjournals.org/AJB (accessed on 1 September 2019).
- Mahendran, G.; Muniappan, V.; Ashwini, M.; Muthukumar, T.; Narmatha Bai, V. Asymbiotic seed germination of Cymbidium bicolor Lindl. (Orchidaceae) and the influence of mycorrhizal fungus on seedling development. Acta Physiol. Plant 2013, 35, 829–840. [Google Scholar] [CrossRef]
- Roy, A.R.; Patel, R.S.; Patel, V.V.; Sajeev, S.; Deka, B.C. Asymbiotic seed germination, mass propagation and seedling development of Vanda coerulea Griff ex.Lindl. (Blue Vanda): An in vitro protocol for an endangered orchid. Sci. Hortic. 2011, 128, 325–331. [Google Scholar] [CrossRef]
- Gallo, F.; Souza, L.; Milaneze, M.; Almeida, O. Seed structure and in vitro seedling development of certain Laeliinae species (Orchidaceae). Rev. Mex. Biodivers. 2016, 87, 68–73. [Google Scholar] [CrossRef]
- Gansau, J.A.; Indan, H.; Abdullah, S.N.; David, D.; Marbawi, H.; Jawan, R. Effects of organic additives and plant growth regulators on protocorm development of Dendrobium lowii. Trans. Sci. Technol. 2016, 3, 462–468. [Google Scholar]
- Reddy, J.; Niveshika; Shaju, A.; Jose, A.; Betty, A.; Yarmichon, H. Plant growth regulators used for in vitro micropropagation of orchids: A research review. Int. J. Biol. Res. 2021, 8, 37–42. [Google Scholar]
- Amiri, S.; Mohammadi, R. Establishment of an efficient in vitro propagation protocol for Sumac (Rhus coriaria L.) and confirmation of the genetic homogeneity. Sci Rep. 2021, 11, 173. [Google Scholar] [CrossRef]
- Salsabila, S.S.; Fatimah, K.; Noorhazira, S.; Halimatun, T.S.T.A.B.; Aurifullah, M.; Suhana, Z. Effect of coconut water and peptone in micropropagation of Phalaeonopsis amabilis (L.) Blume Orchid. IOP Conf. Ser. Earth Environ. Sci. 2022, 1102, 012002. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Chan, M.-T.; Sanjaya; Chai, M.-L.; Tanaka, M. Priming abiotic factors for optimal hybrid Cymbidium (Orchidaceae) PLB and callus induction, plantlet formation, and their subsequent cytogenetic stability analysis. Sci. Hortic. 2006, 109, 368–378. [Google Scholar] [CrossRef]
- Kauth, P.J.; Johnson, T.R.; Stewart, S.L.; Kane, M.E. A classroom exercise in hand pollination and in vitro asymbiotic orchid seed germination. Plant Cell Tissue Organ Cult. 2008, 93, 223–230. [Google Scholar] [CrossRef]
- Hosomi, S.T.; Santos, R.B.; Custódio, C.C.; Seaton, P.T.; Marx, P.R.; Machado-Neto, N.B. Pre-conditioning Cattleya seeds to improve the efficacy of the tetrazolium test for viability Seed. Seed Sci. Technol. 2011, 39, 178–189. [Google Scholar] [CrossRef]
- Alomia, Y.A.; Mosquera-E, A.T.; Flanagan, N.S.; Otero, J.T. Seed viability and symbiotic seed germination in Vanilla spp. (Orchidaceae). Res. J. Seed Sci. 2017, 10, 43–52. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Bot. Gaz. 1962, 15, 473–477. [Google Scholar]
- Knudson, L. A new nutrient solution for orchid seed germination. Bot. Gaz. 1946, 15, 214–217. [Google Scholar]
- Vacin, F.; Went, F.W. Some pH changes in nutrient solutions. Bot. Gaz. 1949, 110, 605–613. [Google Scholar] [CrossRef]
- Harrison, C.R.; Arditti, J. Physiological changes during the germination of Cattleya aurantiaca (Orchidaceae). Bot. Gaz. 1978, 139, 180–189. [Google Scholar] [CrossRef]
- Balilashaki, K.; Ghehsareh, M. Micropropagation of Phalaenopsis amabilis var. Manila by leaves obtained from in vitro culturing the nodes of flower stalks. Not. Sci. Biol. 2016, 8, 164–169. [Google Scholar] [CrossRef][Green Version]
- Edens-Meier, R.; Arduser, M.; Westhus, E.; Bernhardt, P. Pollination ecology of Cypripedium regina Walter (Orchidaceae): Size matters. Telopea 2010, 13, 327–340. [Google Scholar] [CrossRef]
- Muñoz, M.; Jiménez, V. Capsule development, in vitro germination and plantlet acclimatization in Phragmipedium humboldtii, P. longifolium and P. pearcei. Lankesteriana 2008, 8, 23–31. [Google Scholar] [CrossRef]
- Mohammad, N.N.; Rusdi, N.A. Scanning electron microscopy analysis of early floral development in Renanthera bella J.J. Wood, an Endemic orchid from Sabah. Pertanika J. Trop. Agric. Sci. 2020, 43, 77–389. [Google Scholar]
- Koene, F.M.; Amano, É.; Smidt, E.D.C.; Ribas, L.L.F. Asymbiotic germination and morphological studies of seeds of Atlantic Rainforest micro-orchids (Pleurothallidinae). PLoS ONE 2020, 15, e0243297. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Mercado, S.A. Germinación asimbiótica de semillas y desarrollo in vitro de plántulas de Cattleya mendelii Dombrain (Orchidaceae). Acta Agron 2012, 61, 69–78. [Google Scholar]
- Mayo-Mosqueda, A.; Maceda-López, L.F.; Andrade-Canto, S.B.; Noguera-Savelli, E.; Caamal-Velázquez, H.; Cano-Sosa, J.D.S.; Alatorre-Cobos, F. Efficient protocol for in vitro propagation of Laelia rubescens Lindl. from asymbiotic seed germination. S. Afr. J. Bot. 2020, 133, 264–272. [Google Scholar] [CrossRef]
- Lo, S.F.; Nalawade, S.M.; Kuo, C.L.; Chen, C.L.; Tsay, H.S. Asymbiotic germination of immature seeds, plantlet development and ex vitro establishment of plants of Dendrobium tosaense makino—A medicinally important orchid. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 528–535. [Google Scholar] [CrossRef]
- Doria, J. Generalidades sobre las semillas: Su producción, conservación y almacenamiento. Cultiv. Trop. 2010, 31, 74–85. [Google Scholar]
- Mercado, S.A.S.; Caleño, J.D.Q.; Rozo, L.Y.M. Improvement of the methodology of the tetrazolium test using different pretreatments in seeds of the genus Epidendrum (Orchidaceae). J. Seed Sci. 2020, 42, e202042013. [Google Scholar] [CrossRef]
- Hosomi, S.T.; Souza, T.B.; Custódio, C.C.; Machado-Neto, N.B. Refining the tetrazolium test for evaluation of Cattleya labiata and C. tigrina seeds viability. Aust. J. Crop Sci. 2017, 11, 1320–1326. [Google Scholar] [CrossRef]
- Hirano, T.; Godo, T.; Mii, M.; Ishikawa, K. Cryopreservation of immature seeds of Bletilla striata by vitrification. Plant Cell Rep. 2005, 23, 534–539. [Google Scholar] [CrossRef]
- Yamazaki, J.; Miyoshi, K. In vitro asymbiotic germination of immature seed and formation of protocorm by Cephalanthera falcata (Orchidaceae). Ann. Bot. 2006, 98, 1197–1206. [Google Scholar] [CrossRef]
- Filho, M.J. Seed vigor testing: An overview of the past, present and future perspective. Sci. Agric. 2015, 72, 363–374. [Google Scholar] [CrossRef]
- Singh, F. Differential staining of orchid seeds for viability testing. Am. Orchid. Soc. Bull. 1981, 50, 416–418. [Google Scholar]
- Lauzer, D.; St-Arnaud, M.; Barabeâ, D. Tetrazolium staining and in vitro germination of mature seeds of Cypripedium acaule (Orchidaceae). Lindleyana 1994, 9, 197–204. [Google Scholar]
- Vujanovic, V.; St-Arnaud, M.; Barabé, D.; Thibeault, G. Viability testing of orchid seed and the promotion of colouration and germination. Ann. Bot. 2000, 86, 79–86. [Google Scholar] [CrossRef]
- Arditti, J.; Ernst, R. Physiology of germinating orchid seeds. In Orchid Biology, Reviews and Perspectives III; Arditti, J., Ed.; Cornell University Press: Ithaca, NY, USA, 1984; pp. 177–222. [Google Scholar]
- Zeigler, E.; Grivet, C.; Assmann, S.M.; Deitzer, G.F.; Hannegan, M.W. Stomatal limitation of carbon gain in Paphiopedilum sp. (Orchidaceae) and its reversal by blue light. Plant Physiol. 1985, 77, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Van Waes, J.M.; Debergh, P.C. In vitro germination of some Western European orchids. Physiol. Plant. 1986, 67, 253–261. [Google Scholar] [CrossRef]
- Bhowmik, T.K.; Rahman, M.M. Effect of different basal media and PGRs on in vitro seed germination and seedling development of medicinally important orchid Cymbidium aloifolium. J. Pharmacogn. Phytochem. 2017, 6, 167–172. [Google Scholar]
- Stewart, J. Orchid propagation by tissue culture techniques-past, present and future. In Modern Methods in Orchid Conservation: The Role of Physiology, Ecology and Management; Pritchard, H.W., Ed.; Cambridge University Press: Cambridge, UK, 1989; pp. 87–100. [Google Scholar]
- Suzuki, R.M.; Moreira, V.C.; Pescador, R.; Melo Ferreira, W. Asymbiotic seed germination and in vitro seedling development of the threatened orchid Hoffmannseggella cinnabarina. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 500–551. [Google Scholar] [CrossRef]
- Zeng, S.; Wu, K.L.; Teixeira Da Silva, J.; Zhang, J.X.; Chen, Z.L.; Xia, N.H.; Duan, J. Asymbiotic seed germination, seedling development and reintroduction of Paphiopedilum wardii Sumerh., an endangered terrestrial orchid. Sci. Hortic. 2012, 138, 198–209. [Google Scholar] [CrossRef]
- Nambiar, N.; Tee, C.S.; Maziah, M. Effects of organic additives and different carbohydrate sources on proliferation of protocorm like bodies in Dendrobium Alya Pink. Plant Omics 2012, 5, 10–18. [Google Scholar]
- Teixeira Da Silva, J.A. Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues, Vol I–IV.; Global Science Books: London, UK, 2006; p. 2506. [Google Scholar]
- Gnasekaran, P.; Poobathy, R.; Mahmood, M.; Samian, M.R.; Subramaniam, S. Effects of complex organic additives on improving the growth of PLBs of Vanda Kasem’s delight. Aust. J. Crop Sci. 2012, 6, 1245–1248. [Google Scholar]
- Tawara, S.; Suraninpong, P.; Chanprame, S. Germination and Regeneration of Cymbidium findlaysonianum Lindl. on a medium supplemented with some organic sources. Walailak J. Sci. Technol. 2008, 5, 125–135. [Google Scholar]
- Van Winkle, S.C.; Pullman, G.S. Achieving desired plant growth regulator levels in liquid plant tissue culture media that include activated carbon. Plant Cell Rep. 2005, 22, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Banerjee, N. Induction of callus and plant regeneration from shoot tip explant of Dendrobium fimbriatum. Lindl. Var Octum. Sci. Hortic. 2003, 97, 333–340. [Google Scholar] [CrossRef]
- De Paw, M.A.; Remphrey, W.R.; Palmer, C.E. The cytokinin preference for in vitro germination and protocorm growth of Cypripedium candidum. Ann. Bot. 1995, 75, 267–275. [Google Scholar] [CrossRef]
- Parthibhan, S.; Rao, M.V.; Kumar, T.S. In vitro regeneration from protocorms in Dendrobium aqueum Lindley-An imperiled orchid. J. Genet. Eng. Biotechnol. 2015, 13, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Sumontip, B.; Warisa, P. Agrobacterium-mediated transformation of Dendrobium chrysotoxum Lindl. Afr. J. Biotechnol. 2012, 11, 2472–2476. [Google Scholar] [CrossRef]
- Khoddamzadeh, A.A.; Sinniah, U.R.; Kadir, M.A.; Kadzimin, S.B.; Mahmood, M.; Sreeramanan, S. In vitro induction and proliferation of protocorm-like bodies (PLBs) from leaf segments of Phalaenopsis bellina (Rchb.f.) Christenson. Plant Growth Regul. 2011, 65, 381–387. [Google Scholar] [CrossRef]
- Sujjartitthurakarn, P.; Kanchanapoom, K. Efficient direct protocorm-like bodies induction of dwarf Dendrobium using Thidiazuron. Not. Sci. Biol. 2011, 3, 88–92. [Google Scholar] [CrossRef][Green Version]
- Hossain, M.M. In vitro Embryo Morphogenesis and Micropropagation of Dendrobium aggregatum Roxb. Plant Tissue Cult. Biotechnol. 2013, 23, 241–249. [Google Scholar] [CrossRef]
- Pradha, S.; Pant, B. In vitro seed germination in Cymbidium elegans Lindl. and Dendrobium densiflorum Lindl. ex Wall. (Orchidaceae). Botanica Orientalis. J. Plant Sci. 2010, 6, 100–102. [Google Scholar] [CrossRef]
- Chen, J.T.; Chang, W.C. Effects of auxins and cytokinins on direct somatic embryogenesis on leaf explants of Oncidium “Gower Ramsey”. Plant Growth Regul. 2001, 34, 29–232. [Google Scholar] [CrossRef]
- Tao, J.; Yu, L.; Kong, F.; Zhao, D. Effects of plant growth regulators on in vitro propagation of Cymbidium faberi Rolfe. Afr. J. Biotech. 2011, 10, 15639–15646. [Google Scholar] [CrossRef]
- Devina, D.; Jualang, A.G.; Janna, O.A. Effect of NAA and BAP on protocorm proliferation of Borneo scented orchid, Vanda helvola. Asia-Pac. J. Mol. Biol. Biotechnol. 2010, 16, 221–224. [Google Scholar]
- Kaur, S.; Bhutani, K.K. In vitro conservation and asymbiotic propagation of Coelogyne flaccida (Lindl.): A threatened orchid. Plant Biosyst. 2014, 148, 935–944. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sharma, M.; Teixeira Da Silva, J.; Pathak, P. Seed germination and tissue culture of Cymbidium giganteum Wall. ex Lindl. Sci. Hortic. 2010, 123, 479–487. [Google Scholar] [CrossRef]
- Mahendran, G.; Bai, V.N. Mass propagation of Satyrium nepalense D. Don. A medicinal orchid via seed culture. Sci. Hortic. 2009, 119, 203–207. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Kumaria, S.; Tandon, P. High frequency regeneration protocol for Dendrobium nobile: A model tissue culture approach for propagation of medicinally important orchid species. S. Afr. J. Bot. 2016, 104, 232–243. [Google Scholar] [CrossRef]
- Long, B.; Niemiera, A.X.; Cheng, Z.Y.; Long, C.L. In vitro propagation of four threatened Paphiopedilum species (Orchidaceae). Plant Cell Tissue Organ Cult. 2010, 101, 151–162. [Google Scholar] [CrossRef]
- Wotavová-Novotná, K.; Vejsadová, H.; Kindlmann, P. Effects of sugars and growth regulators on in vitro growth of Dactylorhiza species. Biol. Plant. 2007, 51, 198–200. [Google Scholar] [CrossRef]
- Paudel, M.R.; Pant, B. A reliable protocol for micropropagation of Esmeralda Clarkei Rchb.f. (Orchidaceae). Asia-Pac. J. Mol. Biol. Biotechnol. 2013, 21, 114–120. [Google Scholar]
- Santana-Buzzy, N.; Rojas-Herrera, R.; Galaz-Ávalos, R.M.; Ku-Cauich, J.R.; Mijangos-Cortés, J.; Gutiérrez-Pacheco, L.C.; Canto, A.; Quiroz-Figueroa, F.; Loyola-Vargas, V.M. Advances in coffee tissue culture and its practical applications. In Vitro Cell. Dev. Biol. 2007, 43, 507–520. [Google Scholar] [CrossRef]
- Bektaş, E.; Cüce, M.; Sökmen, A. In vitro germination, protocorm formation, and plantlet development of Orchis coriophora (Orchidaceae), A naturally growing orchid species in Turkey. Turk. J. Bot. 2013, 37, 336–342. [Google Scholar] [CrossRef]
- Asa, M.; Kaviani, B. In vitro propagation of orchid Phalaenopsis amabilis (L.) Blume var. Jawa. Iran. J. Plant Physiol. 2020, 10, 3113–3123. [Google Scholar] [CrossRef]
- Decruse, S.W.; Gangaprasad, A.; Seeni, S.; Menon, V.S. Micropropagation and ecorestoration of Vanda spathulata, an exquisite orchid. Plant Cell Tissue Organ Cult. 2003, 41, 924–927. [Google Scholar] [CrossRef]
- Seeni, S.; Latha, P.G. In vitro multiplication and ecorehabilitation of the endangered Blue Vanda. Plant Cell Tissue Organ Cult. 2000, 61, 1–8. [Google Scholar] [CrossRef]
- Takamura, T.; Tanaka, M. Callus formation and plant regeneration from callus through somatic embryo structures in Cymbidium orchid. Plant Sci. 2004, 166, 1443–1449. [Google Scholar] [CrossRef]
- Malabadi, R.B.; Teixeira Da Silva, J.A.; Nataraja, K.; Mulgund, G.S. Shoot tip transverse thin cell layers and 24-epibrassinolide in the micropropagation of Cymbidium bicolor Lindl. Floric. Ornam. Biotech 2008, 2, 44–48. [Google Scholar]
- Baker, A.; Kaviani, B.; Nematzadeh, G.; Negahdar, N. Micropropagation of Orchis catasetum–a rare and endangered orchid. Acta Sci. Pol. Hortorum Cultus. 2014, 13, 197–205. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).