
Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Extraction and Genotyping
2.3. Data Analysis
3. Results
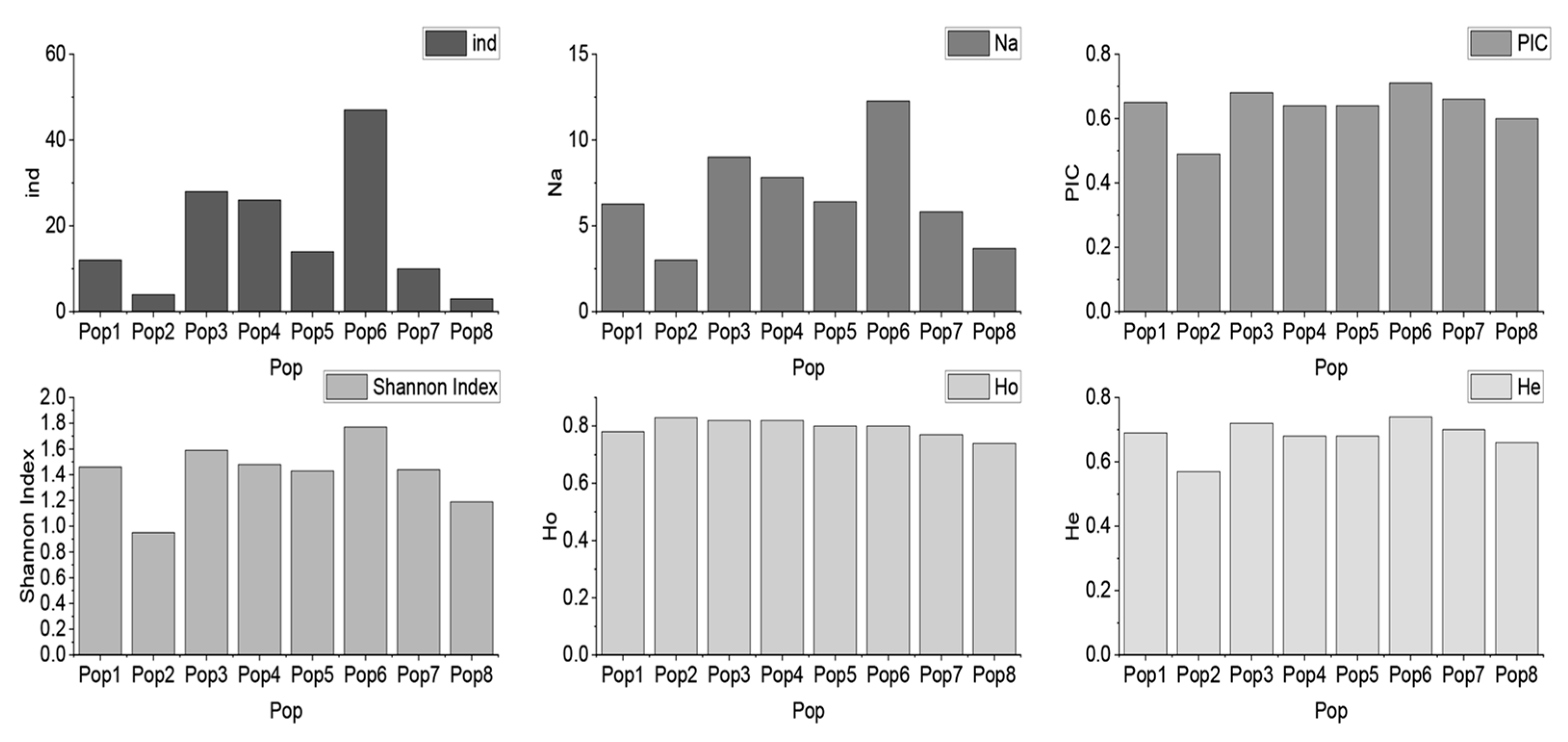
3.1. Genetic Diversity
3.2. Genetic Differentiation and Gene Flow
3.3. Genetic Distance and Cluster Structure
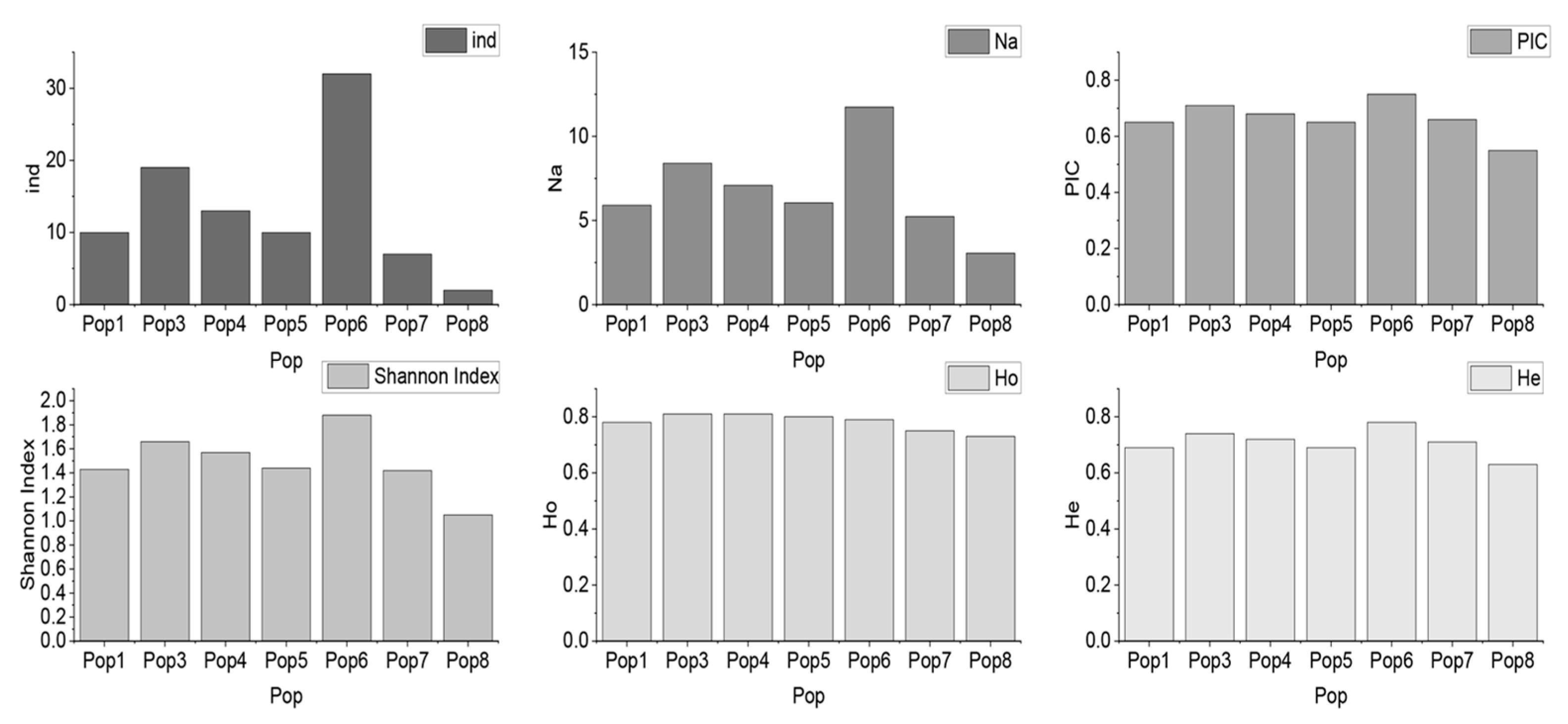
3.4. Construction of the Core Collection
3.5. Principal Component Analysis Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Onofrio, C.; de Lorenzis, G.; Giordani, T.; Natali, L.; Cavallini, A.; Scalabrelli, G. Retrotransposon-based molecular markers for grapevine species and cultivars identification. Tree Genet. Genomes 2010, 6, 451–466. [Google Scholar] [CrossRef]
- Bianchi, D.; Bran Cadoro, L.; de Lorenzis, G. Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers. Diversity 2020, 12, 103. [Google Scholar] [CrossRef] [Green Version]
- Li, M.F.; Zheng, X.Q. Research progress of methods of SSR primers development. Hereditas 2004, 26, 769–776. [Google Scholar] [PubMed]
- Boz, Y.; Bakir, M.; Çelikkol, B.P.; Kazan, K.; Yilmaz, F.; Çakir, B.; Aslantaş, Ş.; Söylemezoğlu, G.; Yaşasin, A.S.; Özer, C.; et al. Genetic characterization of grape (Vitis vinifera L.) germplasm from Southeast Anatolia by SSR markers. Vitis 2011, 50, 99–106. [Google Scholar]
- Zhang, Z.C.; Hou, X.L. Strategies for development of SSR molecular markers. Hereditas 2004, 26, 763–768. [Google Scholar]
- Goto-Yamamoto, N.; Azuma, A.; Mitani, N.; Kobayashi, S. SSR genotyping of wild grape species and grape cultivars of Vitis vinifera and V. vinifera × V. labrusca. J. Jpn. Soc. Hortic. Sci. 2013, 82, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Villano, C.; Carputo, D.; Frusciante, L.; Santoro, X.; Aversano, R. Use of SSR and retrotransposon-based markers to interpret the population structure of native grapevines from Southern Italy. Mol. Biotechnol. 2014, 56, 1011–1020. [Google Scholar] [CrossRef]
- Li, B.; Jiang, J.; Fan, X.; Zhang, Y.; Sun, H.; Zhang, G.; Liu, C. Molecular characterization of Chinese grape landraces (Vitis L.) using microsatellite DNA markers. HortScience 2017, 52, 533–540. [Google Scholar] [CrossRef]
- Zhong, H.; Zhang, F.; Zhou, X.; Pan, M.; Xu, J.; Hao, J.; Han, S.; Mei, C.; Xian, H.; Wang, M.; et al. Genome-wide identification of sequence variations and SSR marker development in the Munake grape cultivar. Front. Ecol. Evol. 2021, 9, 664835. [Google Scholar] [CrossRef]
- Castelli, M.; Cattaneo, G.; Manzoni, L.; Vanneschi, L. A distance between populations for n-points crossover in genetic algorithms. Swarm Evol. Comput. 2018, 106, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot 1997, 129, 157. [Google Scholar]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYSpc Numerical Taxonomy and Multivariate Analysis System Version 2.0 User Guide; Exeter Software: Setauket, NY, USA, 1998. [Google Scholar]
- Zheng, Y.; Xu, S.; Liu, L.; Zhao, Y.; Liu, J. Genetic diversity and population structure of Chinese natural bermudagrass [Cynodon dactylon (L.) Pers.] germplasm based on SRAP markers. PLoS ONE 2017, 12, e0177508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.R.; Cain, P.; Scott, N.S. DNA typing of grapevines: A universal methodology and database for describing cultivars and evaluating genetic relatedness. Plant Mol. Biol. 1994, 25, 939–949. [Google Scholar] [CrossRef]
- Vouillamoz, J.F.; Schneider, A.; Grando, M.S. Microsatellite analysis of alpine grape cultivars (Vitis vinifera L.): Alleged descendants of Pliny the Elder’s Raetica are genetically related. Genet. Resour. Crop Evol. 2007, 54, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, G.; Marrazzo, M.T.; Di Gaspero, G.; Pfeiffer, A.; Morgante, M.; Testolin, R. A set of microsatellite markers with long core repeat optimized for grape (Vitis spp.) genotyping. BMC Plant Biol. 2008, 8, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipriani, G.; Spadotto, A.; Jurman, I.; Di Gaspero, G.; Crespan, M.; Meneghetti, S.; Frare, E.; Vignani, R.; Cresti, M.; Morgante, M.; et al. The SSR-based molecular profile of 1005 grapevine (Vitis vinifera L.) accessions uncovers new synonymy and parentages, and reveals a large admixture amongst varieties of different geographic origin. Theor. Appl. Genet. 2010, 121, 1569–1585. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.L.; Zhang, J.Y.; Liu, C.H.; Zhang, G.H.; Li, M. Genetic relationships of Chinese grape accessions to European and American cultivars assessed by microsatellite markers. Biotechnol. Biotechnol. Equip. 2010, 24, 2054–2059. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.L.; Zhang, Q.; Zhang, G.H. Characterization of grape cultivars from China using microsatellite markers. Czech J. Genet. Plant Breed 2013, 49, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Acosta, K.G.; Riaz, S.; Franco-Mora, O.; Cruz-Castillo, J.G.; Walker, M.A. The genetic diversity of wild grapes in Mexico. Genet. Resour. Crop Evol. 2022, 69, 1329–1347. [Google Scholar] [CrossRef]
- Zheng, X.; Shi, J.; Yu, Y.; Shen, Y.; Tan, B.; Ye, X.; Li, J.; Feng, J. Exploration of Elite Stilbene Synthase alleles for resveratrol concentration in wild Chinese Vitis spp. and Vitis cultivars. Front. Plant Sci. 2017, 8, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laiadi, Z.; Bentchikou, M.M.; Bravo, G.; Cabello, F.; Martinez-Zapater, J.M. Molecular identification and genetic relationship of Algerian grapevine cultivars maintained at the germplasm collection of Skikda (Algeria). Vitis 2009, 48, 25–32. [Google Scholar]
- Štajner, N.; Tomić, L.; Ivanišević, D.; Korać, N.; Cvetković-Jovanović, T.; Beleski, K.; Angelova, E.; Maraš, V.; Javornik, B. Microsatellite inferred genetic diversity and structure of Western Balkan grapevines (Vitis vinifera L.). Tree Genet. Genomes 2014, 10, 127–140. [Google Scholar] [CrossRef]
- Fan, X.C.; Sun, H.S.; Zhang, Y.; Jiang, J.F.; Li, M.; Liu, C.H. Genetic diversity of Vitis davidii accessions revealed using microsatellite and sequence-related amplified polymorphism markers. HortScience 2018, 53, 283–287. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, G.L.; de Souza, A.P.; de Oliveira, F.A.; Zucchi, M.I.; de Souza, L.M.; Moura, M.F. Genetic structure and molecular diversity of Brazilian grapevine germplasm: Management and use in breeding programs. PLoS ONE 2020, 15, e0240665. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Cunha, J.; Ibáñez, J.; Teixeira-Santos, M.; Brazão, J.; Fevereiro, P.; Martínez-Zapater, J.M.; Eiras-Dias, J.E. Genetic Relationships among Portuguese Cultivated and Wild Vitis vinifera L. Germplasm. Front. Plant Sci. 2020, 11, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Cretazzo, E.; Moreno Sanz, P.; Lorenzi, S.; Benítez, M.L.; Velasco, L.; Emanuelli, F. Genetic characterization by SSR markers of a comprehensive wine grape collection conserved at Rancho de la Merced (Andalusia, Spain). Plants 2022, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- Sefc, K.M.; Lopes, M.S.; Lefort, F.; Botta, R.; Roubelakis-Angelakis, K.A.; Ibáñez, J.; Pejić, I.; Wagner, H.W.; Glössl, J.; Steinkellner, H. Microsatellite variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor. Appl. Genet. 2000, 100, 498–505. [Google Scholar] [CrossRef]
- Dallakyan, M.; Esoyan, S.; Gasparyan, B.; Smith, A.; Hovhannisyan, N. Genetic diversity and traditional uses of aboriginal grape (Vitis vinifera L.) varieties from the main viticultural regions of Armenia. Genet. Resour. Crop Evol. 2020, 67, 999–1024. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Fanelli, V.; Roseti, V.; Savoia, M.A.; Miazzi, M.M.; Venerito, P.; Savino, V.N.; Pirolo, C.; La Notte, P.; Falbo, M.; Petrillo, F.; et al. New insight into the identity of Italian grapevine varieties: The case study of Calabrian germplasm. Agronomy 2021, 11, 1538. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, L.; Fan, X.; Jiang, J.; Zheng, X.; Sun, H.; Liu, C. Genome-wide assessment of population structure, linkage disequilibrium and resistant QTLs in Chinese wild grapevine. Sci. Hortic. 2017, 215, 59–64. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Abd El-Zaher, M.; Salwa, B.; Reda, S. Isozyme analysis of genetic variability and population structure of Lactuca L. germplasm. Biochem. Syst. Ecol. 2016, 70, 73–79. [Google Scholar] [CrossRef]
- El-Esawi, M.; El-Esawi, A.; Glascoe, D.; Engle, T.; Ritz, J.; Link, M. Ahmad. Cellular metabolites modulate in vivo signaling of Arabidopsis cryptochrome-1. Plant Signal Behav. 2015, 10, e1063758. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Zhang, Q.; Tang, S.; Liao, W. Genetic and ecophysiological evidence that hybridization facilitated lineage diversification in yellow Camellia (Theaceae) species: A case study of natural hybridization between C. micrantha and C. flavida. BMC Plant Biol. 2023, 23, 154. [Google Scholar] [CrossRef]
- Magris, G.; Di Gaspero, G.; Marroni, F.; Zenoni, S.; Tornielli, G.B.; Celii, M.; De Paoli, E.; Pezzotti, M.; Conte, F.; Paci, P.; et al. Genetic epigenetic and genomic effects on variation of gene expression among grape varieties. Plant J. 2019, 99, 895–909. [Google Scholar] [CrossRef]
- Buhner-Zaharieva, T.; Moussaoui, S.; Lorente, M.; Andreu, J.; Nunez, R.; Ortiz, J.M.; Gogorcena, Y. Preservation and Molecular Characterization of Ancient Varieties in Spanish Grapevine Germplasm Collections. Am. J. Enol. Vitic. 2010, 3, 328–335. [Google Scholar] [CrossRef]
- Mercati, F.; De Lorenzis, G.; Brancadoro, L.; Lupini, A.; Abenavoli, M.R.; Barbagallo, M.G.; Di Lorenzo, R.; Scienza, A.; Sunseri, F. High-throughput 18K SNP array to assess genetic variability of the main grapevine cultivars from Sicily. Tree Genet. Genomes 2016, 12, 59. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Fan, X.; Dong, Y.; Zhang, Z.; Ren, F.; Hu, G.; Li, Z. Complete nucleotide sequence of a new variant of grapevine fanleaf virus from northeastern China. Arch. Virol. 2017, 162, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Karn, A.; Reisch, B.; Nguyen, A.; Sun, Y.; Bao, Y.; Campbell, M.S.; Church, D.; Williams, S.; Xu, X.; et al. Haplotyping the Vitis collinear core genome with rhAmp Seq improves marker transferability in a diverse genus. Nat. Commun. 2020, 11, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Ge, T.; Li, Y.; Hou, X. Genome-wide identification of SSR and SNP markers from the non-heading Chinese cabbage for comparative genomic analyses. BMC Genom. 2015, 16, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbaiyan, G.K.; Waters, D.L.E.; Katiyar, S.K.; Sadananda, A.R.; Vaddadi, S.; Henry, R.J. Genome-wide DNA polymorphisms in elite indica rice inbreds discovered by whole-genome sequencing. Plant Biotechnol. J. 2012, 10, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Cao, X.; Wu, D.; Hui, M.; Han, X.; Yao, F.; Li, Y.; Li, H.; Wang, H. Screening and Validation of SSR Molecular Markers for Identification of Downy Mildew Resistance in Intraspecific Hybrid F1 Progeny (V. vinifera). Horticulturae 2022, 8, 706. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop | Number | Latitude | Longitude |
|---|---|---|---|
| 1 | 106–117 | 86.0987 | 41.6871 |
| 2 | 27–30 | 76.1566 | 39.6706 |
| 75.7933 | 39.6632 | ||
| 3 | 78–105 | 76.1237 | 39.5622 |
| 76.0296 | 39.4867 | ||
| 4 | 1–26 | 80.1261 | 40.9392 |
| 80.0633 | 40.8863 | ||
| 5 | 128–141 | 88.1314 | 39.0237 |
| 6 | 31–77 | 79.8644 | 37.0841 |
| 79.6770 | 37.1756 | ||
| 79.6564 | 37.1431 | ||
| 7 | 118–127 | 85.5674 | 38.3319 |
| 85.5388 | 34.3438 | ||
| 8 | 142–144 | 82.1590 | 44.9062 |
| Name | Primer Sequences (F) (5′-3′) | Primer Sequences (R) (5′-3′) |
|---|---|---|
| Vchr16a | TTCATGTGTGACACCCCTTT | AATGTCCATGCTTCAAAATACC |
| Scu06 | CCTAATGCCAGGAAGGTTGC | CCCTAGTCTCTCTACCTATCCATG |
| VVMD5 | CTAGAGCTACGCCAATCCAA | TATACCAAAAATCATATTCCTAAA |
| VVMD27 | TACCAGATCTGAATACATCCGTAAGT | ACGGGTATAGAGCAAACGGTGT |
| VVMD32 | GGAAAGATGGGATGACTCGC | TATGATTTTTTAGGGGGGTGAGG |
| VVS2 | CAGCCCGTAAATGTATCCATC | AAATTCAAAATTCTAATTCAACTGG |
| Vchr8a | ACCCACTGCCACTCTCTCAT | AAATCTCCGGGATCCTTTTG |
| VVMD7 | AGAGTTGCGGAGAACAGGAT | CGAACCTTCACACGCTTGAT |
| VVIP31 | TATCCAAGAGACAAATTCCCAC | TTCTCTTGTTTCCTGCAAATGG |
| Vchr13a | TGGCAGAGCAAATGAATCAA | TTGGATGGATTGGAATGACC |
| Vchr6a | AATGTTGAGCTTTGGGCTTG | CCAATTCTTCCATACCTCAAAA |
| VMC4F3-1 | AAAGCACTATGGTGGGTGTAAA | TAACCAATACATGCATCAAGGA |
| VRZAG67 | ACCTGGCCCGACTCCTCTTGTATGC | TCCTGCCGGCGATAACCAAGCTATG |
| SCU15W | GCCTATGTGCCAGACCAAAAAC | TTGGAAGTAGCCAGCCCAACCTTC |
| VMC5G8 | CATGCACATCTTGTTTCACTCT | CATCATTGCTTCCAAAAGTCTC |
| T1 | GTGTGCCTACATTTTTCATTCGT | AACAATATGGCACAACAATGTCA |
| T3 | TCAAAAAGAAATAATATTAGATGCGG | AATTCCAAAATCCCAACTTTCTC |
| T4 | CTAATATCGCGATTCACAAATCA | AAAATTGATCAAAACTCATGAAAATG |
| T5 | CCAGTGCTACAAAAACTCTTGCT | GTTGATTTGGAAGCTGAAAATTG |
| T6 | GCCTTTATCTAGAAGCCCTCACT | CAACATAAGAATAGGTAGCATCG |
| T7 | CTTTCTCGAAATTTCCGATTTG | AGAAAACCCTTTGCAGCAGTAATATGG |
| T8 | CCCCAAAATGTATCCCAATTTTA | TTTGGAGACAATGAATGGATAGG |
| Sample Intensity | 0.1 | 0.15 | 0.2 | 0.25 | 0.3 | 0.35 | 0.4 | 0.45 | 0.5 | 0.55 | 0.6 | 0.65 | 0.7 | 0.75 | 0.8 | 0.85 | 0.9 | 0.95 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | 14 | 21 | 28 | 36 | 43 | 50 | 57 | 64 | 72 | 79 | 86 | 93 | 100 | 108 | 115 | 122 | 129 | 136 |
| CV | 0.59 | 0.67 | 0.72 | 0.78 | 0.81 | 0.83 | 0.87 | 0.88 | 0.92 | 0.93 | 0.94 | 0.95 | 0.95 | 0.96 | 0.97 | 0.98 | 0.99 | 1 |
| Pop | ind | Na | PIC | Shannon Index | Ho | He |
|---|---|---|---|---|---|---|
| Pop1 | 12 | 6.27 | 0.65 | 1.46 | 0.78 | 0.69 |
| Pop2 | 4 | 3.00 | 0.49 | 0.95 | 0.83 | 0.57 |
| Pop3 | 28 | 9.00 | 0.68 | 1.59 | 0.82 | 0.72 |
| Pop4 | 26 | 7.82 | 0.64 | 1.48 | 0.82 | 0.68 |
| Pop5 | 14 | 6.41 | 0.64 | 1.43 | 0.80 | 0.68 |
| Pop6 | 47 | 12.27 | 0.71 | 1.77 | 0.80 | 0.74 |
| Pop7 | 10 | 5.82 | 0.66 | 1.44 | 0.77 | 0.70 |
| Pop8 | 3 | 3.68 | 0.60 | 1.19 | 0.74 | 0.66 |
| Total | 144 | 18.14 | 0.68 | 1.76 | 0.80 | 0.73 |
| Pop | ind | Na | PIC | Shannon Index | Ho | He |
|---|---|---|---|---|---|---|
| Pop1 | 10 | 5.91 | 0.65 | 1.43 | 0.78 | 0.69 |
| Pop3 | 19 | 8.41 | 0.71 | 1.66 | 0.81 | 0.74 |
| Pop4 | 13 | 7.09 | 0.68 | 1.57 | 0.81 | 0.72 |
| Pop5 | 10 | 6.05 | 0.65 | 1.44 | 0.80 | 0.69 |
| Pop6 | 32 | 11.73 | 0.75 | 1.88 | 0.79 | 0.78 |
| Pop7 | 7 | 5.23 | 0.66 | 1.42 | 0.75 | 0.71 |
| Pop8 | 2 | 3.05 | 0.55 | 1.05 | 0.73 | 0.63 |
| Total | 93 | 17.27 | 0.74 | 1.92 | 0.79 | 0.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhong, H.; Zhang, F.; Wang, X.; Wu, X.; Wang, J.; Shi, W. Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers. Horticulturae 2023, 9, 917. https://doi.org/10.3390/horticulturae9080917
Liu S, Zhong H, Zhang F, Wang X, Wu X, Wang J, Shi W. Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers. Horticulturae. 2023; 9(8):917. https://doi.org/10.3390/horticulturae9080917
Chicago/Turabian StyleLiu, Shiqing, Haixia Zhong, Fuchun Zhang, Xiyong Wang, Xinyu Wu, Jiancheng Wang, and Wei Shi. 2023. "Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers" Horticulturae 9, no. 8: 917. https://doi.org/10.3390/horticulturae9080917
APA StyleLiu, S., Zhong, H., Zhang, F., Wang, X., Wu, X., Wang, J., & Shi, W. (2023). Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers. Horticulturae, 9(8), 917. https://doi.org/10.3390/horticulturae9080917