Abstract
Plant regeneration through somatic embryogenesis provides a solution for maintaining and genetically improving crop or fruit varieties with desirable agronomic traits. For the fruit tree pomegranate (Punica granatum L.), despite some successful applications, the existing somatic embryogenesis protocols are limited by low availability of explants and susceptibility to browning. To address these problems, in this study, we developed an effective system for induction of high-vigor pomegranate somatic embryos derived from stem explants. The usage of stem explants breaks through the difficulty in obtaining material, thus making our system suitable for widespread commercial production. To enhance the performance of our system, we identified the optimal explants, subculture cycles and combination of basal media and plant growth regulators for each step. The results showed that inoculating stem explants onto a Murashige and Skoog (MS) medium supplemented with 1.0 mg/L 6-benzylaminopurine (6-BA) and 1.0 mg/L 1-naphthaleneacetic acid (NAA) achieved the best induction rate and growth status of pomegranate calli (induction rate = ~72%), and MS medium containing 0.5 mg/L 6-BA and 1.0 mg/L NAA was the optimal condition for the induction of embryogenic calli and somatic embryos (induction rate = ~74% and 79%, respectively). The optimal subculture period for embryogenic calli was found to be 30–35 days. Strong roots were then induced in the developed somatic embryo seedlings, which survived and grew well after transplantation to the natural environment, indicating the good vitality of the induced pomegranate somatic embryos. Together, our system provides a solution to mass somatic embryo induction and plant regeneration of pomegranate and lays a foundation for future genetic transformation and bioengineering improvement of pomegranate with favorable agronomic traits.
1. Introduction
Pomegranate (Punica granatum L.) is one of the most popular fruits worldwide; it has integrated economic, ecological, and social benefits, as well as ornamental and health applications [1,2]. In China, the pomegranate is widely cultivated throughout Sichuan and Yunnan in the southwest, Shanxi in the north, and Shandong and Anhui in the east [3]. However, due to limited technologies, there are many challenges in the cultivation and management of pomegranate trees, such as low vigor and vitality, that greatly restrict the production of pomegranate fruits, especially for rare varieties. Importantly, some pomegranate varieties have been found to present special characteristics, such as dwarfed stature, white berries, or high vitamin content, which are beneficial to their yield and quality [4]. Thus, screening and breeding varieties with favorable agronomic or economic traits have attracted great interests from the pomegranate industry.
Plant regeneration through somatic embryogenesis provides a solution to improve and breed desirable varieties [5]. Without fertilization, somatic embryos are generated by dedifferentiated plant somatic cells and present a capacity to eventually develop into complete plants [6]. Compared to zygotic embryos, somatic embryos have an edge in high heritability from the mother plant that allows it to avoid genetic segregation during sexual reproduction, and maintain desirable agronomic traits [7,8,9]. Furthermore, somatic embryogenesis can be used for genetic transformation, for example, via Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-mediated genome editing, which facilitates the genetic improvement of favorable phenotypes [10,11]. Hence, somatic embryogenesis provides a valuable model for embryo cell biology and molecular biology research [12]. Somatic embryogenesis can be carried out via direct or indirect strategies. Under certain conditions, somatic embryos can be directly induced from explant embryogenesis. For the indirect strategy, a new somatic embryo can be induced from a primary somatic embryo, either with or without a callus stage, or from an embryogenic callus [13,14]. Compared to primary somatic embryos or embryogenic calli, secondary somatic embryoids exhibit greater vitality and longer-lasting regeneration ability, which are better for transformative genetic research and germplasm conservation [15]. In addition, indirect somatic embryogenesis induction is an effective way to obtain tissue-cultured plants for the species that are recalcitrant to somatic embryo formation. However, the induction of somatic embryogenesis is quite complex; thus, there are many factors, such as the genetic background and DNA methylation level of the mother plant, the type of explant, and the level of plant growth regulators during induction and culture, that affect the success rate of somatic embryogenesis [16,17]. Plant growth regulators play important roles in regulating cell growth, division, and differentiation during in vitro plant tissue culture. For example, auxin is usually used for eliciting plant rooting and vegetative propagation from stem and leaf cuttings, and cytokinin is necessary for promoting somatic embryogenesis [18,19,20,21]. However, since plant tissues cannot produce auxin and cytokinin under in vitro culture conditions, exogenous supplementations of auxin and cytokinin are critically required for ensuring plant embryogenic callus induction and somatic embryogenesis [22,23,24]. In pomegranate, despite some successful applications of somatic embryogenesis, its recalcitrant nature poses challenges to the somatic embryo induction for the major reason that the existing protocols mainly employ cotyledonary or leaf explants; however, the low availability of cotyledonary tissues limits the widespread commercial application, while the leaf-derived materials are prone to browning, leading to tissue culture failure [25]. Thus, a stable and high-yield somatic embryogenesis system is urgently required for pomegranate.
To address the abovementioned problem, in this study, we aim to establish an effective somatic embryogenesis system for pomegranate using easily acquired explants. Additionally, we further explored the optimal culture conditions for the induction and development of somatic embryos to improve the efficiency of somatic embryogenesis in pomegranate. Our system offers a solution to bridge the difficulties of mass somatic embryo induction and plant regeneration in pomegranate and also provides a foundation for the future genetic transformation and bioengineering improvement of pomegranate with desirable traits.
2. Materials and Methods
2.1. Cultivation of Tissue Culture Plantlets
The pomegranate plantlets for tissue culture were produced following the methods described in Qian et al. (2022) [26]. Briefly, in the spring of 2022 in Huaiyuan, Anhui Province, 2–3 cm shoot tips were cut off from the tender stems of thriving and disease-free plants of the pomegranate variety ‘White Flower Jade Seeds’. After cultivation in water for 2–3 days, the growth points of the excised shoot tips were collected and rinsed in a beaker with clean water for 15 min. Then, they were disinfected with 75% alcohol for 30 s, washed with sterile water 3 times, disinfected with 0.1% HgCl2 for 8 min, and washed with sterile water 4 times. Tissue sections of ~0.3–0.5 mm were collected from the sterilized materials and inoculated onto a woody plant medium (WPM) supplemented with 0.8 mg/L indole-3-butyric acid (IBA, chemical formula: C12H13NO2), 30 g/L sucrose, and 6 g/L agar. The pH of the medium was adjusted to 6.5 with 1 M NaOH and sterilized at 120 °C and 105 kPa for 20 min.
2.2. Screening Optimum Combination of Media and Plant Growth Regulators for Callus Induction
The pomegranate tissue culture plantlets were then subcultured 3 times, and plantlets with similar growth conditions were selected and cut into ~0.5–0.8 cm length stem segments without bud points or leaves on the ultraclean workbench. The explants were inoculated horizontally on the induction medium. Under in vitro conditions, the main factors affecting callus formation include the types of explants, medium, and concentrations of exogenous plant growth regulators [27]. In the current study, we tested the effects of different combinations of media (WPM or MS medium) and plant growth regulators (1.0 mg/L 6-BA (C12H11N5), 1.0 mg/L NAA (C12H10O2), and 0.5 mg/L IBA) on callus induction to determine the appropriate culture conditions for inducing pomegranate callus [25]. For each combination, 30 biological replicates were performed. The culture was maintained at a constant temperature of 22 ± 2 °C and illumination intensity of 1600 xl by LED lights at a 16/8 h light/dark cycle. After 30 days, the callus induction status was recorded, and the callus induction rate was calculated as follows: callus induction rate = number of calli induced from explants/number of explants inoculated × 100%. Factorial Analysis of Variance (ANOVA) was performed to assess the statistical significance level among 6 different combinations of basal medium and plant growth regulators (see Table 1 for details) in SPSS Statistics v. 17.0 (SPSS Inc., Chicago, IL, USA). Duncan’s Multiple Range test (DMRT) was then conducted to measure the specific difference between each pair of culture conditions.

Table 1.
Induction rate and growth status of calli induced by different combinations of media and plant growth regulators.
2.3. Screening Optimum Concentrations of Plant Growth Regulators for Embryogenic Callus Induction
Pomegranate embryogenic calli were then induced using the calli we obtained from the last step. In general, one key step for embryogenic callus induction is to screen suitable culture conditions, particularly the concentration of exogenous plant growth regulators. Here, we used MS medium as the basal medium and supplemented it with 6-BA and NAA at different concentrations (0.5, 1.0, and 1.5 mg/L for both types of plant growth regulators) to obtain the optimum combination of plant growth regulators for pomegranate embryogenic callus induction. Each combination was examined with 30 biological replicates. After culturing for 30 days, the growth state of the embryogenic calli was recorded and photographed using a Leica M205 (Leica Microsystems Inc., Buffalo Grove, IL, USA), and the induction rate of embryogenic calli was measured as follows: embryogenic callus induction rate = number of embryogenic calli induced from explants/number of inoculated explants × 100%. Statistical differences among 9 combinations of different 6-BA and NAA concentrations (detailed in Table 2) were measured by two-way ANOVA using SPSS software. Multiple comparison procedure was performed by the post hoc DMRT.
Table 2.
Induction rate and growth status of embryogenic calli induced by the media with different types and concentrations of plant growth regulators.
2.4. Evaluation of Different Explants on Embryogenic Callus Induction
We further compared the performance of using stem segments and hypocotyls as explants on embryogenic callus induction. The hypocotyls were from pomegranate seedlings that germinated and grew for 7 days. All the explants were inoculated onto MS medium supplemented with 1.0 mg/L NAA and 0.5 mg/L 6-BA, according to the screening results (detailed in Section 3.2). For each type of explant, the experiments were repeated 105 times, with explants inoculated on 15 Petri dishes and each dish containing 7 replicates. The embryogenic callus induction rate was then calculated by the equation described above. The difference in induction of embryogenic calli between the two types of explants was evaluated by one-way ANOVA in SPSS software.
2.5. Evaluation of Different Subculture Cycles on Proliferation of Embryogenic Calli
We also examined the impacts of the different subculture cycles on the proliferation of embryogenic calli. Well-grown embryogenic calli were selected and inoculated on MS medium with 0.5 mg/L 6-BA and 1.0 mg/L NAA to produce new embryogenic calli. The embryogenic calli were weighed immediately after inoculation to serve as the fresh weight of the initial calli and were then weighed and recorded every 2 days until the fresh weight no longer increased. Origin 2021 (OriginLab Corp., Northampton, MA, USA) was used to fit the growth curve.
2.6. Screening Optimum Concentrations of Plant Growth Regulators for Somatic Embryogenesis
The embryogenic calli of good growth status were transferred to the culture medium for somatic embryo induction. Similar to embryogenic calli, the induction of somatic embryos is also sensitive to the concentrations of plant growth regulators in the medium. Thus, in this study, we tested the impacts of different 6-BA and NAA concentrations on somatic embryogenesis. Specifically, the MS medium was supplemented with 0.1, 0.5, and 1.0 mg/L 6-BA and NAA, and the embryogenic calli were then inoculated. Each culture condition was examined with 75 biological replicates. The growth status of the somatic embryos was recorded and photographed using a Leica M205. The somatic embryo induction rate was computed after 35 days of growth with the following formula: somatic embryo induction rate = number of somatic embryos induced from calli/number of calli inoculated × 100%. Two-way ANOVA was implemented to quantify the statistically significant level among the nine combinations of different 6-BA and NAA concentrations (detailed in Table 3), and DMRT was employed to measure the pairwise differences between culture conditions.
Table 3.
Induction rate and growth status of somatic embryos induced by the media with different concentrations of plant growth regulators.
2.7. Rooting and Regeneration of Somatic Embryo Seedlings
The somatic embryo plantlets were inoculated on WPM supplemented with 0.6 mg/L IBA, 5.5 g/L agar, and 25 g/L sucrose (rooting medium) for rooting and growth. The somatic embryo seedlings were washed and transplanted onto the seedling medium in a 6.5 × 6.5 cm nutrient bowl for cultivation. The seedling medium was a mixture of nutritive soil and vermiculite in a ratio of 1:1. Before use, the seedling medium was disinfected at a high temperature and was sprayed with 50 mL 0.8 mg/L IBA. After the transplantation, the nutrient bowl was covered by plastic film, and the medium was sprayed with water every morning and evening to keep the substrate moist. Air humidity was kept above 50%. Every 5 days, the seedings were sprayed with Hoagland’s nutrient solution until they were viable (~25 days). After taking roots in the medium, the somatic embryo seedlings were washed and transplanted onto the seedling medium in a 6.5 × 6.5 cm nutrient bowl for cultivation. The seedling medium was a mixture of nutritive soil and vermiculite in a ratio of 1:1. Before use, the seedling medium was disinfected at a high temperature, and was sprayed with 50 mL 0.8 mg/L IBA. After the transplantation, the nutrient bowl was covered by plastic films, and the medium was sprayed with water every morning and evening to keep the substrate moist. Air humidity was kept above 50%. EFor every 5 days, the seedings were was sprayed with Hoagland’s nutrient solution until they were viable (~25 days). After that, the root-bearing seedlings were transplanted to the natural environment, and their growth status was recorded.
3. Results
3.1. Effects of Different Combinations of Media and Plant Growth Regulators on Callus Induction
Table 1 shows the induction percentage and growth status of calli formed from pomegranate stem segments growing under different combinations of media and plant growth regulators. For all the growth scenarios, more than 60% of the explants were successfully induced into calli. The explants grown on WPM supplemented with 6-BA and IBA (medium A2) exhibited the highest induction percentage of calli (81.11 ± 5.09%), which was significantly higher than that in other scenarios (p < 0.05). However, on this medium, the calli were light brown and dense and were prone to browning and necrosis over time. In contrast, the calli on the WPM with IBA and NAA (medium A3) and on the MS medium with IBA and NAA (medium A6) exhibited the best growth status in that they were mostly gray-white and not dense but had a relatively low induction rate. Taken together, these results suggested that MS medium supplemented with 1.0 mg/L 6-BA and 1.0 mg/L NAA (medium A4) was the optimal culture condition for callus induction from stem explants and was, therefore, used for subsequent experiments.
3.2. Effects of Different Concentrations of Plant Growth Regulators on Embryogenic Callus Induction
We then tested the impacts of different 6-BA and NAA concentrations on embryogenic callus induction and found that both the induction rate and growth status of embryogenic calli were sensitive to changes in the concentrations of both plant growth regulators (Figure 1; Table 2). Either too high or too low a concentration of NAA in the medium led to a dramatic decrease in the induction rate. On the medium with 1.5 mg/L NAA (media B3, B6 and B9), less than 30% of the explants were induced into embryogenic calli, and they were yellow to brown and did not grow well. In contrast, the medium with 0.5 or 1 mg/L 6-BA was more conducive to the induction of pomegranate embryogenic calli than that with 1.5 mg/L 6-BA. Among the nine combinations, the medium supplemented with 0.5 mg/L 6-BA and 1 mg/L NAA (B2) was most suitable for the induction of pomegranate embryogenic calli, where the induction rate (74.44 ± 5.09%) was significantly higher than all the other culture conditions (p < 0.05), and the produced embryogenic calli were light yellow to light green, fast-growing and not dense (Figure 1 and Figure 2; Table 2). Thus, this medium was used for inducing embryogenic calli from pomegranate stem segments.

Figure 1.
Pomegranate embryogenic callus induced by the media with different concentrations of plant growth regulators.

Figure 2.
Pomegranate embryogenic calli induced using medium B2 (see Table 2 for details). Red arrow points to a magnified shot of the induced embryogenic calli.
3.3. Effect of Different Explants on Embryogenic Callus Induction
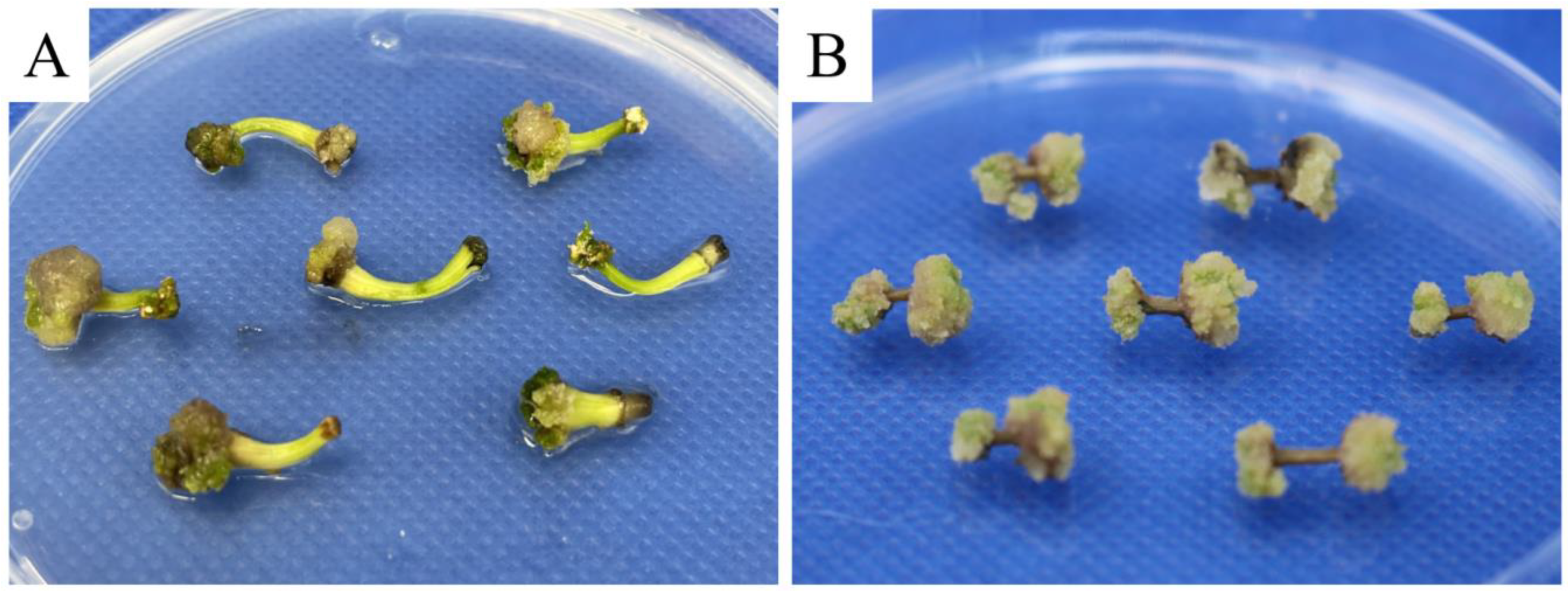
We also examined the performance of different explants on embryogenic callus induction. When using hypocotyl explants, 72.38 ± 4.36% of the inoculated explants were successfully induced into embryogenic calli. However, after 37 days of culture, the growth of induced calli slowed and some calli began to turn brown and necrose (Figure 3A). In contrast, the induction using stem segments performed better than that using hypocotyls, with a significantly higher induction rate of embryogenic calli (80.00 ± 4.95%; p < 0.05). In addition, no obvious stagnation was observed in their growth rate on the 35th day (Figure 3B); thus, stem segments were more suitable than hypocotyl explants for pomegranate somatic embryogenesis.
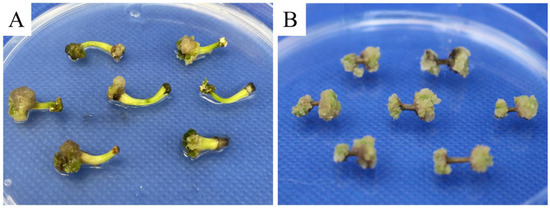
Figure 3.
Pomegranate embryogenic calli induced by hypocotyl (A) and stem explants (B).
3.4. Effect of Subculture Cycle on Embryogenic Callus Proliferation
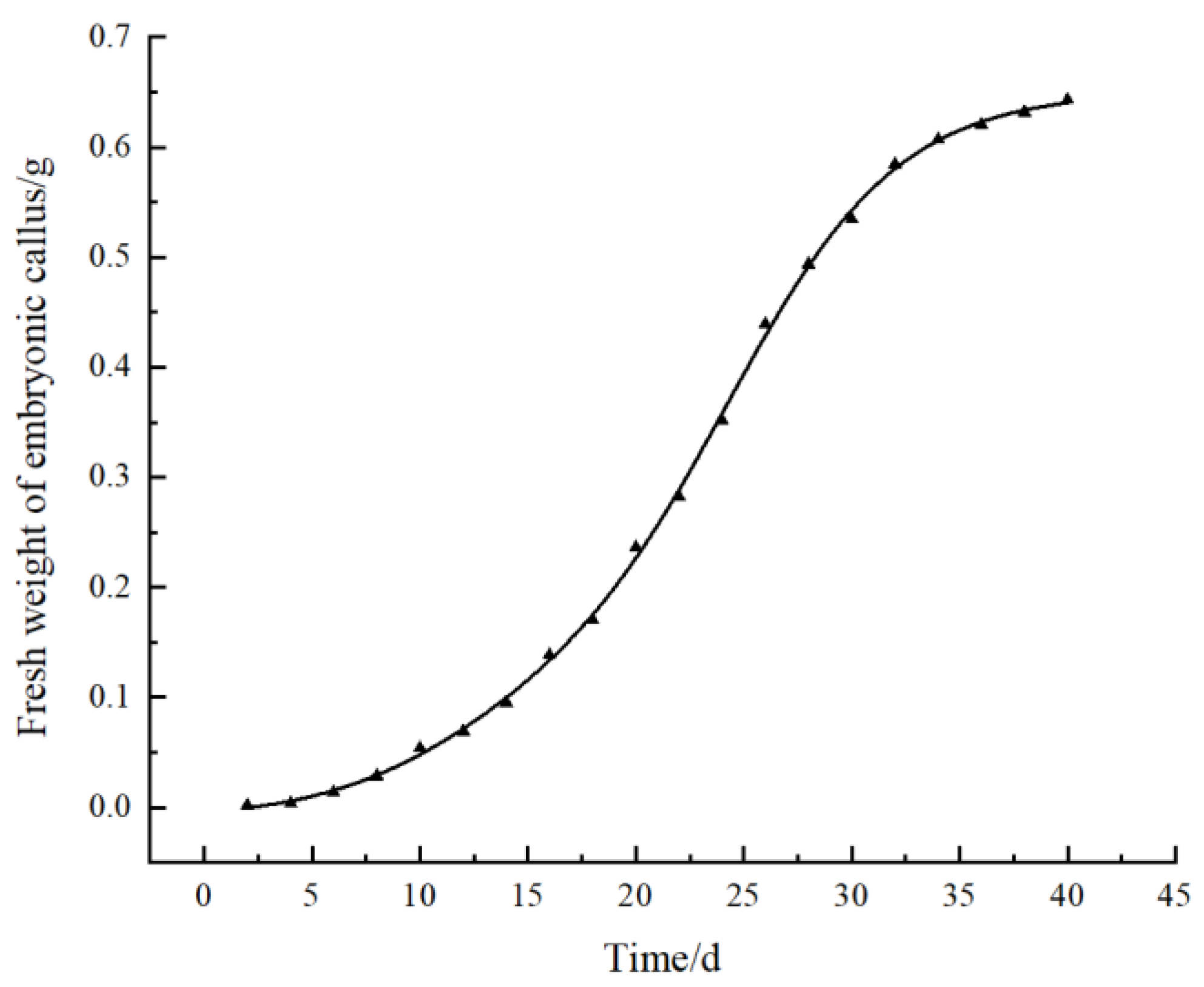
The obtained well-grown embryogenic calli were transferred to fresh B2 medium to maintain embryogenicity and proliferation. Then, we drew the growth curve according to the measurement data from the fresh weight of the embryogenic calli (Figure 4). The growth curve showed that the fresh weight of the embryogenic calli continuously increased over time. The callus proliferation rate increased after 15 days of inoculation and entered the rapid growth period at the 20th to 30th days. During this period, the color of the calli was light green. After 30 days, the growth rate began to slow down, and the tissue gradually degenerated, the color gradually turned yellow, and the granular texture softened. From the 40th to 55th days, almost all calli died of browning. Therefore, a suitable subculture cycle for embryogenic calli was determined to be 30–35 days to effectively maintain callus quality.
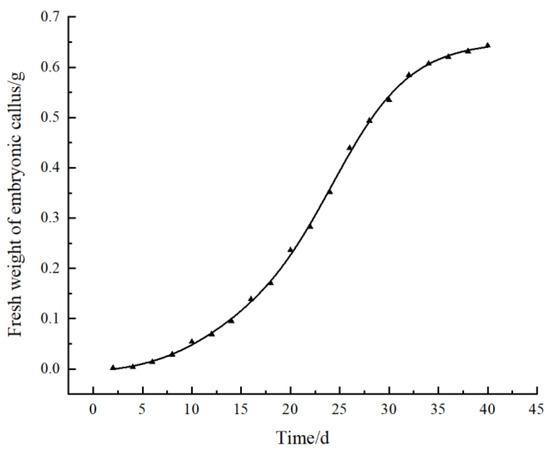
Figure 4.
Growth curve of pomegranate embryonic calli.
3.5. Effects of Different Concentrations of 6-BA and NAA on Somatic Embryo Induction
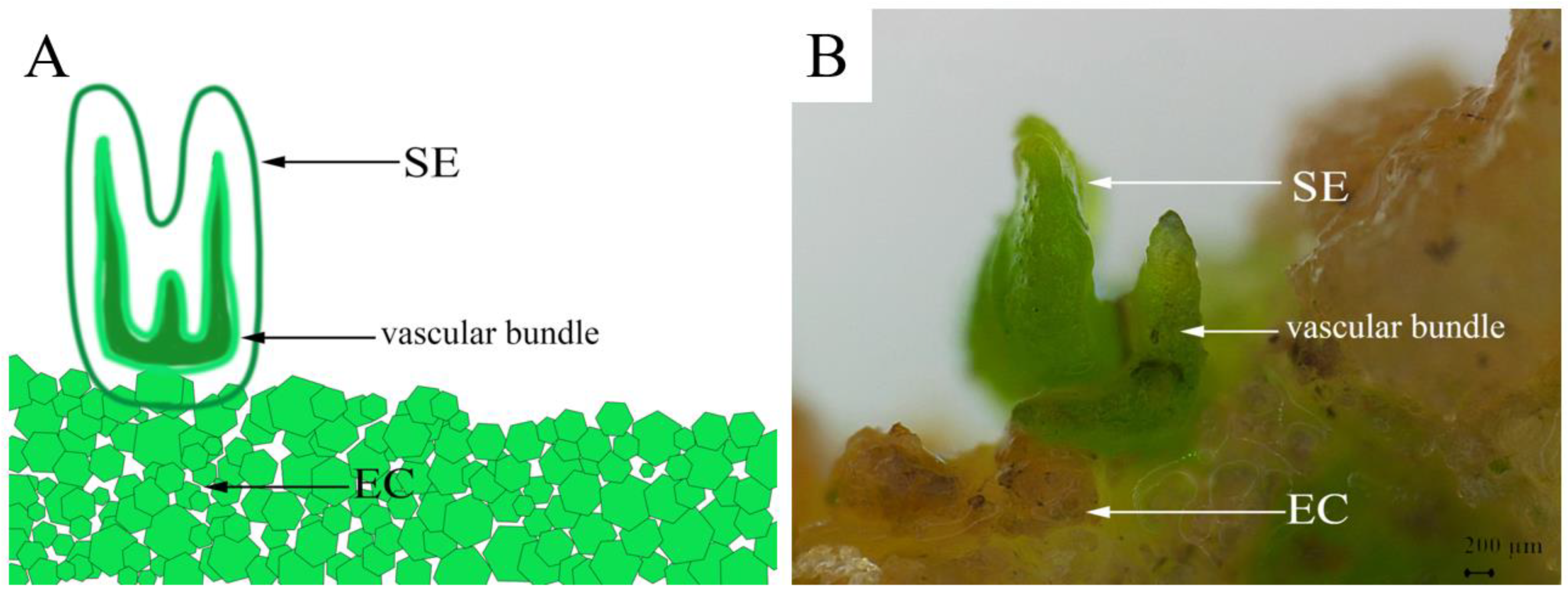
In addition to embryogenic calli, 6-BA and NAA concentrations in the MS medium were also found to substantially affect the induction of somatic embryos. Among all the tested culture conditions, only C5, C6, and C8 exhibited somatic embryo induction rates higher than 50% (Table 3). Of them, the MS medium with 0.5 mg/L 6-BA and 0.5 mg/L NAA (medium C5) was the most suitable for somatic embryo induction, with 78.67 ± 6.11% of the calli induced into somatic embryos after being inoculated for 25 days, which was a significantly higher rate than in the others (p < 0.05). Meanwhile, granular proembryonic masses were observed on the surface of the embryogenic calli after 50 days of culture (Figure 5B). As the culture time was extended, the embryos, in turn, appeared globular and heart-shaped/torpedo-shaped in form (Figure 5C–E), and eventually developed into cotyledon embryos by approximately 50 days (Figure 5F). Under the stereomicroscope, we observed plumules at one end of the cotyledon embryos and the radicles at the other end that were connected to the callus (Figure 6). In addition, we also observed the apparent structure of the vascular bundle in the cotyledon, which was independent of the embryogenic calli.

Figure 5.
Embryogenic calli and different somatic embryo stages of pomegranate. (A) Calli induced by stem explants. (B) Embryogenic calli (C) Globular embryo. (D) Extended globular embryo. (E) Heart-shaped/torpedo-shaped embryo. (F) Cotyledon embryo. In each of the panel (B–F), the induced somatic embryos was marked by red arrow.
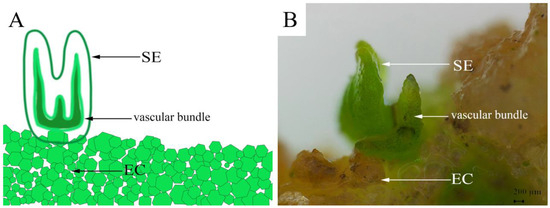
Figure 6.
Schematic (A) and microscopic views (B) of pomegranate somatic embryogenesis. SE, somatic embryo; EC, embryonic callus.
3.6. Rooting and Regeneration of Somatic Embryo Seedlings
The embryogenic calli were inoculated on the medium C5 for 45 days, and the mature somatic embryos developed complete seedlings (Figure 7A,B). The somatic embryo seedlings were transferred to the natural environment to form regenerated plants (Figure 7C,D). During this process, radicle formation is a key step that determines whether the production of somatic embryo seedlings can be industrialized and commercialized. On the rooting medium, all the somatic embryo seedlings grew normally and generated healthy roots (Figure 7C,D). Then, these root-bearing seedlings were developed into healthy and vigorous plants in the natural environment (Figure 7E,F).

Figure 7.
Somatic embryo seedlings of pomegranate. (A,B) Mature somatic embryos with buds. (C) Somatic embryo seedlings inoculated onto rooting medium. (D) Roots of somatic embryo seedlings in culture medium. (E,F) Somatic embryo seedlings after seedling refinement and transplantation.
4. Discussion
Somatic embryogenesis faces many difficulties and problems in woody plants that are not encountered with herbaceous plants. In this study, we first established a protocol for the effective induction of somatic embryogenesis in pomegranate using stem explants. In this system, we employed an indirect organogenesis strategy for somatic embryogenesis induction, that involved first generating embryogenic calli from dedifferentiated pomegranate explant cells as an intermediate stage and then inducing somatic embryos from the produced calli. The results showed that with our system, we successfully induced pomegranate embryogenic calli from stem segments (Figure 2 and Figure 3). Under the optimal combination of plant growth regulators, the granular proembryo mass, as well as somatic embryos at various stages, was then elicited at the surface of the calli, and the mature cotyledonary somatic embryos could develop into healthy somatic seedlings (Figure 5 and Figure 7). After the somatic embryo seedlings were transferred to the rooting medium, all the pomegranate seedlings developed healthy roots within 20 days, indicating their good rooting ability both in vivo and in vitro (Figure 7).
During this process, obtaining high-quality embryogenic calli is a vital step [12]. Here, we first explored the effects of different combinations of two types of media, WPM and MS, and three types of plant growth regulators on pomegranate callus induction. The results showed that the induction rate of calli when using MS medium was higher than that when using WPM. This may be because of their differences in the composition of nutrients and elements [28]. In addition to the basal medium, plant growth regulators also play a critical role in regulating plant somatic embryogenesis. For pomegranate, exogenous auxin and cytokinin are generally supplemented to the culture medium to improve embryogenesis induction and shoot proliferation [25,29,30]. In this study, we assessed the performance of the combination of different types of plant growth regulators 6-BA, NAA, and IBA in pomegranate somatic embryogenesis and showed that supplementing the MS medium with 6-BA and NAA achieved the best balance between callus induction rate and growth status (Table 1). NAA is a synthetic auxin that is usually used for eliciting plant rooting and vegetative propagation from stem and leaf cuttings [18,19], and 6-BA is a plant growth regulator of the cytokinin family that can stimulate plant growth and development. However, for both types of plant growth regulators, high concentrations in the medium were found to have adverse impacts on the callus induction and growth. In our system, the appropriate 6-BA concentration for induction of pomegranate embryogenic calli and somatic embryos is 0.5 mg/L, and the optimum NAA concentration is 1.0 and 0.5 mg/L, respectively, where the induction rates of embryogenic calli and somatic embryos were ~74% and 79%, respectively (Table 2 and Table 3). This is because auxins at high concentrations are toxic to plants, while a high cytokinin concentration inhibits auxin polar transport and, thus, inhibits somatic embryogenesis [21,30,31]. However, different combinations of plant growth regulators are required for in vitro propagation of pomegranate when using different explants [25]. For instance, Kantharajah et al. [32] showed that MS medium supplemented with 1.0 mg/L 6-BA and 0.4 mg/L NAA gave rise to the best callus initiation and growth from nodal explants, while the medium containing 1.0 mg/L 6-BA achieved the best performance for leaf explant-derived callus culture. These results indicate that the choice of plant growth regulators used in a nutrient medium and their ratio (auxin/cytokinin) are largely dependent on the morphogenic potentials of the pomegranate explants employed for in vitro culture.
In general, all types of plant tissues with strong meristematic ability can be used as explants for tissue culture, but different explants have different abilities in forming calli [33]. Jaidka and Mehra [34] demonstrated the success of using pomegranate stem explants in inducing calli when cultured in MS medium containing 4.0 mg/L NAA, 2.0 mg/L kinetin, and 15% coconut water, indicating the feasibility of using stem explants for pomegranate somatic embryogenesis. In addition, Yang and Ludders [35] reported organogenesis in pomegranate initiated from stem explants. In the current study, we first induced pomegranate somatic embryogenesis using stem segments as explants and showed better performance using stem explants than using hypocotyls in producing pomegranate calli in terms of both induction rate and the growth status of embryogenic calli. Although pomegranate calli and somatic embryos can also be generated using other types of explants, such as leaf, shoot tip, nodal segment, and cotyledonary tissue, these explants exhibit their own shortcomings: shoot tips are not able to provide enough materials, and the materials from leaves and leafstalk are prone to browning, which leads to tissue culture failure [25]. Comparatively, stem segments are more easily assessable, and the embryogenic calli formed by stem segments did not show obvious browning and grew well. These advantages make stem explants more suitable for initial culture, especially for industrial and commercial production.
The callus subculture cycle also plays a key role in eliciting plant somatic embryos, and an appropriate number of subculture cycles can ensure the good growth status of embryogenic calli [36]. The pomegranate embryogenic calli inoculated for 20–30 days entered the rapid growth period, during which the callus proliferation rate was the fastest, and its color was brightly light green. The calli turned brown after 40 days of culture, and almost all died after 55 days, indicating that they had lost their capacity for somatic embryogenesis. Therefore, taking 30–35 days as the subculture cycle is more suitable for long-term subculture of pomegranate calli and maintenance of somatic embryogenesis.
5. Conclusions
In this study, we established an effective system for somatic embryogenesis and plant regeneration in pomegranate using stem segments as explants. In our system, the MS medium containing 1.0 mg/L 6-BA and 1.0 mg/L NAA achieved the best performance in pomegranate callus induction, and the MS medium with 0.5 mg/L 6-BA and 1.0 and 0.5 mg/L NAA was the optimal condition for the induction of embryogenic calli and somatic embryos, where the induction rates of calli, embryogenic calli, and somatic embryos were ~72%, 74%, and 79%, respectively. Good somatic embryo induction indicates the effectiveness and stability of our system. In addition, compared to the existing systems, the stem explant employed in our protocol is more easily acquired and suitable for widespread commercial production. Taken together, our system overcomes the recalcitrant nature of pomegranate and provides a solution to mass somatic embryo induction and plant regeneration of pomegranate. With the help of this somatic embryogenesis protocol, we aim to build a genetic transformation system for future bioengineering improvement of pomegranate with favorable agronomic traits.
Author Contributions
Conceptualization, G.C. and J.Q.; validation, J.W. (Jingting Wang) and J.Q.; formal analysis, J.W. (Jingting Wang); investigation, J.W. (Jingting Wang), X.X., J.T., J.W. (Jun Wang), W.Z., M.Q. and J.L.; writing—original draft preparation, J.W. (Jingting Wang), X.X., Y.Y. and J.Q.; writing—review and editing, X.X., Y.Y., G.Q. and J.Q.; visualization, J.W.(Jingting Wang) and J.Q; supervision, G.C., Y.Y. and J.Q.; project administration, G.C., Y.Y. and J.Q.; funding acquisition, J.Q., Y.Y. and G.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Anhui Province Key Research and Development Plan Project, grant number 202204c06020062; Project of Industry and School and Research Institution, grant number AKZY2022110; Anhui Province Natural Sciences Fund, grant number 2023AH040281; National Natural Science Foundation of China, grant number 32201420; Natural Science Foundation of Anhui Province, grant number 2308085MC95.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caruso, A.; Barbarossa, A.; Tassone, A.; Ceramella, J.; Carocci, A.; Catalano, A.; Basile, G.; Fazio, A.; Iacopetta, D.; Franchini, C.; et al. Pomegranate: Nutraceutical with Promising Benefits on Human Health. Appl. Sci. 2020, 10, 6915. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, X.; Ni, H.; Yuan, F.; Zhang, S. Identification and Functional Analysis of CAD Gene Family in Pomegranate (Punica granatum). Genes 2023, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, J.; Qin, G. Genome-wide distribution of simple sequence repeats in pomegranate and their application to the analysis of genetic diversity. Tree Genet. Genom. 2020, 16, 36. [Google Scholar] [CrossRef]
- Xia, X.; Fan, M.; Liu, Y.; Chang, X.; Wang, J.; Qian, J.; Yang, Y. Genome-wide alternative polyadenylation dynamics underlying plant growth retardant-induced dwarfing of pomegranate. Front. Plant Sci. 2023, 14, 1189456. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, S.; Qin, C.; Li, Q.; Cai, J.; Sun, C.; Wang, W.; Weng, Y. Somatic embryogenesis in mature zygotic embryos of Picea pungens. Sci. Rep. 2021, 11, 19072. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romero, C. Somatic embryogenesis in Olea spp. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 138, 403–426. [Google Scholar] [CrossRef]
- Park, Y.-S. Conifer Somatic Embryogenesis and Multi-Varietal Forestry. In Challenges and Opportunities for the World’s Forests in the 21st Century; Fenning, T., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 425–439. [Google Scholar] [CrossRef]
- Corredoira, E.; Ballester, A.; Ibarra, M.; Vieitez, A.M. Induction of somatic embryogenesis in explants of shoot cultures established from adult Eucalyptus globulus and E. saligna × E. maidenii trees. Tree Physiol. 2015, 35, 678–690. [Google Scholar] [CrossRef]
- Pais, M.S. Somatic Embryogenesis Induction in Woody Species: The Future After OMICs Data Assessment. Front. Plant Sci. 2019, 10, 240. [Google Scholar] [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, C.; Wang, H.; Ma, X.; Shen, H.; Yang, L. Indirect somatic embryogenesis and regeneration of Fraxinus mandshurica plants via callus tissue. J. For. Res. 2021, 32, 1613–1625. [Google Scholar] [CrossRef]
- Yan, R.; Sun, Y.; Sun, H. Current status and future perspectives of somatic embryogenesis in Lilium. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 143, 229–240. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.-G.; Fan, X.-F.; Su, Z.-H. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef]
- Baskaran, P.; Van Staden, J. Plant regeneration via somatic embryogenesis in Drimia robusta. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 119, 281–288. [Google Scholar] [CrossRef]
- Khan, T.; Reddy, V.S.; Leelavathi, S. High-frequency regeneration via somatic embryogenesis of an elite recalcitrant cotton genotype (Gossypium hirsutum L.) and efficient Agrobacterium-mediated transformation. Plant Cell Tissue Organ Cult. (PCTOC) 2010, 101, 323–330. [Google Scholar] [CrossRef]
- Lu, L.; Holt, A.; Chen, X.; Liu, Y.; Knauer, S.; Tucker, E.J.; Sarkar, A.K.; Hao, Z.; Roodbarkelari, F.; Shi, J.; et al. miR394 enhances WUSCHEL-induced somatic embryogenesis in Arabidopsis thaliana. New Phytol. 2023, 238, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Syeed, R.; Mujib, A.; Malik, M.Q.; Gulzar, B.; Zafar, N.; Mamgain, J.; Ejaz, B. Direct somatic embryogenesis and flow cytometric assessment of ploidy stability in regenerants of Caladium × hortulanum ‘Fancy’. J. Appl. Genet. 2022, 63, 199–211. [Google Scholar] [CrossRef]
- Xiong, Y.; Wei, Z.; Yu, X.; Pang, J.; Zhang, T.; Wu, K.; Ren, H.; Jian, S.; Teixeira da Silva, J.A.; Ma, G. Shoot proliferation, embryogenic callus induction, and plant regeneration in Lepturus repens (G. Forst.) R. Br. Vitr. Cell. Dev. Biol. Plant 2021, 57, 1031–1039. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, F.; Wang, Y.; Li, X.; Liang, H. Efficient Somatic Embryogenesis and Plant Regeneration from Immature Embryos of Tapiscia sinensis Oliv., an Endemic and Endangered Species in China. HortScience Horts 2014, 49, 1558–1562. [Google Scholar] [CrossRef]
- Bernula, D.; Benkő, P.; Kaszler, N.; Domonkos, I.; Valkai, I.; Szőllősi, R.; Ferenc, G.; Ayaydin, F.; Fehér, A.; Gémes, K. Timely removal of exogenous cytokinin and the prevention of auxin transport from the shoot to the root affect the regeneration potential of Arabidopsis roots. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 327–339. [Google Scholar] [CrossRef]
- Binte Mostafiz, S.; Wagiran, A. Efficient Callus Induction and Regeneration in Selected Indica Rice. Agronomy 2018, 8, 77. [Google Scholar] [CrossRef]
- Ming, N.J.; Binte Mostafiz, S.; Johon, N.S.; Abdullah Zulkifli, N.S.; Wagiran, A. Combination of Plant Growth Regulators, Maltose, and Partial Desiccation Treatment Enhance Somatic Embryogenesis in Selected Malaysian Rice Cultivar. Plants 2019, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.-W.; Yan, Y.-Y.; Liu, Y.-M.; Liu, Y.-M.; Deng, Y.-W.; Yao, H.-Y. The regeneration of Acer rubrum L. “October Glory” through embryonic callus. BMC Plant Biol. 2020, 20, 309. [Google Scholar] [CrossRef] [PubMed]
- Teixeira da Silva, J.A.; Rana, T.S.; Narzary, D.; Verma, N.; Meshram, D.T.; Ranade, S.A. Pomegranate biology and biotechnology: A review. Sci. Hortic. 2013, 160, 85–107. [Google Scholar] [CrossRef]
- Qian, J.; Wang, N.; Ren, W.; Zhang, R.; Hong, X.; Chen, L.; Zhang, K.; Shu, Y.; Hu, N.; Yang, Y. Molecular Dissection Unveiling Dwarfing Effects of Plant Growth Retardants on Pomegranate. Front. Plant Sci. 2022, 13, 866193. [Google Scholar] [CrossRef] [PubMed]
- Shin, U.; Chandra, R.; Kang, H. In vitro and Ex vitro Propagations of Astilboides tabularis (Hemsl.) Engl. as a Rare and Endangered Species. Hortic. J. 2019, 6, 261. [Google Scholar]
- Hazrati, R.; Zare, N.; Asghari-Zakaria, R.; Sheikhzadeh, P.; Johari-Ahar, M. Factors affecting the growth, antioxidant potential, and secondary metabolites production in hazel callus cultures. AMB Express 2022, 12, 109. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors Influencing Somatic Embryogenesis Induction and Plant Regeneration with Particular Reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Fehér, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, Q.; Kita, D.; Huang, J.-B.; Liu, G.; Wu, X.; Zhu, X.; Cheung, A.Y.; Wu, H.-M.; Tao, L.-Z. RopGEF1 Plays a Critical Role in Polar Auxin Transport in Early Development. Plant Physiol. 2017, 175, 157–171. [Google Scholar] [CrossRef]
- Kantharajah, A.; Dewitz, I.; Jabbari, S. The effect of media, plant growth regulators and source of explants on in vitro culture of pomegranate (Punica granatum L.). Erwerbsobstbau 1998, 40, 54–58. [Google Scholar]
- Vinterhalter, B.; Mitić, N.; Vinterhalter, D.; Uzelac, B.; Krstić-Milošević, D. Somatic embryogenesis and in vitro shoot propagation of Gentianautriculosa. Biologia 2016, 71, 139–148. [Google Scholar] [CrossRef]
- Jaidka, K.; Mehra, P.N. Morphogenesis in Punica granatum (pomegranate). Can. J. Bot. 1986, 64, 1644–1653. [Google Scholar] [CrossRef]
- Yang, Z.; Ludders, P. Organogenesis of Punica granatum L. var. nana. Angew. Bot. 1993, 67, 151–156. [Google Scholar]
- Ren, Y.; Yu, X.; Xing, H.; Tretyakova, I.N.; Nosov, A.M.; Yang, L.; Shen, H. Interaction of Subculture Cycle, Hormone Ratio, and Carbon Source Regulates Embryonic Differentiation of Somatic Cells in Pinus koraiensis. Forests 2022, 13, 1557. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).