
Harnessing Gut Microbiota for Biomimetic Innovations in Health and Biotechnology



{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. The Importance of the Intestinal Microbiota in Human Health
1.2. Concept of Biomimetics: Definition and Relevance to Medical Care
2. Materials and Methods
3. Gut Microbiota: A Complex Biological Ecosystem
3.1. Composition and Main Functions of the Gut Microbiota
3.2. Microbiota–Host Interaction: Immune Regulation, Metabolism, and Communication
3.3. Production of Key Metabolites: Short-Chain Fatty Acids, Indoles, and Neurotransmitters
4. Biomimetics Applied to Gut Microbiota
4.1. Bioinspired Concepts Derived from the Microbiota
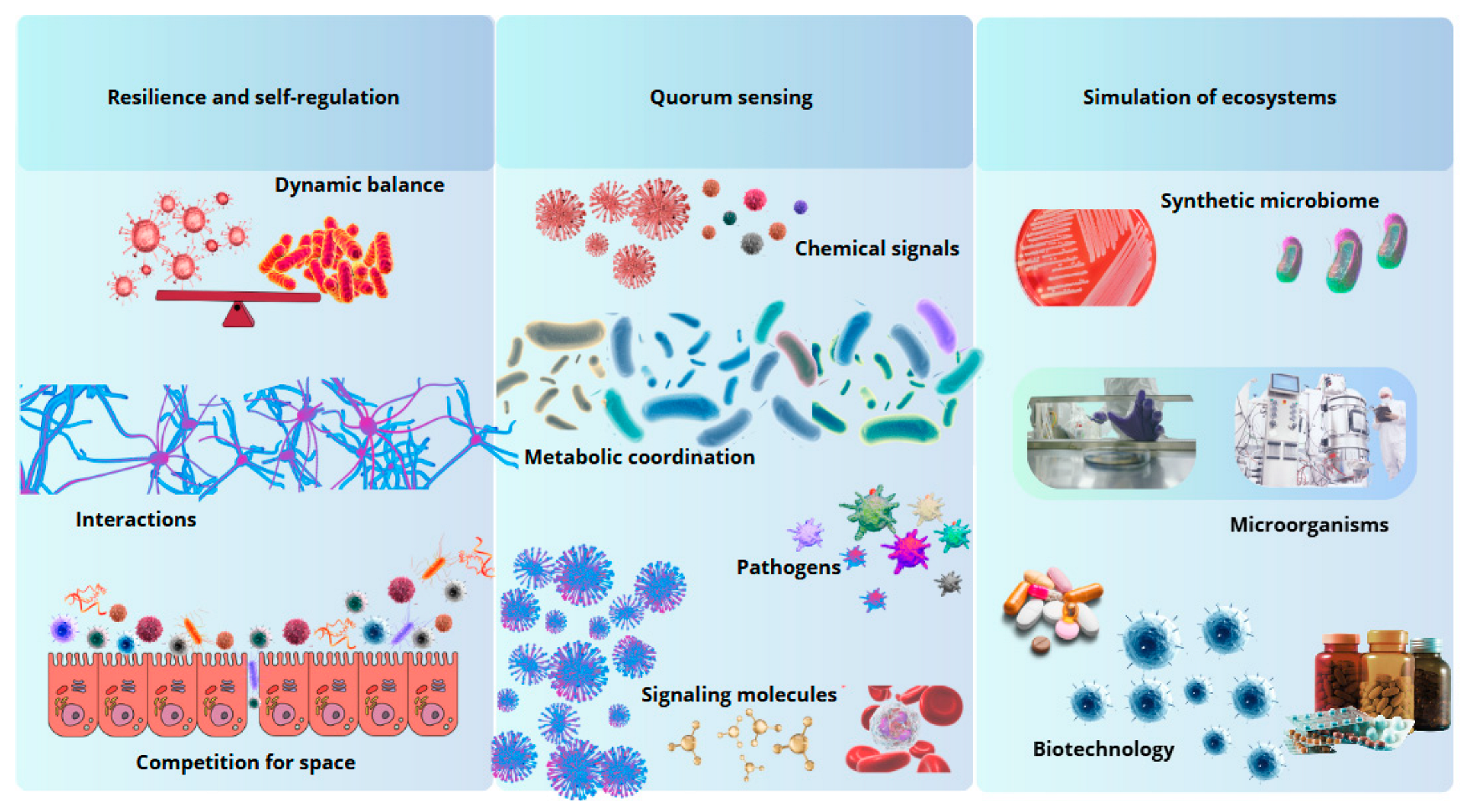
4.1.1. Resilience and Self-Regulation of Microbial Ecosystems
4.1.2. Microbial Intercellular Communication (Quorum Sensing)
4.2. Simulate Microbial Ecosystems in Artificial Environments
5. Innovations Inspired by the Intestinal Microbiota
5.1. Therapies Based on Probiotics and Synbiotics
5.1.1. The Development of Probiotics Inspired by Biological Processes
5.1.2. Synbiotics Designed to Replicate Specific Functions of the Microbiota
5.2. Artificial Microbiomes and Bioartificial Systems
5.2.1. Artificial Models for Simulating Host–Microbiota Interactions
5.2.2. Applications in Fecal Microbiota Transplants
5.3. Bioinspired Devices and Biosensors
5.3.1. Diagnostic Tools Based on Functional Patterns of the Microbiota
5.3.2. Sensors That Detect Microbial Metabolites in Real Time
6. Clinical Applications and Challenges
7. Prospective Trends in Microbial Biomimetics
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.A.A.A.; Weersma, R.K.; Vila, A.V. The emerging role of the small intestinal microbiota in human health and disease. Gut Microbes 2023, 15, 2201155. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar]
- Clemente-Suárez, V.J.; Rubio-Zarapuz, A.; Belinchón-deMiguel, P.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. Impact of Physical Activity on Cellular Metabolism Across Both Neurodegenerative and General Neurological Conditions: A Narrative Review. Cells 2024, 13, 1940. [Google Scholar] [CrossRef] [PubMed]
- Suganya, K.; Koo, B.-S. Gut–Brain Axis: Role of Gut Microbiota on Neurological Disorders and How Probiotics/Prebiotics Beneficially Modulate Microbial and Immune Pathways to Improve Brain Functions. Int. J. Mol. Sci. 2020, 21, 7551. [Google Scholar] [CrossRef]
- Wei, H.; Yu, C.; Zhang, C.; Ren, Y.; Guo, L.; Wang, T.; Chen, F.; Li, Y.; Zhang, X.; Wang, H.; et al. Butyrate ameliorates chronic alcoholic central nervous damage by suppressing microglia-mediated neuroinflammation and modulating the microbiome-gut-brain axis. Biomed. Pharmacother. 2023, 160, 114308. [Google Scholar] [CrossRef]
- Padilla, S.; Nurden, A.T.; Prado, R.; Nurden, P.; Anitua, E. Healing through the lens of immunothrombosis: Biology-inspired, evolution-tailored, and human-engineered biomimetic therapies. Biomaterials 2021, 279, 121205. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.M.; Da, L.C.; Wang, R.; Wang, L.; Jiang, Y.L.; Zhang, X.Z.; Li, Y.X.; Lei, X.X.; Song, Y.T.; Zou, C.Y.; et al. Promotion of uterine reconstruction by a tissue-engineered uterus with biomimetic structure and extracellular matrix microenvironment. Sci. Adv. 2023, 9, eadi6488. [Google Scholar]
- Chen, R.; Yang, J.; Wu, M.; Zhao, D.; Yuan, Z.; Zeng, L.; Hu, J.; Zhang, X.; Wang, T.; Xu, J.; et al. M2 Macrophage Hybrid Membrane-Camouflaged Targeted Biomimetic Nanosomes to Reprogram Inflammatory Microenvironment for Enhanced Enzyme-Thermo-Immunotherapy. Adv. Mater. 2023, 35, e2304123. [Google Scholar] [PubMed]
- Hua, H.; Wang, Y.; Wang, X.; Wang, S.; Zhou, Y.; Liu, Y.; Liang, Z.; Ren, H.; Lu, S.; Wu, S.; et al. Remodeling ceramide homeostasis promotes functional maturation of human pluripotent stem cell-derived β cells. Cell Stem Cell 2024, 31, 850–865.e10. [Google Scholar] [PubMed]
- Hu, H.; Hua, S.Y.; Lin, X.; Lu, F.; Zhang, W.; Zhou, L.; Cui, J.; Wang, R.; Xia, J.; Xu, F.; et al. Hybrid Biomimetic Membrane Coated Particles-Mediated Bacterial Ferroptosis for Acute MRSA Pneumonia. ACS Nano 2023, 17, 11692–11712. [Google Scholar]
- McDougald, D.; Rice, S.A.; Kjelleberg, S. Bacterial quorum sensing and interference by naturally occurring biomimics. Anal. Bioanal. Chem. 2007, 387, 445–453. [Google Scholar] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [PubMed]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Belinchón-deMiguel, P.; Ramos-Campo, D.J.; Curiel-Regueros, A.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. The Interplay of Sports and Nutrition in Neurological Health and Recovery. J. Clin. Med. 2024, 13, 2065. [Google Scholar] [CrossRef]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cellular and Molecular Life Sciences. Cell. Mol. Life Sci. 2018, 76, 473–493. [Google Scholar] [PubMed]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of Antibiotics on Gut Microbiota. Dig. Dis. 2016, 34, 260–268. [Google Scholar]
- Rutayisire, E.; Huang, K.; Liu, Y.; Tao, F. The mode of delivery affects the diversity and colonization pattern of the gut microbiota during the first year of infants’ life: A systematic review. BMC Gastroenterol. 2016, 16, 86. [Google Scholar]
- Li, L.-Y.; Han, J.; Wu, L.; Fang, C.; Li, W.-G.; Gu, J.-M.; Deng, T.; Qin, C.-J.; Nie, J.-Y.; Zeng, X.-T. Alterations of gut microbiota diversity, composition and metabonomics in testosterone-induced benign prostatic hyperplasia rats. Mil. Med. Res. 2022, 9, 12. [Google Scholar] [CrossRef]
- Procházková, N.; Falony, G.; Dragsted, L.O.; Licht, T.R.; Raes, J.; Roager, H.M. Advancing human gut microbiota research by considering gut transit time. Gut 2022, 72, 180–191. [Google Scholar] [CrossRef]
- Guzior, D.V.; Quinn, R.A. Review: Microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Alkhalil, S.S. The role of bacteriophages in shaping bacterial composition and diversity in the human gut. Front. Microbiol. 2023, 14, 1232413. [Google Scholar] [CrossRef] [PubMed]
- Paukkonen, I.; Törrönen, E.-N.; Lok, J.; Schwab, U.; El-Nezami, H. The impact of intermittent fasting on gut microbiota: A systematic review of human studies. Front. Nutr. 2024, 11, 1342787. [Google Scholar]
- Vital, M.; Rud, T.; Rath, S.; Pieper, D.H.; Schlüter, D. Diversity of Bacteria Exhibiting Bile Acid-inducible 7α-dehydroxylation Genes in the Human Gut. Comput. Struct. Biotechnol. J. 2019, 17, 1016–1019. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Rodionov, D.A.; Osterman, A.L.; Peterson, S.N. B Vitamins and Their Role in Immune Regulation and Cancer. Nutrients 2020, 12, 3380. [Google Scholar] [CrossRef]
- Menees, K.B.; Otero, B.A.; Tansey, M.G. Microbiome influences on neuro-immune interactions in neurodegenerative disease. Int. Rev. Neurobiol. 2022, 167, 25–57. [Google Scholar] [PubMed]
- MacPherson, K.P.; Eidson, L.N.; Houser, M.C.; Weiss, B.E.; Gollihue, J.L.; Herrick, M.K.; de Sousa Rodrigues, M.E.; Sniffen, L.; Weekman, E.M.; Hamilton, A.M.; et al. Soluble TNF mediates amyloid-independent, diet-induced alterations to immune and neuronal functions in an Alzheimer’s disease mouse model. Front. Cell Neurosci. 2023, 17, 895017. [Google Scholar] [CrossRef] [PubMed]
- Aho, V.T.; Houser, M.C.; Pereira, P.A.; Chang, J.; Rudi, K.; Paulin, L.; Hertzberg, V.; Auvinen, P.; Tansey, M.G.; Scheperjans, F. Relationships of gut microbiota, short-chain fatty acids, inflammation, and the gut barrier in Parkinson’s disease. Mol. Neurodegener. 2021, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Jiang, P.; Yu, F.; Zhou, X.; Shi, H.; He, Q.; Song, X. Dissecting causal links between gut microbiota, inflammatory cytokines, and DLBCL: A Mendelian randomization study. Blood Adv. 2024, 8, 2268–2278. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Wu, L.; Li, X. IL-17 family: Cytokines, receptors and signaling. Cytokine 2013, 64, 477–485. [Google Scholar]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Brockmann, L.; Tran, A.; Huang, Y.; Edwards, M.; Ronda, C.; Wang, H.H.; Ivanov, I.I. Intestinal microbiota-specific Th17 cells possess regulatory properties and suppress effector T cells via c-MAF and IL-10. Immunity 2023, 56, 2719–2735.e7. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [PubMed]
- Effendi, R.M.R.A.; Anshory, M.; Kalim, H.; Dwiyana, R.F.; Suwarsa, O.; Pardo, L.M.; Nijsten, T.E.C.; Thio, H.B. Akkermansia muciniphila and Faecalibacterium prausnitzii in Immune-Related Diseases. Microorganisms 2022, 10, 2382. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Z.; Xu, H.M.; Liang, Y.J.; Xu, J.; Yue, N.N.; Zhang, Y.; Tian, C.M.; Yao, J.; Wang, L.S.; Nie, Y.Q.; et al. Edible exosome-like nanoparticles from portulaca oleracea L mitigate DSS-induced colitis via facilitating double-positive CD4+CD8+T cells expansion. J. Nanobiotechnol. 2023, 21, 309. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Kikuta, J.; Matsui, T.; Hasegawa, T.; Fujii, K.; Okuzaki, D.; Liu, Y.-C.; Yoshioka, T.; Seno, S.; Motooka, D.; et al. Periportal macrophages protect against commensal-driven liver inflammation. Nature 2024, 629, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Navarro-Jiménez, E.; Jimenez, M.; Hormeño-Holgado, A.; Martinez-Gonzalez, M.B.; Benitez-Agudelo, J.C.; Perez-Palencia, N.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. Impact of COVID-19 Pandemic in Public Mental Health: An Extensive Narrative Review. Sustainability 2021, 13, 3221. [Google Scholar] [CrossRef]
- Vicentini, F.A.; Keenan, C.M.; Wallace, L.E.; Woods, C.; Cavin, J.-B.; Flockton, A.R.; Macklin, W.B.; Belkind-Gerson, J.; Hirota, S.A.; Sharkey, K.A. Intestinal microbiota shapes gut physiology and regulates enteric neurons and glia. Microbiome 2021, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Huang, Z.; Yao, J.; Jin, Y. Extracellular vesicles-mediated interaction within intestinal microenvironment in inflammatory bowel disease. J. Adv. Res. 2021, 37, 221–233. [Google Scholar] [CrossRef]
- Shi, H.; Ge, X.; Ma, X.; Zheng, M.; Cui, X.; Pan, W.; Zheng, P.; Yang, X.; Zhang, P.; Hu, M.; et al. A fiber-deprived diet causes cognitive impairment and hippocampal microglia-mediated synaptic loss through the gut microbiota and metabolites. Microbiome 2021, 9, 223. [Google Scholar] [CrossRef]
- Kang, X.; Liu, C.; Ding, Y.; Ni, Y.; Ji, F.; Lau, H.C.; Jiang, L.; Sung, J.J.; Wong, S.H.; Yu, J. Roseburia intestinalis generated butyrate boosts anti-PD-1 efficacy in colorectal cancer by activating cytotoxic CD8 + T cells. Gut 2023, 72, 2112–2122. [Google Scholar] [PubMed]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.; Patterson, A.D. Bile acid metabolism and signaling, the microbiota, and metabolic disease. Pharmacol. Ther. 2022, 237, 108238. [Google Scholar] [CrossRef]
- Ko, C.-W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Gul, L.; Modos, D.; Fonseca, S.; Madgwick, M.; Thomas, J.P.; Sudhakar, P.; Booth, C.; Stentz, R.; Carding, S.R.; Korcsmaros, T. Extracellular vesicles produced by the human commensal gut bacterium Bacteroides thetaiotaomicron affect host immune pathways in a cell-type specific manner that are altered in inflammatory bowel disease. J. Extracell. Vesicles 2022, 11, e12189. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Morcillo, J.; Rodriguez-Besteiro, S.; Clemente-Suárez, V.J. The Nexus of Training Duration, Body Image, Nutritional Practices, and Mental Health: Insights from a Strength Training Cohort. Behav. Sci. 2024, 14, 267. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, T.L.; Eckstrom, K.; Lile, K.H.; Caldwell, S.; Heney, E.R.; Lahue, K.G.; D’alessandro, A.; Wargo, M.J.; Krementsov, D.N. Lactobacillus reuteri tryptophan metabolism promotes host susceptibility to CNS autoimmunity. Microbiome 2022, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef]
- Uceda, S.; Echeverry-Alzate, V.; Reiriz-Rojas, M.; Martínez-Miguel, E.; Pérez-Curiel, A.; Gómez-Senent, S.; Beltrán-Velasco, A.I. Gut Microbial Metabolome and Dysbiosis in Neurodegenerative Diseases: Psychobiotics and Fecal Microbiota Transplantation as a Therapeutic Approach—A Comprehensive Narrative Review. Int. J. Mol. Sci. 2023, 24, 13294. [Google Scholar] [CrossRef]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Leser, T.; Baker, A. Bifidobacterium adolescentis—A beneficial microbe. Benef. Microbes 2023, 14, 525–551. [Google Scholar] [CrossRef]
- Yang, S.-C.; Lin, C.-H.; Aljuffali, I.A.; Fang, J.-Y. Current pathogenic Escherichia coli foodborne outbreak cases and therapy development. Arch. Microbiol. 2017, 199, 811–825. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Liu, X. The microbiota–gut–brain axis and neurodevelopmental disorders. Protein Cell. 2023, 14, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huh, J.R.; Shah, K. Microbiota and the gut-brain-axis: Implications for new therapeutic design in the CNS. EBioMedicine 2022, 77, 103908. [Google Scholar] [CrossRef]
- Kuijer, E.J.; Steenbergen, L. The microbiota-gut-brain axis in hippocampus-dependent learning and memory: Current state and future challenges. Neurosci. Biobehav. Rev. 2023, 152, 105296. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Mhanna, A.; Martini, N.; Hmaydoosh, G.; Hamwi, G.; Jarjanazi, M.; Zaifah, G.; Kazzazo, R.; Mohamad, A.H.; Alshehabi, Z. The correlation between gut microbiota and both neurotransmitters and mental disorders: A narrative review. Medicine 2024, 103, e37114. [Google Scholar] [CrossRef]
- Naufel, M.F.; Truzzi, G.d.M.; Ferreira, C.M.; Coelho, F.M.S. The brain-gut-microbiota axis in the treatment of neurologic and psychiatric disorders. Arq. de Neuro-Psiquiatr. 2023, 81, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.F.; Zhu, Y.L.; Zhou, Z.L.; Jia, X.B.; Xu, Y.D.; Yang, Q.; Cui, C.; Shen, Y.Q. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson’s disease mice: Gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The Microbiota–Gut–Brain Axis in Psychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 11245. [Google Scholar] [CrossRef]
- Nikolova, V.L.; Smith, M.R.B.; Hall, L.J.; Cleare, A.J.; Stone, J.M.; Young, A.H. Perturbations in Gut Microbiota Composition in Psychiatric Disorders. JAMA Psychiatry 2021, 78, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Lai, X.; Li, C.; Ding, D.; Wang, Y.; Zhu, Y. The Role of Gut Microbiota in Various Neurological and Psychiatric Disorders—An Evidence Mapping Based on Quantified Evidence. Mediat. Inflamm. 2023, 2023, 5127157. [Google Scholar] [CrossRef] [PubMed]
- Gazerani, P. Probiotics for Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 4121. [Google Scholar] [CrossRef]
- Generoso, J.S.; Giridharan, V.V.; Lee, J.; Macedo, D.; Barichello, T. The role of the microbiota-gut-brain axis in neuropsychiatric disorders. Rev. Bras. de Psiquiatr. 2021, 43, 293–305. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Brain-Gut-Microbiota Axis and Mental Health. Psychosom. Med. 2017, 79, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Gao, Y.; Yang, R. Gut Microbiota-Derived Tryptophan Metabolites Maintain Gut and Systemic Homeostasis. Cells 2022, 11, 2296. [Google Scholar] [CrossRef]
- Swer, N.M.; Venkidesh, B.S.; Murali, T.S.; Mumbrekar, K.D. Gut microbiota-derived metabolites and their importance in neurological disorders. Mol. Biol. Rep. 2023, 50, 1663–1675. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Merino del Portillo, M.; Clemente-Suárez, V.J.; Ruisoto, P.; Jimenez, M.; Ramos-Campo, D.J.; Beltran-Velasco, A.I.; Martínez-Guardado, I.; Rubio-Zarapuz, A.; Navarro-Jiménez, E.; Tornero-Aguilera, J.F. Nutritional Modulation of the Gut–Brain Axis: A Comprehensive Review of Dietary Interventions in Depression and Anxiety Management. Metabolites 2024, 14, 549. [Google Scholar] [CrossRef] [PubMed]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Zhou, M.; Fan, Y.; Xu, L.; Yu, Z.; Wang, S.; Xu, H.; Zhang, J.; Zhang, L.; Liu, W.; Wu, L.; et al. Microbiome and tryptophan metabolomics analysis in adolescent depression: Roles of the gut microbiota in the regulation of tryptophan-derived neurotransmitters and behaviors in human and mice. Microbiome 2023, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan Metabolism and Gut-Brain Homeostasis. Int. J. Mol. Sci. 2021, 22, 2973. [Google Scholar] [CrossRef] [PubMed]
- Krause, F.F.; Mangold, K.I.; Ruppert, A.-L.; Leister, H.; Hellhund-Zingel, A.; Krol, A.L.; Pesek, J.; Watzer, B.; Winterberg, S.; Raifer, H.; et al. Clostridium sporogenes -derived metabolites protect mice against colonic inflammation. Gut Microbes 2024, 16, 2412669. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Koay, Y.C.; Pan, C.; Zhou, Z.; Tang, W.; Wilcox, J.; Li, X.S.; Zagouras, A.; Marques, F.; Allayee, H.; et al. Indole-3-Propionic Acid Protects Against Heart Failure With Preserved Ejection Fraction. Circ. Res. 2024, 134, 371–389. [Google Scholar] [CrossRef]
- Queiroz, S.d.S.; Jofre, F.M.; Bianchini, I.d.A.; Boaes, T.d.S.; Bordini, F.W.; Chandel, A.K.; Felipe, M.d.G.d.A. Current advances in Candida tropicalis: Yeast overview and biotechnological applications. Biotechnol. Appl. Biochem. 2023, 70, 2069–2087. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef] [PubMed]
- KishanRaj, S.; Sumitha, S.; Tang, T.-H.; Citartan, M.; Chinni, S.V. Comparative genomic identification and characterization of npcRNA homologs in Proteus vulgaris. J. Biosci. 2021, 46, 1–10. [Google Scholar] [CrossRef]
- Dowling, L.R.; Strazzari, M.R.; Keely, S.; Kaiko, G.E. Enteric nervous system and intestinal epithelial regulation of the gut-brain axis. J. Allergy Clin. Immunol. 2022, 150, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, B.; Luo, M.; Xie, L.; Lu, M.; Lu, X.; Zhang, S.; Wei, L.; Zhou, X.; Yao, B.; et al. Microbiota-indole 3-propionic acid-brain axis mediates abnormal synaptic pruning of hippocampal microglia and susceptibility to ASD in IUGR offspring. Microbiome 2023, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, X.; Chen, J.; Dai, Z.; Zhu, Y.; Wu, T.; Liu, Q.; Qin, H.; Zhang, Y.; Chen, H. Enzyme-like biomimetic oral-agent enabling modulating gut microbiota and restoring redox homeostasis to treat inflammatory bowel disease. Bioact. Mater. 2024, 35, 167–180. [Google Scholar] [CrossRef]
- Yu, Z.; Chen, W.; Zhang, L.; Chen, Y.; Chen, W.; Meng, S.; Lu, L.; Han, Y.; Shi, J. Gut-derived bacterial LPS attenuates incubation of methamphetamine craving via modulating microglia. Brain Behav. Immun. 2023, 111, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Moossavi, S.; Arrieta, M.-C.; Sanati-Nezhad, A.; Bishehsari, F. Gut-on-chip for ecological and causal human gut microbiome research. Trends Microbiol. 2022, 30, 710–721. [Google Scholar] [CrossRef]
- Wang, X.; Tian, R.; Liang, C.; Jia, Y.; Zhao, L.; Xie, Q.; Huang, F.; Yuan, H. Biomimetic nanoplatform with microbiome modulation and antioxidant functions ameliorating insulin resistance and pancreatic β-cell dysfunction for T2DM management. Biomaterials 2025, 313, 122804. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Kang, Z.; Shen, J.; Zhao, R.; Miao, Y.; Zhang, L.; Zheng, Z.; Zhang, Z.; Liu, N.; Wang, C.; et al. An emerging antibacterial nanovaccine for enhanced chemotherapy by selectively eliminating tumor-colonizing bacteria. Sci. Bull. 2024, 69, 2565–2579. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Ding, N.; Guo, H.; Hua, R.; Lin, Z.; Tian, H.; Yu, Y.; Fan, D.; Yuan, Z.; Gonzalez, F.J.; et al. A gut microbiota-bile acid axis promotes intestinal homeostasis upon aspirin-mediated damage. Cell Host Microbe 2024, 32, 191–208.e9. [Google Scholar] [CrossRef]
- Xiao, L.; Tang, R.; Wang, J.; Wan, D.; Yin, Y.; Xie, L. Gut microbiota bridges the iron homeostasis and host health. Sci. China Life Sci. 2023, 66, 1952–1975. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Costa, A.; Becagli, M.V.; Monroy, M.M.; Provensi, G.; Passani, M.B. Gut microbiota and oleoylethanolamide in the regulation of intestinal homeostasis. Front. Endocrinol. 2023, 14, 1135157. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M. Inflammasomes in the gastrointestinal tract: Infection, cancer and gut microbiota homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 721–737. [Google Scholar] [CrossRef] [PubMed]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: A narrative review. Intern. Emerg. Med. 2024, 19, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yuan, T.-J.; Wan, Y.; Li, W.-W.; Liu, C.; Jiang, S.; Duan, J.-A. Quorum Sensing: A New Perspective to Reveal the Interaction Between Gut Microbiota and Host. Futur. Microbiol. 2022, 17, 293–309. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Cabral, V.; Torcato, I.; Xavier, K.B. Deciphering the quorum-sensing lexicon of the gut microbiota. Cell Host Microbe 2023, 31, 500–512. [Google Scholar] [CrossRef]
- Xavier, K.B. Bacterial interspecies quorum sensing in the mammalian gut microbiota. Comptes Rendus Biol. 2018, 341, 297–299. [Google Scholar] [CrossRef]
- Zhu, P.; Lu, T.; Chen, Z.; Liu, B.; Fan, D.; Li, C.; Wu, J.; He, L.; Zhu, X.; Du, Y.; et al. 5-hydroxytryptamine produced by enteric serotonergic neurons initiates colorectal cancer stem cell self-renewal and tumorigenesis. Neuron 2022, 110, 2268–2282.e4. [Google Scholar] [CrossRef]
- Landry, B.P.; Tabor, J.J. Engineering Diagnostic and Therapeutic Gut Bacteria. Microbiol. Spectr. 2017, 5, 331–361. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Chiang, H.C.; Gordon, J.I. Mucosal Glycan Foraging Enhances Fitness and Transmission of a Saccharolytic Human Gut Bacterial Symbiont. Cell Host Microbe 2008, 4, 447–457. [Google Scholar] [CrossRef]
- Silvestri, A.; Gil-Gomez, A.; Vitale, M.; Braga, D.; Demitri, C.; Brescia, P.; Madaghiele, M.; Spadoni, I.; Jones, B.; Fornasa, G.; et al. Biomimetic superabsorbent hydrogel acts as a gut protective dynamic exoskeleton improving metabolic parameters and expanding A. muciniphila. Cell Rep. Med. 2023, 4, 101235. [Google Scholar] [CrossRef] [PubMed]
- Zu, M.; Liu, G.; Xu, H.; Zhu, Z.; Zhen, J.; Li, B.; Shi, X.; Shahbazi, M.; Reis, R.L.; Kundu, S.C.; et al. Extracellular Vesicles from Nanomedicine-Trained Intestinal Microbiota Substitute for Fecal Microbiota Transplant in Treating Ulcerative Colitis. Adv. Mater. 2024, 36, e2409138. [Google Scholar] [CrossRef]
- Zu, M.; Liu, G.; Chen, N.; Chen, L.; Gao, Q.; Reis, R.L.; Kundu, S.C.; Jin, M.; Xiao, B.; Shi, X. Oral exosome-like nanovesicles from Phellinus linteus suppress metastatic hepatocellular carcinoma by reactive oxygen species generation and microbiota rebalancing. Nanoscale 2024, 16, 8046–8059. [Google Scholar] [CrossRef] [PubMed]
- Gościniak, A.; Eder, P.; Walkowiak, J.; Cielecka-Piontek, J. Artificial Gastrointestinal Models for Nutraceuticals Research—Achievements and Challenges: A Practical Review. Nutrients 2022, 14, 2560. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.D.; Durand, H.K.; Bloom, R.J.; Mukherjee, S.; David, L.A. Dynamic linear models guide design and analysis of microbiota studies within artificial human guts. Microbiome 2018, 6, 202. [Google Scholar]
- Xiang, Y.; Wen, H.; Yu, Y.; Li, M.; Fu, X.; Huang, S. Gut-on-chip: Recreating human intestine in vitro. J. Tissue Eng. 2020, 11, 2041731420965318. [Google Scholar] [CrossRef] [PubMed]
- Calvigioni, M.; Panattoni, A.; Biagini, F.; Donati, L.; Mazzantini, D.; Massimino, M.; Daddi, C.; Celandroni, F.; Vozzi, G.; Ghelardi, E. Development of an In Vitro Model of the Gut Microbiota Enriched in Mucus-Adhering Bacteria. Microbiol. Spectr. 2023, 11, e0033623. [Google Scholar] [CrossRef]
- Mabwi, H.A.; Hitayezu, E.; Mauliasari, I.R.; Mwaikono, K.S.; Yoon, H.S.; Komba, E.V.; Pan, C.-H.; Cha, K.H. Simulation of the mucosal environment in the re-construction of the synthetic gut microbial ecosystem. J. Microbiol. Methods 2021, 191, 106351. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.E.; Dreesen, D.W.; Wiegert, R.G. A simulation of microbial competition in the human colonic ecosystem. Appl. Environ. Microbiol. 1996, 62, 3632–3639. [Google Scholar] [CrossRef]
- Hellweger, F.L. Combining Molecular Observations and Microbial Ecosystem Modeling: A Practical Guide. Annu. Rev. Mar. Sci. 2020, 12, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Guevarra, R.B.; Kim, Y.-T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.-H. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Qu, C.; Wang, B.; Liang, S.; Zeng, B. Probiotics for Preventing and Treating Small Intestinal Bacterial Overgrowth. J. Clin. Gastroenterol. 2017, 51, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhong, D.; Qi, Y.; Liu, C.; Liu, X.; Chen, S.; Yan, S.; Zhou, M. Bioinspired pH-Responsive Microalgal Hydrogels for Oral Insulin Delivery with Both Hypoglycemic and Insulin Sensitizing Effects. ACS Nano. 2023, 17, 14161–14175. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, Z.; Zhang, M.; Meng, L.; Ming, Z.; Liu, J. Bioinspired oral delivery of gut microbiota by self-coating with biofilms. Sci. Adv. 2020, 6, eabb1952. [Google Scholar] [CrossRef]
- Wang, J.; Ninan, N.; Nguyen, N.H.; Nguyen, M.T.; Sahu, R.; Nguyen, T.T.; Mierczynska-Vasilev, A.; Vasilev, K.; Truong, V.K.; Tang, Y. Biomimetic Bacterium-like Particles Loaded with Aggregation-Induced Emission Photosensitizers as Plasma Coatings for Implant-Associated Infections. ACS Appl. Mater. Interfaces 2024, 16, 18449–18458. [Google Scholar] [CrossRef]
- Gui, Y.; Sun, Q.; Li, K.; Lin, L.; Zhou, H.; Ma, J.; Li, C. Bioinspired gelated cell sheet–supported lactobacillus biofilm for aerobic vaginitis diagnosis and treatment. Sci. Adv. 2024, 10, eadq2732. [Google Scholar] [CrossRef]
- Chen, T.; Meng, W.; Li, Y.; Li, X.; Yu, X.; Qi, J.; Ding, D.; Li, W. Probiotics Armed with In Situ Mineralized Nanocatalysts and Targeted Biocoatings for Multipronged Treatment of Inflammatory Bowel Disease. Nano Lett. 2024, 24, 7321–7331. [Google Scholar] [CrossRef]
- Alli, S.R.; Gorbovskaya, I.; Liu, J.C.W.; Kolla, N.J.; Brown, L.; Müller, D.J. The Gut Microbiome in Depression and Potential Benefit of Prebiotics, Probiotics and Synbiotics: A Systematic Review of Clinical Trials and Observational Studies. Int. J. Mol. Sci. 2022, 23, 4494. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Nguyen, P.T.; Pham, M.N.; Razafindralambo, H.; Hoang, Q.K.; Nguyen, H.T. Synbiotics: A New Route of Self-production and Applications to Human and Animal Health. Probiotics Antimicrob. Proteins. 2022, 14, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Mounir, M.; Ibijbijen, A.; Farih, K.; Rabetafika, H.N.; Razafindralambo, H.L. Synbiotics and Their Antioxidant Properties, Mechanisms, and Benefits on Human and Animal Health: A Narrative Review. Biomolecules 2022, 12, 1443. [Google Scholar] [CrossRef] [PubMed]
- Allahyari, P.; Torki, S.A.; Kavkani, B.A.; Mahmoudi, Z.; Hoseini, M.S.M.; Moradi, M.; Alami, F.; Mohammadian, M.K.; Bani, S.B.S.; Mobarakeh, K.A.; et al. A systematic review of the beneficial effects of prebiotics, probiotics, and synbiotics on ADHD. Neuropsychopharmacol. Rep. 2024, 44, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Olas, B. Probiotics, Prebiotics and Synbiotics—A Promising Strategy in Prevention and Treatment of Cardiovascular Diseases? Int. J. Mol. Sci. 2020, 21, 9737. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.W.; Khoo, S.C.; Paul, R.P.M.; Luang-In, V.; Lam, S.D.; Ma, N.L. Potential of Synbiotics and Probiotics as Chemopreventive Agent. Probiotics Antimicrob. Proteins 2024, 16, 2085–2101. [Google Scholar] [CrossRef]
- Hojsak, I.; Kolaček, S.; Mihatsch, W.; Mosca, A.; Shamir, R.; Szajewska, H.; Vandenplas, Y.; Akker, C.H.C.v.D.; Canani, R.B.; Dinleyici, E.C.; et al. Synbiotics in the Management of Pediatric Gastrointestinal Disorders. J. Pediatr. Gastroenterol. Nutr. 2023, 76, 102–108. [Google Scholar]
- Daelemans, S.; Deseck, V.; Levy, E.I.; Vandenplas, Y. Are pro- and/or synbiotics beneficial in Helicobacter pylori eradication therapy in children? A narrative review. Eur. J. Pediatr. 2022, 181, 3225–3234. [Google Scholar] [CrossRef]
- Rufino, M.N.; da Costa, A.L.; Jorge, E.N.; Paiano, V.F.; Camparoto, M.L.; Keller, R.; Bremer-Neto, H. Synbiotics improve clinical indicators of ulcerative colitis: Systematic review with meta-analysis. Nutr. Rev. 2022, 80, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Musazadeh, V.; Assadian, K.; Rajabi, F.; Faghfouri, A.H.; Soleymani, Y.; Kavyani, Z.; Najafiyan, B. The effect of synbiotics on liver enzymes, obesity indices, blood pressure, lipid profile, and inflammation in patients with non-alcoholic fatty liver: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2024, 208, 107398. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht, E.; Khalesi, S.; Singh, I.; Williams, L.T.; West, N.P.; Colson, N. Effect of probiotics and synbiotics on blood glucose: A systematic review and meta-analysis of controlled trials. Eur. J. Nutr. 2018, 57, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ding, T.; Ai, L. Editorial: Effects and mechanisms of probiotics, prebiotics, synbiotics and postbiotics on intestinal health and disease. Front. Cell Infect. Microbiol. 2024, 14, 1430312. [Google Scholar] [CrossRef]
- Porcari, S.; Fusco, W.; Spivak, I.; Fiorani, M.; Gasbarrini, A.; Elinav, E.; Cammarota, G.; Ianiro, G. Fine-tuning the gut ecosystem: The current landscape and outlook of artificial microbiome therapeutics. Lancet Gastroenterol. Hepatol. 2024, 9, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.S. SynMADE: Synthetic microbiota across diverse ecosystems. Trends Biotechnol. 2022, 40, 1405–1414. [Google Scholar] [CrossRef]
- Afrizal, A.; Jennings, S.A.; Hitch, T.C.; Riedel, T.; Basic, M.; Panyot, A.; Treichel, N.; Hager, F.T.; Wong, E.O.-Y.; Wolter, B.; et al. Enhanced cultured diversity of the mouse gut microbiota enables custom-made synthetic communities. Cell Host Microbe. 2022, 30, 1630–1645.e25. [Google Scholar] [CrossRef]
- Vrancken, G.; Gregory, A.C.; Huys, G.R.B.; Faust, K.; Raes, J. Synthetic ecology of the human gut microbiota. Nat. Rev. Microbiol. 2019, 17, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Carlström, C.I.; Field, C.M.; Bortfeld-Miller, M.; Müller, B.; Sunagawa, S.; Vorholt, J.A. Synthetic microbiota reveal priority effects and keystone strains in the Arabidopsis phyllosphere. Nat. Ecol. Evol. 2019, 3, 1445–1454. [Google Scholar] [CrossRef]
- Stecher, B. Establishing causality in Salmonella-microbiota-host interaction: The use of gnotobiotic mouse models and synthetic microbial communities. Int. J. Med. Microbiol. 2021, 311, 151484. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Díaz, A.; Ng, K.M.; Thomsen, T.; Real-Ramírez, I.; Dahan, D.; Dittmar, S.; Gonzalez, C.G.; Chavez, T.; Vasquez, K.S.; Nguyen, T.H.; et al. Establishment and characterization of stable, diverse, fecal-derived in vitro microbial communities that model the intestinal microbiota. Cell Host Microbe 2022, 30, 260–272.e5. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Castellanos, J.F.; Biclot, A.; Vrancken, G.; Huys, G.R.; Raes, J. Design of synthetic microbial consortia for gut microbiota modulation. Curr. Opin. Pharmacol. 2019, 49, 52–59. [Google Scholar] [CrossRef]
- Ioannou, A.; Berkhout, M.D.; Scott, W.T.; Blijenberg, B.; Boeren, S.; Mank, M.; Knol, J.; Belzer, C. Resource sharing of an infant gut microbiota synthetic community in combinations of human milk oligosaccharides. ISME J. 2024, 18, wrae209. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, W.; Zhang, F. The Next Generation Fecal Microbiota Transplantation: To Transplant Bacteria or Virome. Adv. Sci. 2023, 10, 2301097. [Google Scholar] [CrossRef] [PubMed]
- Porcari, S.; Benech, N.; Valles-Colomer, M.; Segata, N.; Gasbarrini, A.; Cammarota, G.; Sokol, H.; Ianiro, G. Key determinants of success in fecal microbiota transplantation: From microbiome to clinic. Cell Host Microbe 2023, 31, 712–733. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, I.; Ali, N.M.; Arshad, N.; Sajjad, S.; Rashid, F.; Hanif, U.; Ara, C.; Ulfat, M.; Andleeb, S.; Awan, U.F.; et al. Gut dysbiosis, inflammation and type 2 diabetes in mice using synthetic gut microbiota from diabetic humans. Braz. J. Biol. 2023, 83, e242818. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.J.; Plichta, D.; Hogstrom, L.; Borren, N.Z.; Lau, H.; Gregory, S.M.; Tan, W.; Khalili, H.; Clish, C.; Vlamakis, H.; et al. Multi-omics reveal microbial determinants impacting responses to biologic therapies in inflammatory bowel disease. Cell Host Microbe 2021, 29, 1294–1304.e4. [Google Scholar] [CrossRef]
- Everett, B.A.; Tran, P.; Prindle, A. Toward manipulating serotonin signaling via the microbiota–gut–brain axis. Curr. Opin. Biotechnol. 2022, 78, 102826. [Google Scholar] [CrossRef] [PubMed]
- Ooijevaar, R.E.; Terveer, E.M.; Verspaget, H.W.; Kuijper, E.J.; Keller, J.J. Clinical Application and Potential of Fecal Microbiota Transplantation. Annu. Rev. Med. 2019, 70, 335–351. [Google Scholar] [CrossRef]
- Costello, S.P.; Hughes, P.A.; Waters, O.; Bryant, R.V.; Vincent, A.D.; Blatchford, P.; Katsikeros, R.; Makanyanga, J.; Campaniello, M.A.; Mavrangelos, C.; et al. Effect of Fecal Microbiota Transplantation on 8-Week Remission in Patients With Ulcerative Colitis. JAMA 2019, 321, 156. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tan, G.; Zhu, Q.; Wang, C.; Ruan, G.; Ying, S.; Qie, J.; Hu, X.; Xiao, Z.; Xu, F.; et al. Efficacy of fecal microbiota transplantation in patients with Parkinson’s disease: Clinical trial results from a randomized, placebo-controlled design. Gut Microbes 2023, 15, 2284247. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, B.; Zheng, Q.; Li, H.; Meng, X.; Zhou, F.; Zhang, L. A Review of Gut Microbiota-Derived Metabolites in Tumor Progression and Cancer Therapy. Adv. Sci. 2023, 10, 2207366. [Google Scholar] [CrossRef] [PubMed]
- Quaranta, G.; Ianiro, G.; De Maio, F.; Guarnaccia, A.; Fancello, G.; Agrillo, C.; Iannarelli, F.; Bibbo, S.; Amedei, A.; Sanguinetti, M.; et al. “Bacterial Consortium”: A Potential Evolution of Fecal Microbiota Transplantation for the Treatment of Clostridioides difficile Infection. Biomed. Res. Int. 2022, 2022, 5787373. [Google Scholar] [CrossRef]
- Woloszynek, S.; Pastor, S.; Mell, J.C.; Nandi, N.; Sokhansanj, B.; Rosen, G.L. Engineering Human Microbiota: Influencing Cellular and Community Dynamics for Therapeutic Applications. Int. Rev. Cell Mol. Biol. 2016, 324, 67–124. [Google Scholar] [PubMed]
- Dong, Y.; Xu, T.; Xiao, G.; Hu, Z.; Chen, J. Opportunities and challenges for synthetic biology in the therapy of inflammatory bowel disease. Front. Bioeng. Biotechnol. 2022, 10, 909591. [Google Scholar] [CrossRef]
- Dang, Z.; Gao, M.; Wang, L.; Wu, J.; Guo, Y.; Zhu, Z.; Huang, H.; Kang, G. Synthetic bacterial therapies for intestinal diseases based on quorum-sensing circuits. Biotechnol. Adv. 2023, 65, 108142. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Song, Y.; Xu, Y.; Xu, H. Manipulating Microbiota in Inflammatory Bowel Disease Treatment: Clinical and Natural Product Interventions Explored. Int. J. Mol. Sci. 2023, 24, 11004. [Google Scholar] [CrossRef] [PubMed]
- Camacho, D.M.; Collins, K.M.; Powers, R.K.; Costello, J.C.; Collins, J.J. Next-Generation Machine Learning for Biological Networks. Cell 2018, 173, 1581–1592. [Google Scholar] [CrossRef]
- Maringanti, V.S.; Bucci, V.; Gerber, G.K. MDITRE: Scalable and Interpretable Machine Learning for Predicting Host Status from Temporal Microbiome Dynamics. mSystems 2022, 7, e00132-22. [Google Scholar] [CrossRef]
- Jones, E.W.; Carlson, J.M.; Sivak, D.A.; Ludington, W.B. Stochastic microbiome assembly depends on context. Proc. Natl. Acad. Sci. USA 2022, 119, e2115877119. [Google Scholar] [CrossRef]
- Parker, D.R.; Nugen, S.R. Bacteriophage-Based Bioanalysis. Annu. Rev. Anal. Chem. 2024, 17, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Biosensors and Bioassays Based on Lipases, Principles and Applications, a Review. Molecules 2019, 24, 616. [Google Scholar] [CrossRef]
- Hua, Y.; Ma, J.; Li, D.; Wang, R. DNA-Based Biosensors for the Biochemical Analysis: A Review. Biosensors 2022, 12, 183. [Google Scholar] [CrossRef]
- Marzorati, M.; Negroni, A.; Fava, F.; Verstraete, W.; Boon, N. Application of a molecular based approach for the early detection of short term 3-chloroaniline shock loads on activated sludge bacterial community and functionality. N. Biotechnol. 2013, 30, 763–771. [Google Scholar] [CrossRef]
- Lehtinen, T.; Santala, V.; Santala, S. Twin-layer biosensor for real-time monitoring of alkane metabolism. FEMS Microbiol. Lett. 2017, 364, fnx053. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Childers, W.S. A Biosensor for Detection of Indole Metabolites. ACS Synth. Biol. 2021, 10, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Zhai, H.; Hou, J. Biosensors design in yeast and applications in metabolic engineering. FEMS Yeast Res. 2019, 19, foz082. [Google Scholar] [CrossRef]
- Song, K.; Hwang, S.J.; Jeon, Y.; Yoon, Y. The Biomedical Applications of Biomolecule Integrated Biosensors for Cell Monitoring. Int. J. Mol. Sci. 2024, 25, 6336. [Google Scholar] [CrossRef]
- Liang, A.; Zhao, W.; Lv, T.; Zhu, Z.; Haotian, R.; Zhang, J.; Xie, B.; Yi, Y.; Hao, Z.; Sun, L.; et al. Advances in novel biosensors in biomedical applications. Talanta 2024, 280, 126709. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Katz, E. Digital biosensors with built-in logic for biomedical applications—Biosensors based on a biocomputing concept. Anal. Bioanal. Chem. 2010, 398, 1591–1603. [Google Scholar] [CrossRef]
- Misawa, N.; Osaki, T.; Takeuchi, S. Membrane protein-based biosensors. J. R. Soc. Interface. 2018, 15, 20170952. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Alberstein, R.G.; Zhang, Z.; Lu, Y.-S.; Chan, A.; Wahl, C.E.; Ha, J.S.; Hunka, D.E.; Boss, G.R.; Sailor, M.J.; et al. Designed 2D protein crystals as dynamic molecular gatekeepers for a solid-state device. Nat. Commun. 2024, 15, 6326. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beltrán-Velasco, A.I.; Clemente-Suárez, V.J. Harnessing Gut Microbiota for Biomimetic Innovations in Health and Biotechnology. Biomimetics 2025, 10, 73. https://doi.org/10.3390/biomimetics10020073
Beltrán-Velasco AI, Clemente-Suárez VJ. Harnessing Gut Microbiota for Biomimetic Innovations in Health and Biotechnology. Biomimetics. 2025; 10(2):73. https://doi.org/10.3390/biomimetics10020073
Chicago/Turabian StyleBeltrán-Velasco, Ana Isabel, and Vicente Javier Clemente-Suárez. 2025. "Harnessing Gut Microbiota for Biomimetic Innovations in Health and Biotechnology" Biomimetics 10, no. 2: 73. https://doi.org/10.3390/biomimetics10020073
APA StyleBeltrán-Velasco, A. I., & Clemente-Suárez, V. J. (2025). Harnessing Gut Microbiota for Biomimetic Innovations in Health and Biotechnology. Biomimetics, 10(2), 73. https://doi.org/10.3390/biomimetics10020073