
Biowelding 3D-Printed Biodigital Brick of Seashell-Based Biocomposite by Pleurotus ostreatus Mycelium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Elastic Biodigital Brick: V3 Linear Brick Model
1.2. Biocomposite Materials: Fiber Based to Platelet Based
Seashell Biocomposite from Recycled Food Waste
1.3. Mycelium Biocomposites and Biowelding
2. Results
2.1. Seashell Biocomposite Material Rheology
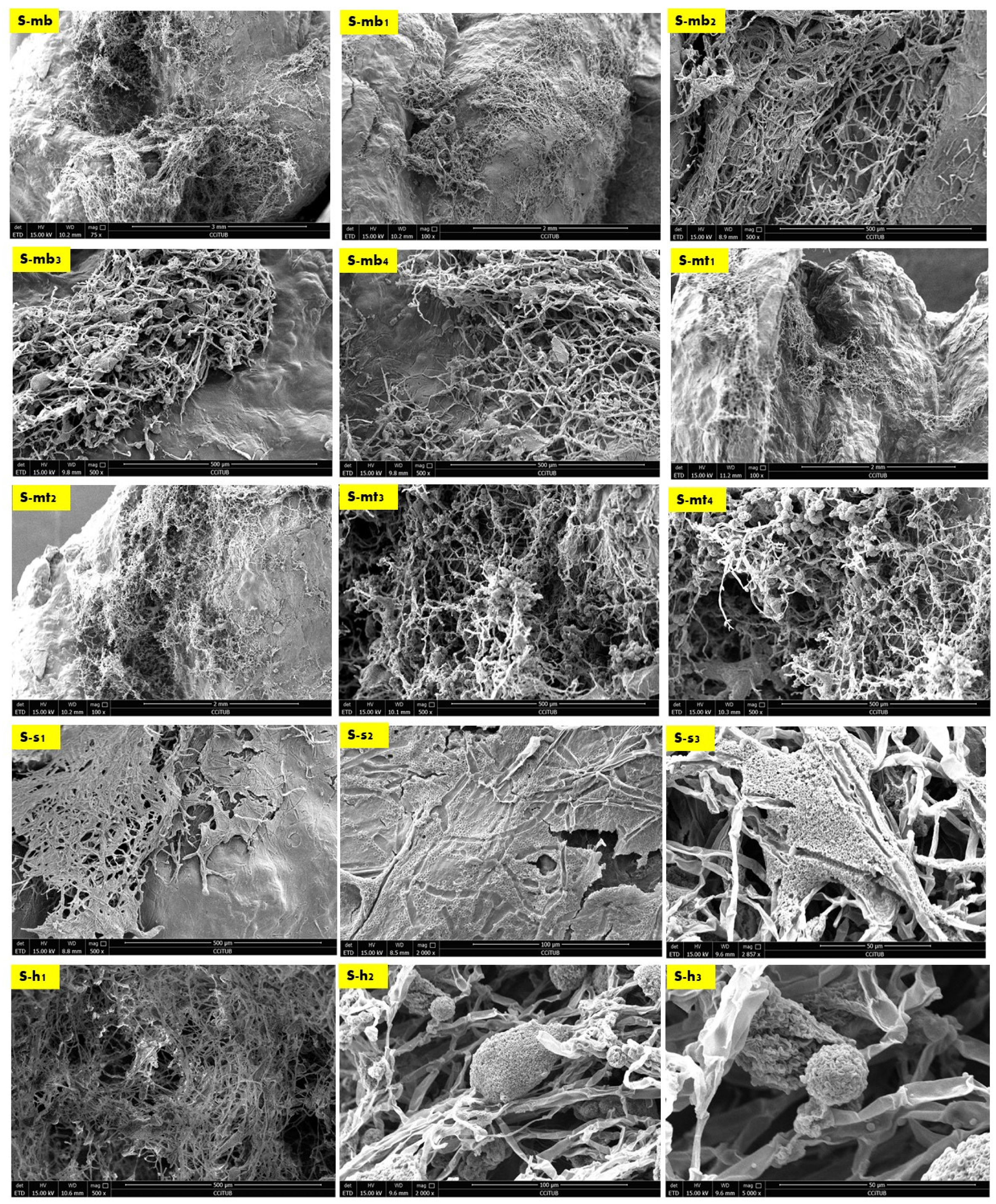
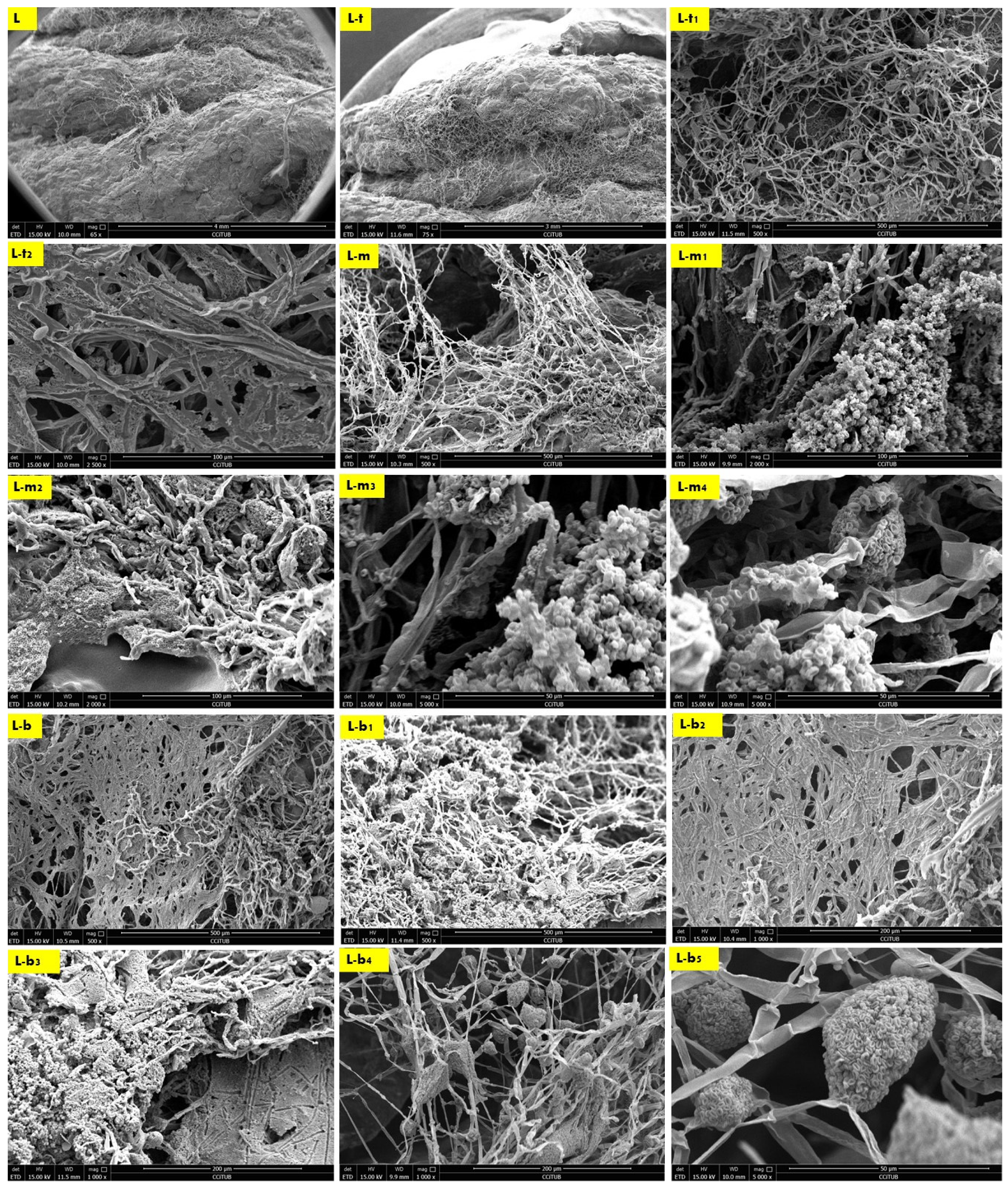
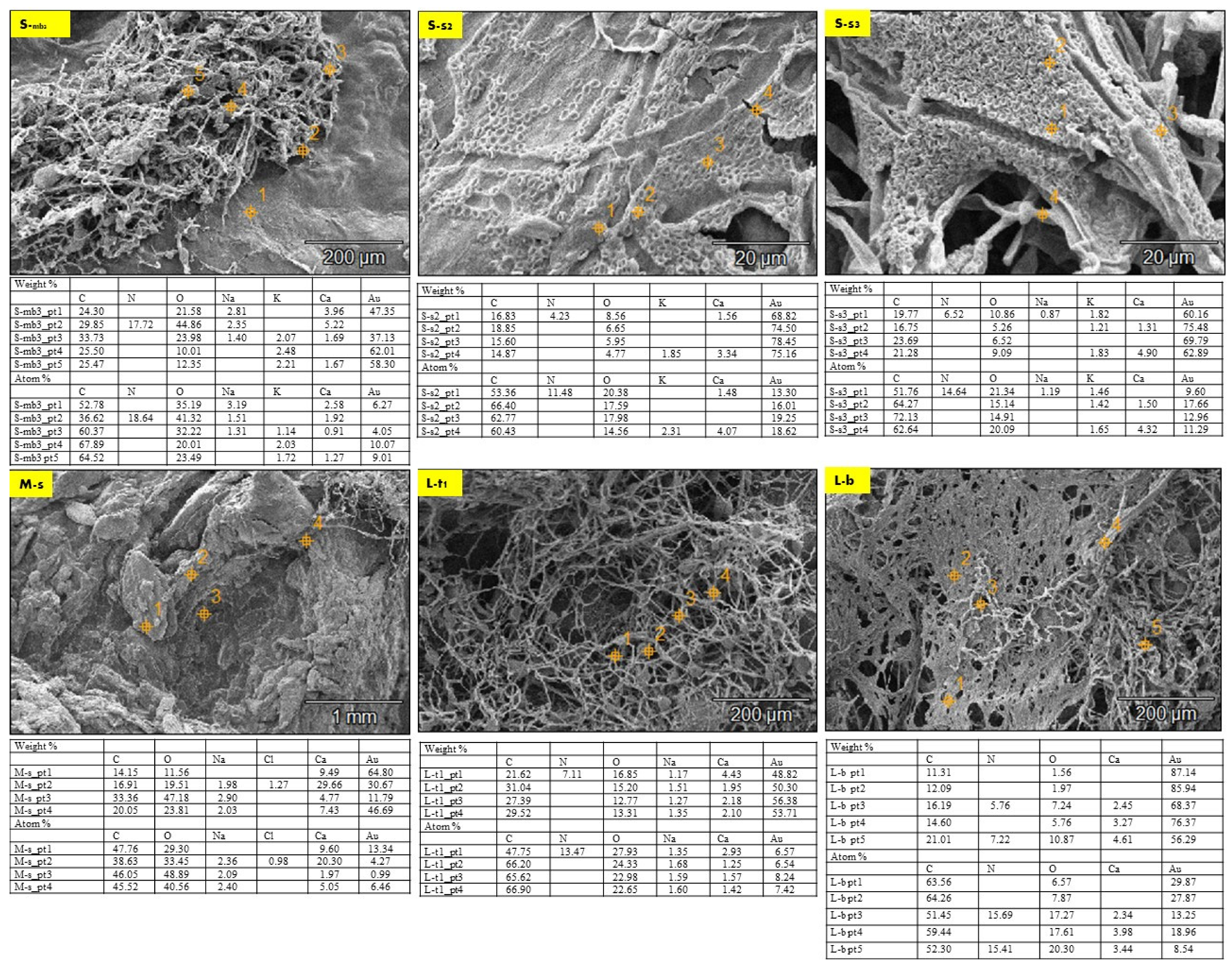
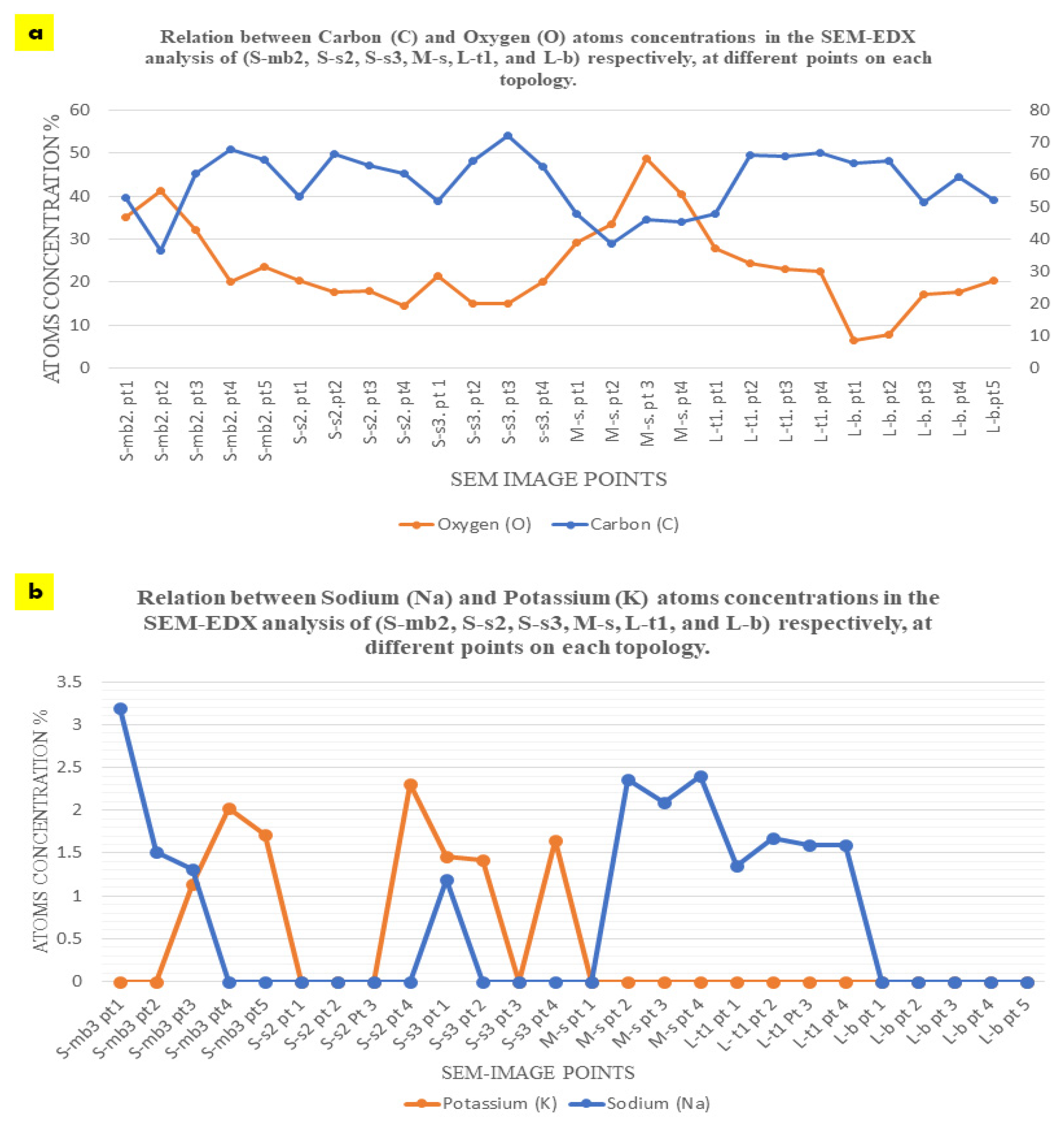
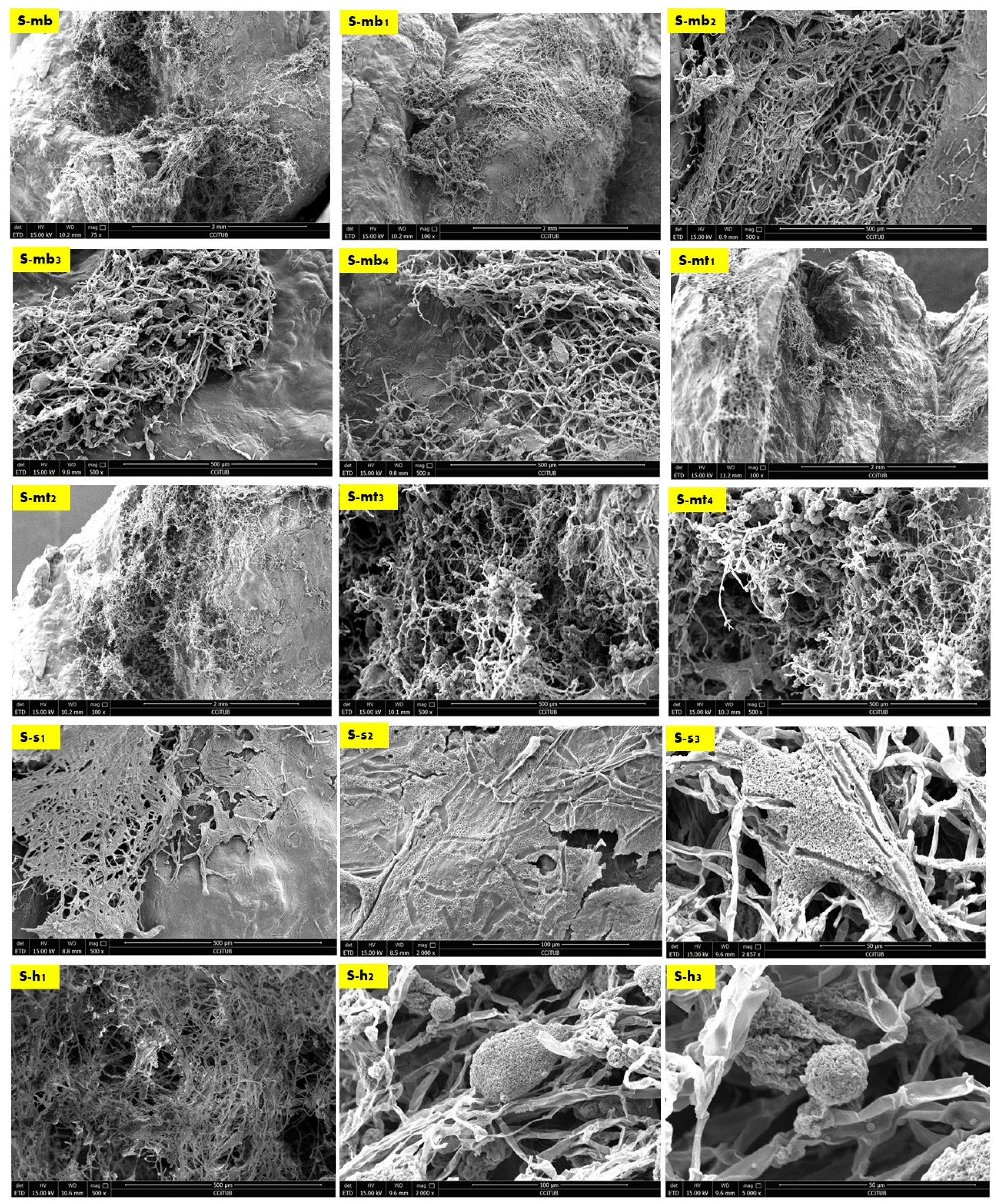
2.2. Biowelding by Mycelium (SEM and EDX-Chemical Analysis)
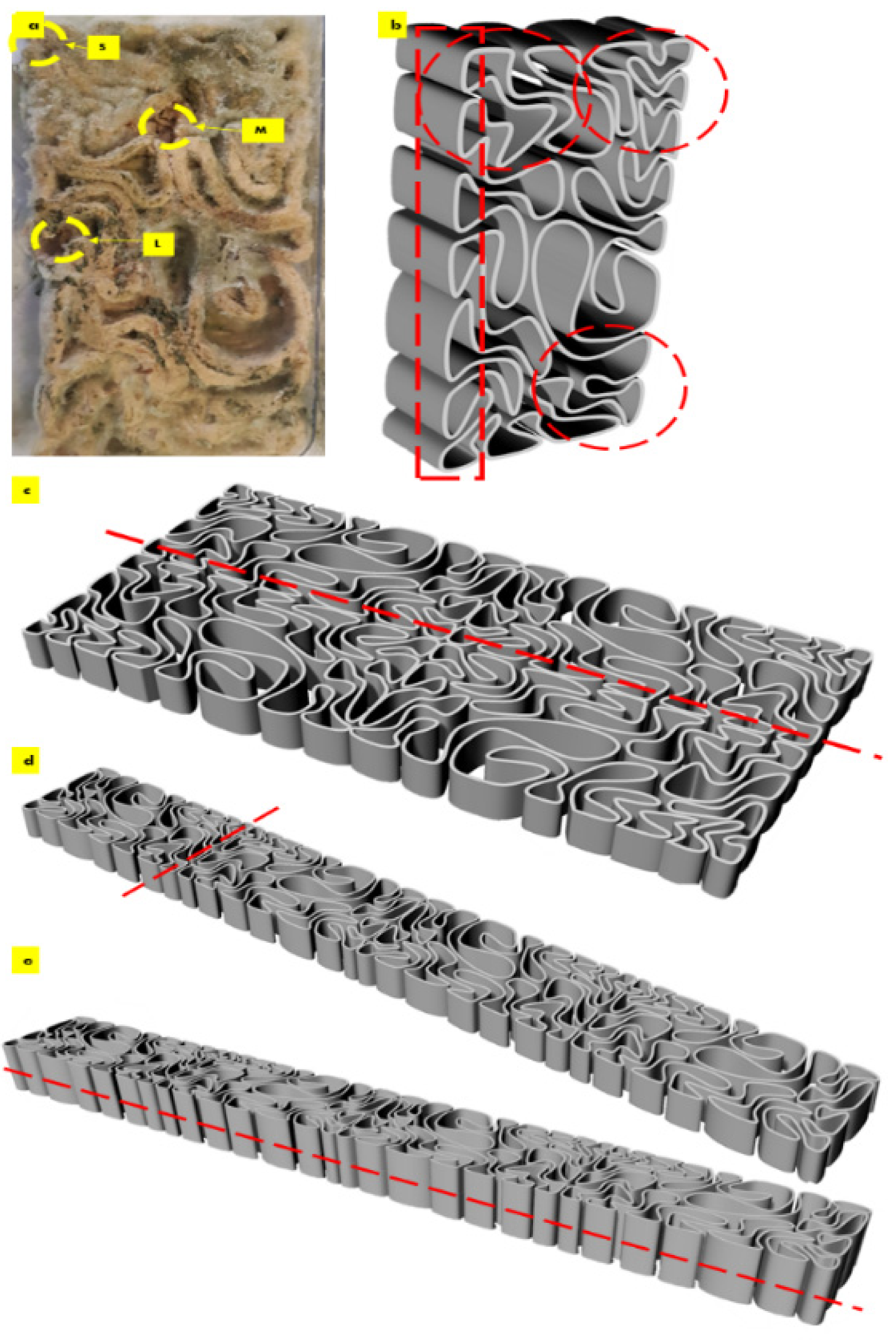
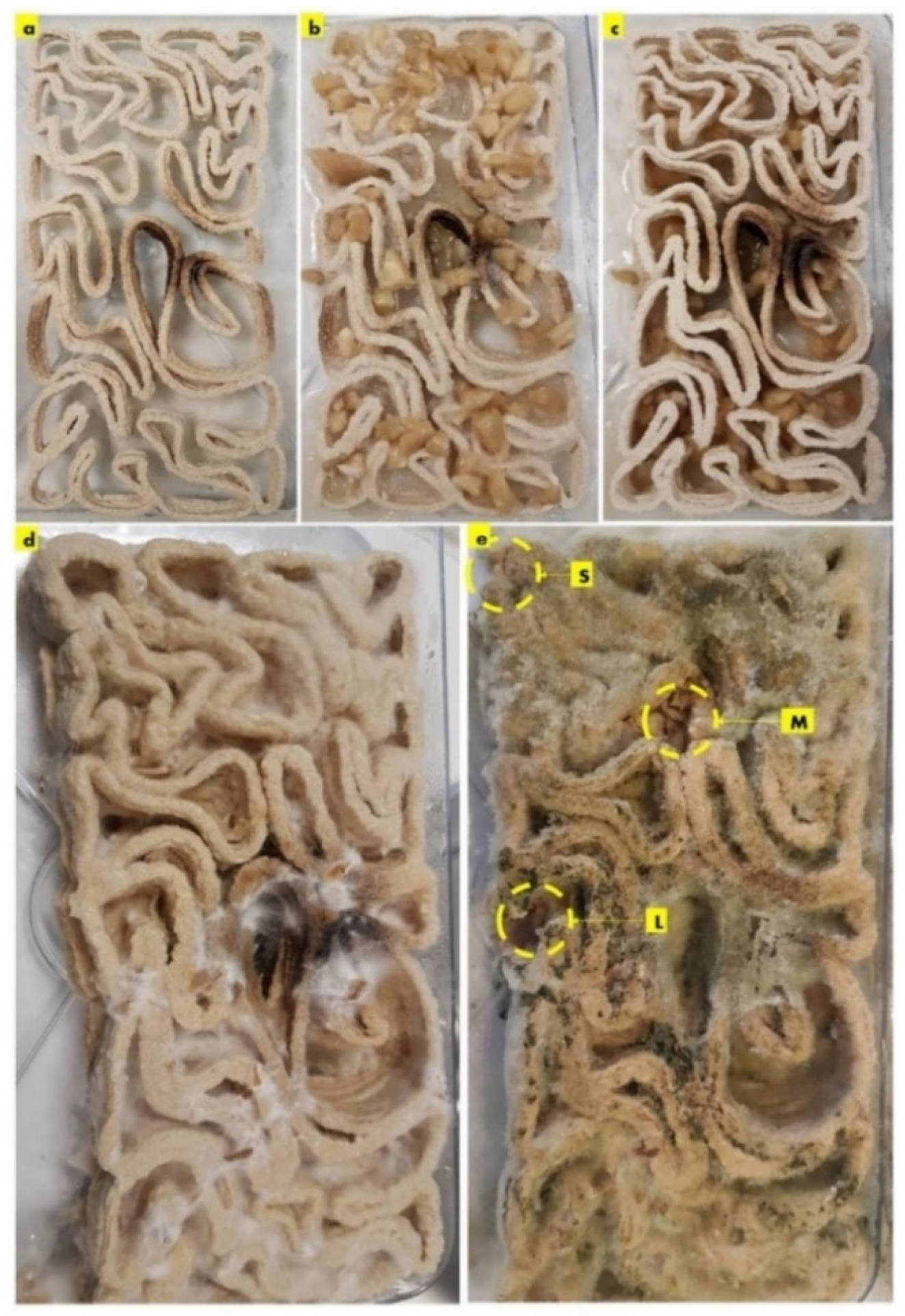
2.3. Seashell–Mycelium Biowelded Brick-Tessellation & Active Biowelding
3. Discussion
4. Materials and Methods
4.1. Seashell Biocomposite Material Preparation
4.2. 3D Printing by Paste Extrusion
4.3. P. ostreatus Culturing (Culture Medium and Conditions) and V3 Biocomposite Brick Inoculation (Seeding)
4.4. SEM Microscopy and EDX Chemical Analysis (Sampling, Sample Preparation)
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Estévez, A.T.; Abdallah, Y.K. The New Standard Is Biodigital: Durable and Elastic 3D-Printed Biodigital Clay Bricks. Biomimetics 2022, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, Y.K.; Estévez, A.T. 3D-Printed Biodigital Clay Bricks. Biomimetics 2021, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Msu.edu. 2021. Available online: https://tech.msu.edu/about/guidelines-policies/afs-retirement/ (accessed on 1 July 2023).
- Refiadi, G.; Syamsiar, Y.; Judawisastra, H. The Tensile Strength of Petung Bamboo Fiber Reinforced Epoxy Composites: The Effects of Alkali Treatment, Composites Manufacturing, and Water Absorption. IOP Conf. Ser. Mater. Sci. Eng. 2019, 547, 012043. [Google Scholar] [CrossRef]
- Judawisastra, H.; Sitohang, R.D.R.; Rosadi, M.S. Water absorption and tensile strength degradation of Petung bamboo (Dendrocalamus asper) fiber—Reinforced polymeric composites. Mater. Res. Express 2017, 4, 094003. [Google Scholar] [CrossRef]
- Judawisastra, H.; Sitohang, R.D.R.; Marta, L.; Mardiyati. Water absorption and its effect on the tensile properties of tapioca starch/polyvinyl alcohol bioplastics. IOP Conf. Ser. Mater. Sci. Eng. 2017, 223, 012066. [Google Scholar] [CrossRef]
- Gigante, V.; Cinelli, P.; Righetti, M.C.; Sandroni, M.; Tognotti, L.; Seggiani, M.; Lazzeri, A. Evaluation of Mussel Shells Powder as Reinforcement for PLA-Based Biocomposites. Int. J. Mol. Sci. 2020, 21, 5364. [Google Scholar] [CrossRef]
- Karunakaran, G.; Cho, E.-B.; Thirumurugan, K.; Kumar, G.S.; Kolesnikov, E.; Boobalan, S.; Janarthanan, G.; Pillai, M.M.; Rajendran, S. Mesoporous Mn-doped hydroxyapatite nanorods obtained via pyridinium chloride enabled microwave-assisted synthesis by utilizing Donax variabilis seashells for implant applications. Mater. Sci. Eng. C 2021, 126, 112170. [Google Scholar] [CrossRef]
- Sickels-Taves, L.B.; Sheehan, M.S.; Rovner, I. Computer-Assisted Optical Stereology in Historic Preservation: Tabby, a Case Study. APT Bull. 1997, 28, 30. [Google Scholar] [CrossRef]
- Razali, M.S.; Khimeche, K.; Melouki, R.; Boudjella, A.; Vroman, I.; Alix, S.; Ramdani, N. Preparation and properties enhancement of poly(lactic acid)/calcined-seashell biocomposites for 3D printing applications. J. Appl. Polym. Sci. 2021, 139, 51591. [Google Scholar] [CrossRef]
- Ramnath, B.V.; Jeykrishnan, J.; Ramakrishnan, G.; Barath, B.; Ejoelavendhan, E.; Raghav, P.A. Sea Shells And Natural Fibers Composites: A Review. Mater. Today Proc. 2018, 5, 1846–1851. [Google Scholar] [CrossRef]
- Razali, M.S.; Khimeche, K.; Boudjellal, A.; Ramdani, N. Effect of newly developed sintered seashell on the microhardness properties of biocomposites. Mater. Lett. 2021, 291, 129565. [Google Scholar] [CrossRef]
- Singamneni, S.; Behera, M.P.; Le Guen, M.; Zeidler, H. Mechanism of Bonding in Seashell Powder Based Ceramic Composites Used for Binder-Jet 3D Printing. Bioceram. Dev. Appl. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Tayeh, B.A.; Hasaniyah, M.W.; Zeyad, A.M.; Awad, M.M.; Alaskar, A.; Mohamed, A.M.; Alyousef, R. Durability and mechanical properties of seashell partially-replaced cement. J. Build. Eng. 2020, 31, 101328. [Google Scholar] [CrossRef]
- Safi, B.; Saidi, M.; Daoui, A.; Bellal, A.; Mechekak, A.; Toumi, K. The use of seashells as a fine aggregate (by sand substitution) in self-compacting mortar (SCM). Constr. Build. Mater. 2015, 78, 430–438. [Google Scholar] [CrossRef]
- Bamigboye, G.; Enabulele, D.; Odetoyan, A.O.; Kareem, M.A.; Nworgu, A.; Bassey, D. Mechanical and durability assessment of concrete containing seashells: A review. Cogent Eng. 2021, 8, 1883830. [Google Scholar] [CrossRef]
- Kumar Sharma, N. Experimental study of concrete prepared by different waste products. Mater. Today Proc. 2021, 45, 3618–3624. [Google Scholar] [CrossRef]
- Tayeh, B.A.; Hasaniyah, M.W.; Zeyad, A.M.; Yusuf, M.O. Properties of concrete containing recycled seashells as cement partial replacement: A review. J. Clean. Prod. 2019, 237, 117723. [Google Scholar] [CrossRef]
- Maryoto, A.; Sthenly Gan, B.; Intang Setyo Hermanto, N.; Setijadi, R. Effect of Calcium Stearate in the Mechanical and Physical Properties of Concrete with PCC and Fly Ash as Binders. Materials 2020, 13, 1394. [Google Scholar] [CrossRef]
- White, M.M.; Chejlava, M.; Fried, B.; Sherma, J. The concentration of calcium carbonate in shells of freshwater snails. Am. Malacol. Bull. 2007, 22, 139–142. [Google Scholar] [CrossRef]
- Hart, A. Mini-review of waste shell-derived materials’ applications. Waste Manag. Res. 2020, 38, 514–527. [Google Scholar] [CrossRef]
- Owuamanam, S.; Cree, D. Progress of Bio-Calcium Carbonate Waste Eggshell and Seashell Fillers in Polymer Composites: A Review. J. Compos. Sci. 2020, 4, 70. [Google Scholar] [CrossRef]
- Bazargan, A.; Kostic, M.D.; Stamenković, O.S.; Veljković, V.B.; McKay, G. A calcium oxide-based catalyst derived from palm kernel shell gasification residues for biodiesel production. Fuel 2015, 150, 519–525. [Google Scholar] [CrossRef]
- Manso, S.; De Muynck, W.; Segura, I.; Aguado, A.; Steppe, K.; Boon, N.; De Belie, N. Bioreceptivity evaluation of cementitious materials designed to stimulate biological growth. Sci. Total Environ. 2014, 481, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Bone, J.R.; Stafford, R.; Hall, A.E.; Herbert, R.J.H. The intrinsic primary bioreceptivity of concrete in the coastal environment—A review. Dev. Built Environ. 2022, 10, 100078. [Google Scholar] [CrossRef]
- Jones, J.W.; Antle, J.M.; Basso, B.; Boote, K.J.; Conant, R.T.; Foster, I.; Godfray, H.C.J.; Herrero, M.; Howitt, R.E.; Janssen, S.; et al. Toward a new generation of agricultural system data, models, and knowledge products: State of agricultural systems science. Agric. Syst. 2017, 155, 269–288. [Google Scholar] [CrossRef]
- Mat Zin, M.I.; Jimat, D.N.; Wan Nawawi, W.M.F. Physicochemical properties of fungal chitin nanopaper from shiitake (L. edodes), enoki (F. velutipes) and oyster mushrooms (P. ostreatus). Carbohydr. Polym. 2022, 281, 119038. [Google Scholar] [CrossRef]
- Yang, L.; Park, D.; Qin, Z. Material Function of Mycelium-Based Bio-Composite: A Review. Front. Mater. 2021, 8, 737377. [Google Scholar] [CrossRef]
- Fricker, M.; Boddy, L.; Bebber, D. Network Organisation of Mycelial Fungi. In Biology of the Fungal Cell; Howard, R.J., Gow, N.A.R., Eds.; The Mycota; Springer: Berlin/Heidelberg, Germany, 2007; Volume 8. [Google Scholar] [CrossRef]
- Sejian, V.; Samal, L.; Haque, N.; Bagath, M.; Hyder, I.; Maurya, V.P.; Bhatta, R.; Ravindra, J.P.; Prasad, C.S.; Lal, R. Overview on Adaptation, Mitigation and Amelioration Strategies to Improve Livestock Production Under the Changing Climatic Scenario. Clim. Change Impact Livest. Adapt. Mitig. 2015, 359–397. [Google Scholar] [CrossRef]
- Holt, G.A.; Mcintyre, G.; Flagg, D.; Bayer, E.; Wanjura, J.D.; Pelletier, M.G. Fungal Mycelium and Cotton Plant Materials in the Manufacture of Biodegradable Molded Packaging Material: Evaluation Study of Select Blends of Cotton Byproducts. J. Biobased Mater. Bioenergy 2012, 6, 431–439. [Google Scholar] [CrossRef]
- Pelletier, M.G.; Holt, G.A.; Wanjura, J.D.; Bayer, E.; McIntyre, G. An evaluation study of mycelium based acoustic absorbers grown on agricultural by-product substrates. Ind. Crops Prod. 2013, 51, 480–485. [Google Scholar] [CrossRef]
- Haneef, M.; Ceseracciu, L.; Canale, C.; Bayer, I.S.; Heredia-Guerrero, J.A.; Athanassiou, A. Advanced Materials From Fungal Mycelium: Fabrication and Tuning of Physical Properties. Sci. Rep. 2017, 7, srep41292. [Google Scholar] [CrossRef]
- Islam, M.R.; Tudryn, G.; Bucinell, R.; Schadler, L.; Picu, R.C. Morphology and mechanics of fungal mycelium. Sci. Rep. 2017, 7, 13070. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Huynh, T.; Dekiwadia, C.; Daver, F.; John, S. Mycelium Composites: A Review of Engineering Characteristics and Growth Kinetics. J. Bionanosci. 2017, 11, 241–257. [Google Scholar] [CrossRef]
- Bansal, V.; Ahmad, A.; Sastry, M. Fungus-mediated Biotransformation of Amorphous Silica in rice Husk to Nanocrystalline Silica. J. Am. Chem. Soc. 2006, 128, 14059–14066. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Bhat, T.; Huynh, T.; Kandare, E.; Yuen, R.; Wang, C.H.; John, S. Waste-derived low-cost mycelium composite construction materials with improved fire safety. Fire Mater. 2018, 42, 816–825. [Google Scholar] [CrossRef]
- Maraveas, C. Production of Sustainable and Biodegradable Polymers from Agricultural Waste. Polymers 2020, 12, 1127. [Google Scholar] [CrossRef] [PubMed]
- Defonseka, C.P. Polymeric Composites with Rice Hulls; Walter de Gruyter: Berlin, Germany, 2019. [Google Scholar] [CrossRef]
- Girometta, C.; Picco, A.M.; Baiguera, R.M.; Dondi, D.; Babbini, S.; Cartabia, M.; Pellegrini, M.; Savino, E. Physico-mechanical and Thermodynamic Properties of Mycelium-Based Biocomposites: A Review. Sustainability 2019, 11, 281. [Google Scholar] [CrossRef]
- Appels, F.V.W.; Dijksterhuis, J.; Lukasiewicz, C.E.; Jansen, K.M.B.; Wösten, H.A.B.; Krijgsheld, P. Hydrophobin gene deletion and environmental growth conditions impact mechanical properties of mycelium by affecting the density of the material. Sci. Rep. 2018, 8, 4703. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S.; Nickerson, W.J. Isolation, composition, and structure of cell walls of filamentous and yeast-like forms of Mucor rouxii. Biochim. Biophys. Acta 1962, 58, 102–119. [Google Scholar] [CrossRef]
- Jiang, L.; Walczyk, D.; McIntyre, G.; Bucinell, R.; Tudryn, G. Manufacturing of biocomposite sandwich structures using mycelium-bound cores and preforms. J. Manuf. Process. 2017, 28, 50–59. [Google Scholar] [CrossRef]
- Xing, Y.; Brewer, M.; El-Gharabawy, H.; Griffith, G.; Jones, P. Growing and testing mycelium bricks as building insulation materials. IOP Conf. Ser. Earth Environ. Sci. 2018, 121, 022032. [Google Scholar] [CrossRef]
- Jiang, L.; Walczyk, D.; McIntyre, G.; Bucinell, R.; Li, B. Bioresin infused then cured mycelium-based sandwich-structure biocomposites: Resin transfer molding (RTM) process, flexural properties, and simulation. J. Clean. Prod. 2019, 207, 123–135. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Manan, S.; Ullah, M.W.; Ul-Islam, M.; Atta, O.M.; Yang, G. Synthesis and applications of fungal mycelium-based advanced functional materials. J. Bioresour. Bioprod. 2021, 6, 1–10. [Google Scholar] [CrossRef]
- Sydor, M.; Cofta, G.; Doczekalska, B.; Bonenberg, A. Fungi in Mycelium-Based Composites: Usage and Recommendations. Materials 2022, 15, 6283. [Google Scholar] [CrossRef]
- Soh, E.; Le Ferrand, H. Woodpile structural designs to increase the stiffness of mycelium-bound composites. Mater. Des. 2023, 225, 111530. [Google Scholar] [CrossRef]
- Jauk, J.; Gosch, L.; Vašatko, H.; Christian, I.; Klaus, A.; Stavric, M. MyCera. Application of mycelial growth within digitally manufactured clay structures. Int. J. Archit. Comput. 2022, 20, 31–40. [Google Scholar] [CrossRef]
- Özdemir, E.; Saeidi, N.; Javadian, A.; Rossi, A.; Nolte, N.; Ren, S.; Dwan, A.; Acosta, I.; Hebel, D.E.; Wurm, J.; et al. Wood-Veneer-Reinforced Mycelium Composites for Sustainable Building Components. Biomimetics 2022, 7, 39. [Google Scholar] [CrossRef]
- Blast Studio. Tree Column. 2020. Available online: https://www.blast-studio.com/tree-column (accessed on 17 May 2023).
- Bitting, S.; Derme, T.; Lee, J.; Van Mele, T.; Dillenburger, B.; Block, P. Challenges and Opportunities in Scaling up Architectural Applications of Mycelium-Based Materials with Digital Fabrication. Biomimetics 2022, 7, 44. [Google Scholar] [CrossRef]
- Modanloo, B.; Ghazvinian, A.; Matini, M.; Andaroodi, E. Tilted Arch; Implementation of Additive Manufacturing and Bio-Welding of Mycelium-Based Composites. Biomimetics 2021, 6, 68. [Google Scholar] [CrossRef]
- Goidea, A.; Dimitrios Floudas and Andréen, D. Pulp Faction: 3d printed material assemblies through microbial biotransformation. In Fabricate 2020; UCL Press: London, UK, 2020; pp. 42–49. [Google Scholar]
- Elsacker, E.; Vandelook, S.; Van Wylick, A.; Ruytinx, J.; De Laet, L.; Peeters, E. A comprehensive framework for the production of mycelium-based lignocellulosic composites. Sci. Total Environ. 2020, 725, 138431. [Google Scholar] [CrossRef] [PubMed]
- Souza, E. Mushroom Buildings? The Possibilities of Using Mycelium in Architecture. ArchDaily. 2020. Available online: https://www.archdaily.com/949007/mushroom-buildings-the-possibilities-of-using-mycelium-in-architecture (accessed on 5 July 2023).
- Dahy, H. Biocomposite materials based on annual natural fibers and biopolymers—Design, fabrication and customized applications in architecture. Constr. Build. Mater. 2017, 147, 212–220. [Google Scholar] [CrossRef]
- fcl.ethz.ch. Mycelium-Bound Biocomposites as Potential Sustainable Particleboards. 2021. Available online: https://fcl.ethz.ch/news-events/news/2021/11/mycelium-bound-biocomposite-a-sustainable-replacement-for-particleboards.html (accessed on 9 August 2023).
- ArchDaily. Mycelium Materials: The Future of Growing Our Homes. 2022. Available online: https://www.archdaily.com/985570/mycelium-materials-the-future-of-growing-our-homes (accessed on 5 July 2023).
- Elsacker, E.; Vandelook, S.; Brancart, J.; Peeters, E.; De Laet, L. Mechanical, physical and chemical characterisation of mycelium-based composites with different types of lignocellulosic substrates. PLoS ONE 2019, 14, e0213954. [Google Scholar] [CrossRef] [PubMed]
- Houette, T.; Maurer, C.; Niewiarowski, R.; Gruber, P. Growth and Mechanical Characterization of Mycelium-Based Composites towards Future Bioremediation and Food Production in the Material Manufacturing Cycle. Biomimetics 2022, 7, 103. [Google Scholar] [CrossRef]
- Muiruri, J.K.; Yeo, J.C.C.; Zhu, Q.; Ye, E.; Loh, X.J.; Li, Z. Sustainable Mycelium-Bound Biocomposites: Design Strategies, Materials Properties, and Emerging Applications. ACS Sustain. Chem. Eng. 2023, 11, 6801–6821. [Google Scholar] [CrossRef]
- Gibson, L.J. The hierarchical structure and mechanics of plant materials. J. R. Soc. Interface 2012, 9, 2749–2766. [Google Scholar] [CrossRef]
- Arifin, Y.H.; Yusuf, Y. Mycelium Fibers as New Resource for Environmental Sustainability. Procedia Eng. 2013, 53, 504–508. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Green synthesis approach: Extraction of chitosan from fungus mycelia. Crit. Rev. Biotechnol. 2012, 33, 379–403. [Google Scholar] [CrossRef]
- López-Nava, J.A.; Méndez González, J.; Ruelas Chacón, X.; Nájera Luna, J.A. Assessment of Edible Fungi and Films Bio-Based Material Simulating Expanded Polystyrene. Mater. Manuf. Process. 2015, 31, 1085–1090. [Google Scholar] [CrossRef]
- Appels, F.V.; Camere, S.; Montalti, M.; Karana, E.; Jansen, K.M.; Dijksterhuis, J.; Krijgsheld, P.; Wösten, H.A. Fabrication Factors Influencing Mechanical, Moisture- and Water-Related Properties of Mycelium-Based Composites. Mater. Des. 2019, 161, 64–71. [Google Scholar] [CrossRef]
- Irbe, I.; Loris, G.D.; Filipova, I.; Andze, L.; Skute, M. Characterization of Self-Growing Biomaterials Made of Fungal Mycelium and Various Lignocellulose-Containing Ingredients. Materials 2022, 15, 7608. [Google Scholar] [CrossRef]
- Sun, W.; Tajvidi, M.; Hunt, C.G.; McIntyre, G.; Gardner, D.J. Fully Bio-Based Hybrid Composites Made of Wood, Fungal Mycelium and Cellulose Nanofibrils. Sci. Rep. 2019, 9, 3766. [Google Scholar] [CrossRef] [PubMed]
- Short-Term Outlook Report: War in Ukraine Continues to Impact EU Farmers. Available online: https://agriculture.ec.europa.eu/news/short-term-outlook-report-war-ukraine-continues-impact-eu-farmers-2023-03-30_en (accessed on 15 July 2023).
- Agriculture and Food Security: Casualties of the War in Ukraine. 2022. Available online: https://www.csis.org/analysis/agriculture-and-food-security-casualties-war-ukraine (accessed on 15 July 2023).
- Topić Popović, N.; Lorencin, V.; Strunjak-Perović, I.; Čož-Rakovac, R. Shell Waste Management and Utilization: Mitigating Organic Pollution and Enhancing Sustainability. Appl. Sci. 2023, 13, 623. [Google Scholar] [CrossRef]
- Yao, Z.; Xia, M.; Li, H.; Chen, T.; Ye, Y.; Zheng, H. Bivalve Shell: Not an Abundant Useless Waste but a Functional and Versatile Biomaterial. Crit. Rev. Environ. Sci. Technol. 2014, 44, 2502–2530. [Google Scholar] [CrossRef]
- GLOBEFISH | Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/in-action/globefish/publications/details-publication/en/c/347665 (accessed on 9 August 2023).
- Yoon, G.-L.; Kim, B.-T.; Kim, B.-O.; Han, S.-H. Chemical–mechanical characteristics of crushed oyster-shell. Waste Manag. 2003, 23, 825–834. [Google Scholar] [CrossRef]
- Suarez-Riera, D.; Merlo, A.; Lavagna, L.; Nisticò, R.; Pavese, M. Mechanical properties of mortar containing recycled Acanthocardia tuberculata seashells as aggregate partial replacement. Boletín Soc. Española Cerámica Vidr. 2021, 60, 206–210. [Google Scholar] [CrossRef]
- Mo, K.H.; Alengaram, U.J.; Jumaat, M.Z.; Lee, S.C.; Goh, W.I.; Yuen, C.W. Recycling of seashell waste in concrete: A review. Constr. Build. Mater. 2018, 162, 751–764. [Google Scholar] [CrossRef]
- Hamester, M.R.R.; Balzer, P.S.; Becker, D. Characterization of calcium carbonate obtained from oyster and mussel shells and incorporation in polypropylene. Mater. Res. 2012, 15, 204–208. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Hashimoto, S. Cold sintering of calcium carbonate derived from seashells. Open Ceram. 2022, 12, 100302. [Google Scholar] [CrossRef]
- Alabaraoye, E.; Achilonu, M.; Hester, R. Biopolymer (Chitin) from Various Marine Seashell Wastes: Isolation and Characterization. J. Polym. Environ. 2017, 26, 2207–2218. [Google Scholar] [CrossRef]
- Li, Q.; Dunn, E.T.; Grandmaison, E.W.; Goosen, M.F.A. Applications and Properties of Chitosan. J. Bioact. Compat. Polym. 1992, 7, 370–397. [Google Scholar] [CrossRef]
- Leone, R.; Calà, A.; Capela, M.N.; Colajanni, S.; Campisi, T.; Saeli, M. Recycling Mussel Shells as Secondary Sources in Green Construction Materials: A Preliminary Assessment. Sustainability 2023, 15, 3547. [Google Scholar] [CrossRef]
- Li, T.; Xin, R.; Wang, D.; Yuan, L.; Wu, D.; Wu, X. Research Progress on the Applications of Seashell Adsorption Behaviors in Cement-Based Materials. Buildings 2023, 13, 1289. [Google Scholar] [CrossRef]
- Attias, N.; Danai, O.; Abitbol, T.; Tarazi, E.; Ezov, N.; Pereman, I.; Grobman, Y.J. Mycelium Bio-Composites in Industrial Design and Architecture: Comparative Review and Experimental Analysis. J. Clean. Prod. 2020, 246, 119037. [Google Scholar] [CrossRef]
- Islam, M.R.; Omar, M.; Pk, M.M.U.; Mia, R. Phytochemicals and Antibacterial Activity Screening of Three Edible Mushrooms Pleurotus Ostreatus, Ganoderma Lucidum and Lentinula Edodes Accessible in Bangladesh. Am. J. Biol. Life Sci. 2015, 3, 31–35. [Google Scholar]
- Petre, M. Mushroom Biotechnology: Developments and Applications; Academic Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Jones, M.; Mautner, A.; Luenco, S.; Bismarck, A.; John, S. Engineered Mycelium Composite Construction Materials from Fungal Biorefineries: A Critical Review. Mater. Des. 2020, 187, 108397. [Google Scholar] [CrossRef]
- Corner, E.J.H. The construction of polypores. 1. Introduction-polyporus-sulphureus, p-squamosus, p-betulinus and polystictus-microcyclus. Phytomorphology 1953, 3, 152–167. [Google Scholar]
- Ghareeb, B.A. Impact different Level of Calcium Carbonate (CaCo3) on Growth and yield of Oyster Mushroom (Pleurotus spp.). Int. J. Eng. Technol. 2019, 11, 785–792. [Google Scholar] [CrossRef]
- Choi, U.-K.; Lee, O.-H.; Kim, Y.-C. Effect of Calcinated Oyster Shell Powder on Growth, Yield, Spawn Run, and Primordial Formation of King Oyster Mushroom (Pleurotus Eryngii). Molecules 2011, 16, 2313–2322. [Google Scholar] [CrossRef]
- Tang, Z.-X.; Shi, L.-E.; Jiang, Z.-B.; Bai, X.-L.; Ying, R.-F. Calcium Enrichment in Edible Mushrooms: A Review. J. Fungi 2023, 9, 338. [Google Scholar] [CrossRef]
- Zhang, R. Functional characterization of cellulose-degrading AA9 lytic polysaccharide monooxygenases and their potential exploitation. Appl. Microbiol. Biotechnol. 2020, 104, 3229–3243. [Google Scholar] [CrossRef] [PubMed]
- Oloye, O.; O’Mullane, A.P. Electrochemical Capture and Storage of CO2 as Calcium Carbonate. ChemSusChem 2021, 14, 1767–1775. [Google Scholar] [CrossRef] [PubMed]
- Babbar, S.B.; Jain, R. Xanthan Gum: An Economical Partial Substitute for Agar in Microbial Culture Media. Curr. Microbiol. 2006, 52, 287–292. [Google Scholar] [CrossRef]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [CrossRef]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Drozdowski, L.A.; Reimer, R.A.; Temelli, F.; Bell, R.C.; Vasanthan, T.; Thomson, A.B.R. β-Glucan extracts inhibit the in vitro intestinal uptake of long-chain fatty acids and cholesterol and down-regulate genes involved in lipogenesis and lipid transport in rats. J. Nutr. Biochem. 2010, 21, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Lau, C.C.; Ismail, S.M. Potential use of Lentinus squarrosulus mushroom as fermenting agent and source of natural antioxidant additive in livestock feed. J. Sci. Food Agric. 2015, 96, 1459–1466. [Google Scholar] [CrossRef]
- Miles, P.G.; Chang, S.T. Mushroom Biology: Concise Basics and Current Developments, 1st ed.; World Scientific: Singapore, 1997. [Google Scholar]
- Ortega, G.M.; Martinez, E.O.; Betancourt, D.; Gonzaléz, A.E.; Otero, M.A. Bioconversion of sugar cane crop residues with white-rot fungi Pleurotus sp. World J. Microbiol. Biotechnol. 1992, 8, 402–405. [Google Scholar] [CrossRef]
- Sturion, G.L.; Oetterer, M. Utilização da folha da bananeira como substrato para cultivo de cogumelos comestíveis (Pleurotus spp.). Ciênc. Tecnol. Alimen. 1995, 15, 194–200. [Google Scholar]
- Reitsma, J.; Murphy, D.C.; Archer, A.F.; York, R.H. Nitrogen extraction potential of wild and cultured bivalves harvested from nearshore waters of Cape Cod, USA. Mar. Pollut. Bull. 2017, 116, 175–181. [Google Scholar] [CrossRef]
- Pyeon, H.-Y.; Park, Y.-J.; Lee, S.-C.; Jang, M.-J. Transcriptomic Analysis of Salt Stress Response in Pleurotus ostreatus. Horticulturae 2022, 8, 1147. [Google Scholar] [CrossRef]
- Adebayo-Tayo, B.C. Optimization of growth conditions for mycelial yield and exopolysaccharride production by Pleurotus ostreatus cultivated in Nigeria. Afr. J. Microbiol. Res. 2011, 5, 2130–2138. [Google Scholar] [CrossRef]
- Rozsa, S.; Măniuțiu, D.-N.; Lazăr, V.; Gocan, T.-M.; Andreica, I. The Influence of pH and the Source of Nitrogen on the Mycelial Growth of the Pleurotus ostreatus. Mushrooms. Uaiasi.ro. 2016. Available online: https://repository.uaiasi.ro/xmlui/handle/20.500.12811/1386 (accessed on 26 July 2023).
- Hoa, H.T.; Wang, C.-L. The Effects of Temperature and Nutritional Conditions on Mycelium Growth of Two Oyster Mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.R.; Bajwa, S.G.; Holt, G.A.; McIntyre, G.; Bajwa, D.S. Evaluation of Physico-Mechanical Properties of Mycelium Reinforced green Biocomposites Made from Cellulosic Fibers. Appl. Eng. Agric. 2016, 32, 931–938. [Google Scholar] [CrossRef]
- Dai, C.; Yu, C.; Xu, C.; He, G. Heat and Mass Transfer in wood Composite Panels during Hot Pressing: Part 4. Experimental Investigation and Model Validation. Wood Fiber Sci. 2007, 61, 83–88. [Google Scholar] [CrossRef]
- Qin, Z.; Jung, G.S.; Kang, M.J.; Buehler, M.J. The Mechanics and Design of a Lightweight Three-Dimensional Graphene Assembly. Sci. Adv. 2017, 3, e1601536. [Google Scholar] [CrossRef]
- Butterfield, B.; Chapman, K.; Christie, L. Ultrastructural Characteristics of Failure Surfaces in Medium Density Fiberboard. For. Prod. J. 1992, 42, 55–60. [Google Scholar]
- Carvalho, L.M.H.; Costa, C.A.V. Modeling and Simulation of the Hot-Pressing Process in the Production of Medium Density Fiberboard (MDF). Chem. Eng. Commun. 1998, 170, 1–21. [Google Scholar] [CrossRef]
- Karana, E.; Blauwhoff, D.; Hultink, E.J.; Camere, S. When the Material Grows: A Case Study on Designing (With) Mycelium-Based Materials. Int. J. Des. 2018, 12, 119–136. [Google Scholar]
- Di Mario, F.; Rapanà, P.; Tomati, U.; Galli, E. Chitin and Chitosan from Basidiomycetes. Int. J. Biol. Macromol. 2008, 43, 8–12. [Google Scholar] [CrossRef]
- Hassainia, A.; Satha, H.; Boufi, S. Chitin from Agaricus Bisporus: Extraction and Characterization. Int. J. Biol. Macromol. 2018, 117, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Azimi, B.; Ricci, C.; Fusco, A.; Zavagna, L.; Linari, S.; Donnarumma, G.; Hadrich, A.; Cinelli, P.; Coltelli, M.-B.; Danti, S.; et al. Electrosprayed Shrimp and Mushroom Nanochitins on Cellulose Tissue for Skin Contact Application. Molecules 2021, 26, 4374. [Google Scholar] [CrossRef] [PubMed]
- Nawawi, W.M.F.W.; Lee, K.-Y.; Kontturi, E.; Murphy, R.J.; Bismarck, A. Chitin Nanopaper from Mushroom Extract: Natural Composite of Nanofibers and Glucan from a Single Biobased Source. ACS Sustain. Chem. Eng. 2019, 7, 6492–6496. [Google Scholar] [CrossRef]
- Maximino, C.; Ongpeng, J.; Inciong, E.; Sendo, V.; Soliman, C.; Siggaoat, A. Using Waste in Producing Bio-Composite Mycelium Bricks. Appl. Sci. 2020, 10, 5303. [Google Scholar] [CrossRef]
- Erjavec, J.; Kos, J.; Ravnikar, M.; Dreo, T.; Sabotič, J. Proteins of Higher Fungi—from forest to Application. Trends Biotechnol. 2012, 30, 259–273. [Google Scholar] [CrossRef]
- Baldrian, P.; Valášková, V. Degradation of Cellulose by Basidiomycetous Fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef]
- Hatakka, A.; Hammel, K.E. Fungal Biodegradation of Lignocelluloses. In Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2011; pp. 319–340. [Google Scholar] [CrossRef]
- Sánchez, C. Lignocellulosic Residues: Biodegradation and Bioconversion by Fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose Degradation: An Overview of Fungi and Fungal Enzymes Involved in Lignocellulose Degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Whiteford, J.R.; Spanu, P.D. Hydrophobins and the Interactions between Fungi and Plants. Mol. Plant Pathol. 2002, 3, 391–400. [Google Scholar] [CrossRef]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setälä, T.; Penttilä, M.E. Hydrophobins: The Protein-Amphiphiles of Filamentous Fungi. FEMS Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Ortiz-Castellanos, L. Cell wall Glucans of Fungi. A Review. Cel Surf. 2019, 5, 100022. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 Million Species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, D.S.; Holt, G.A.; Bajwa, S.G.; Duke, S.E.; McIntyre, G. Enhancement of Termite (Reticulitermes flavipes L.) Resistance in Mycelium Reinforced Biofiber-Composites. Ind. Crops Prod. 2017, 107, 420–426. [Google Scholar] [CrossRef]
- Shumway, S.E.; Parsons, G.J. Scallops: Biology, Ecology, Aquaculture, and Fisheries; Elsevier Science: Oxford, UK, 2016. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdallah, Y.K.; Estévez, A.T. Biowelding 3D-Printed Biodigital Brick of Seashell-Based Biocomposite by Pleurotus ostreatus Mycelium. Biomimetics 2023, 8, 504. https://doi.org/10.3390/biomimetics8060504
Abdallah YK, Estévez AT. Biowelding 3D-Printed Biodigital Brick of Seashell-Based Biocomposite by Pleurotus ostreatus Mycelium. Biomimetics. 2023; 8(6):504. https://doi.org/10.3390/biomimetics8060504
Chicago/Turabian StyleAbdallah, Yomna K., and Alberto T. Estévez. 2023. "Biowelding 3D-Printed Biodigital Brick of Seashell-Based Biocomposite by Pleurotus ostreatus Mycelium" Biomimetics 8, no. 6: 504. https://doi.org/10.3390/biomimetics8060504
APA StyleAbdallah, Y. K., & Estévez, A. T. (2023). Biowelding 3D-Printed Biodigital Brick of Seashell-Based Biocomposite by Pleurotus ostreatus Mycelium. Biomimetics, 8(6), 504. https://doi.org/10.3390/biomimetics8060504