
Biomimetic Collagen Membranes as Drug Carriers of Geranylgeraniol to Counteract the Effect of Zoledronate
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Methods
2.1. Collagen Membranes Functionalization
2.2. Cell Culture
2.3. Cell Proliferation Assay
2.4. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5. Nodules Formation and Matrix Mineralization
2.6. Scanning Electron Microscopy (SEM)
2.7. Statistical Analysis
3. Results
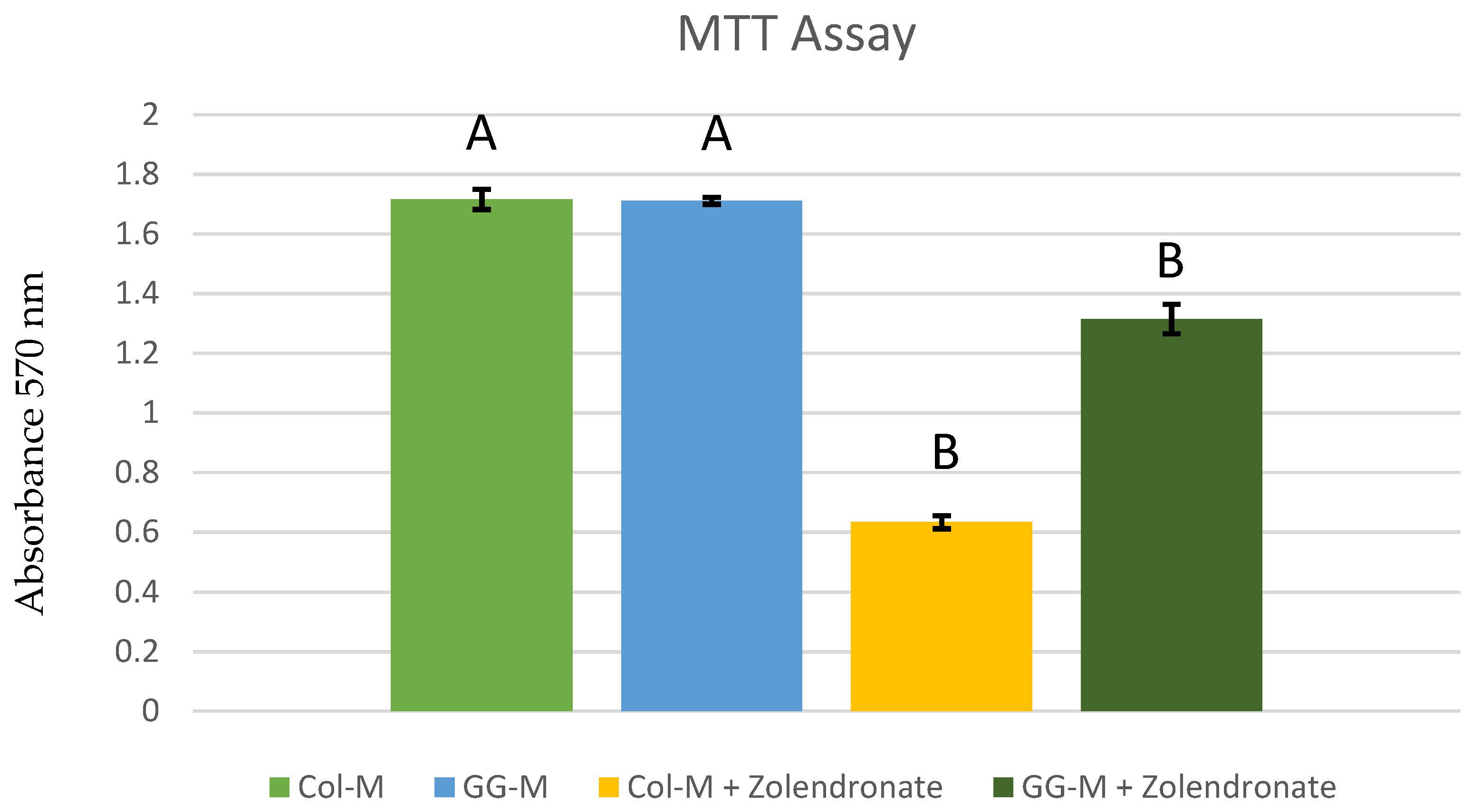
3.1. Cell Proliferation Assay
3.2. Real-Time Quantitative Polymerase Chain Reaction
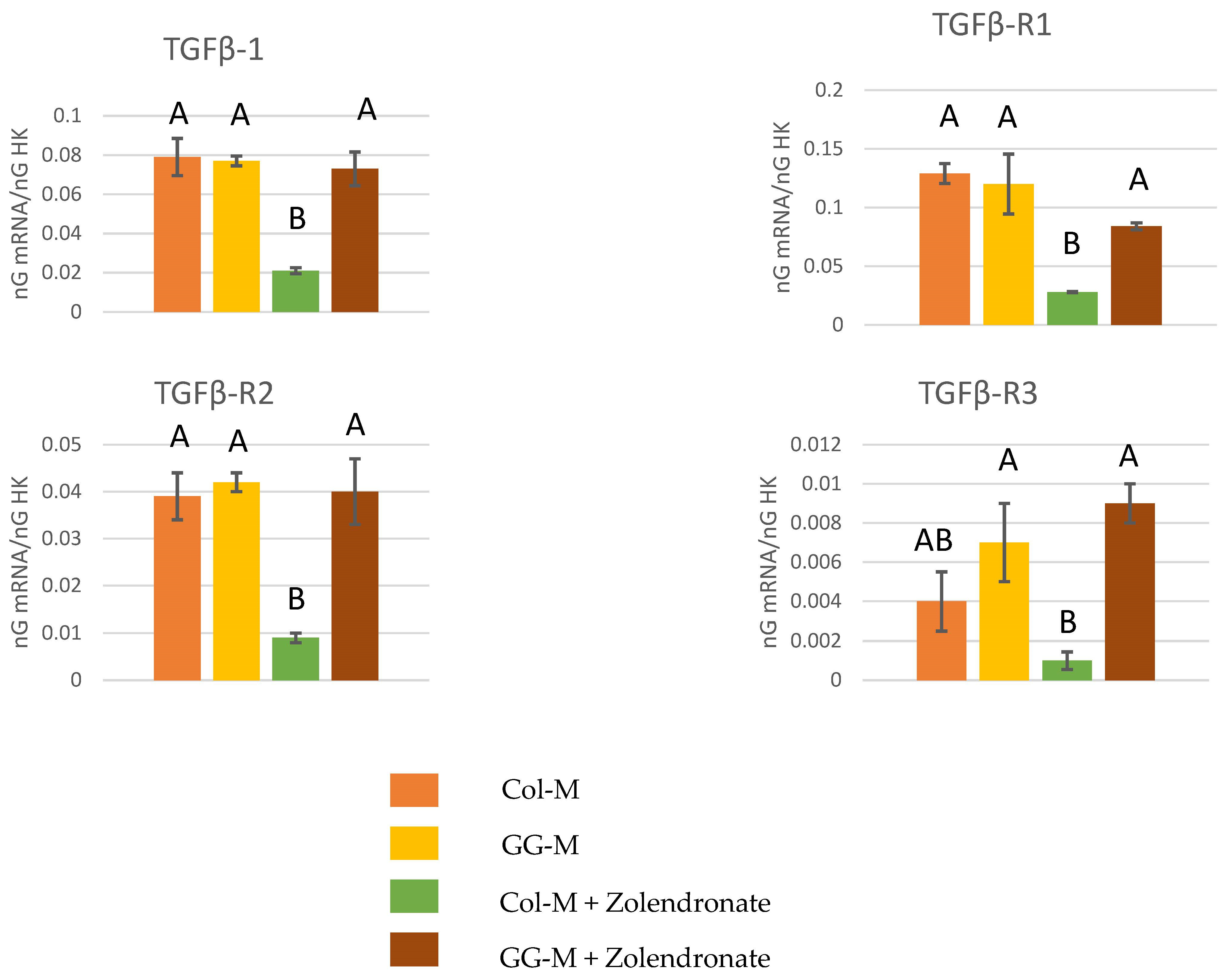
3.2.1. Gene Expression of TGF-β1 and Its Receptors (TGF-β R1, TGF-β R2, and TGF-β R3)
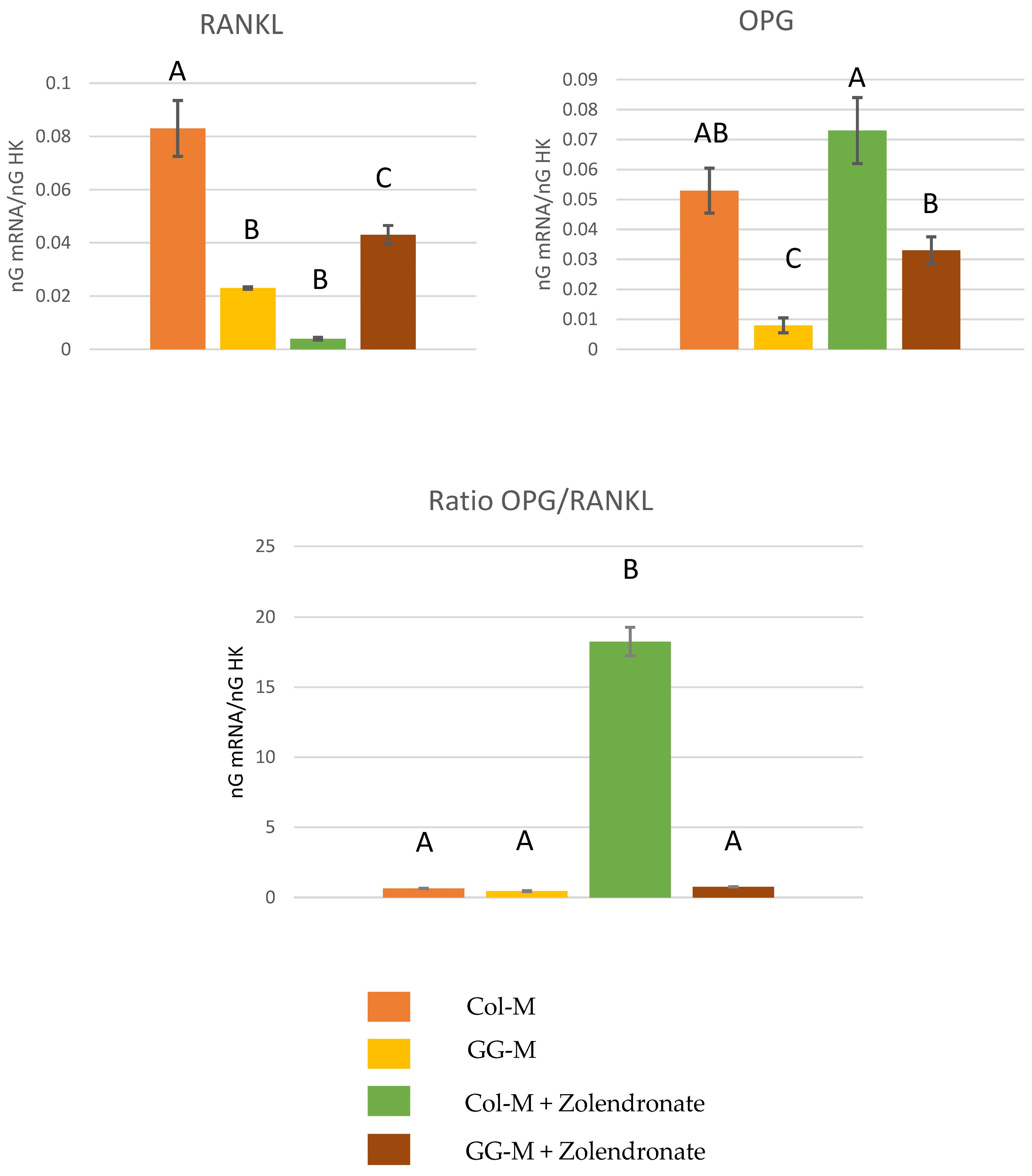
3.2.2. Gene Expression of OPG-RANKL Complex
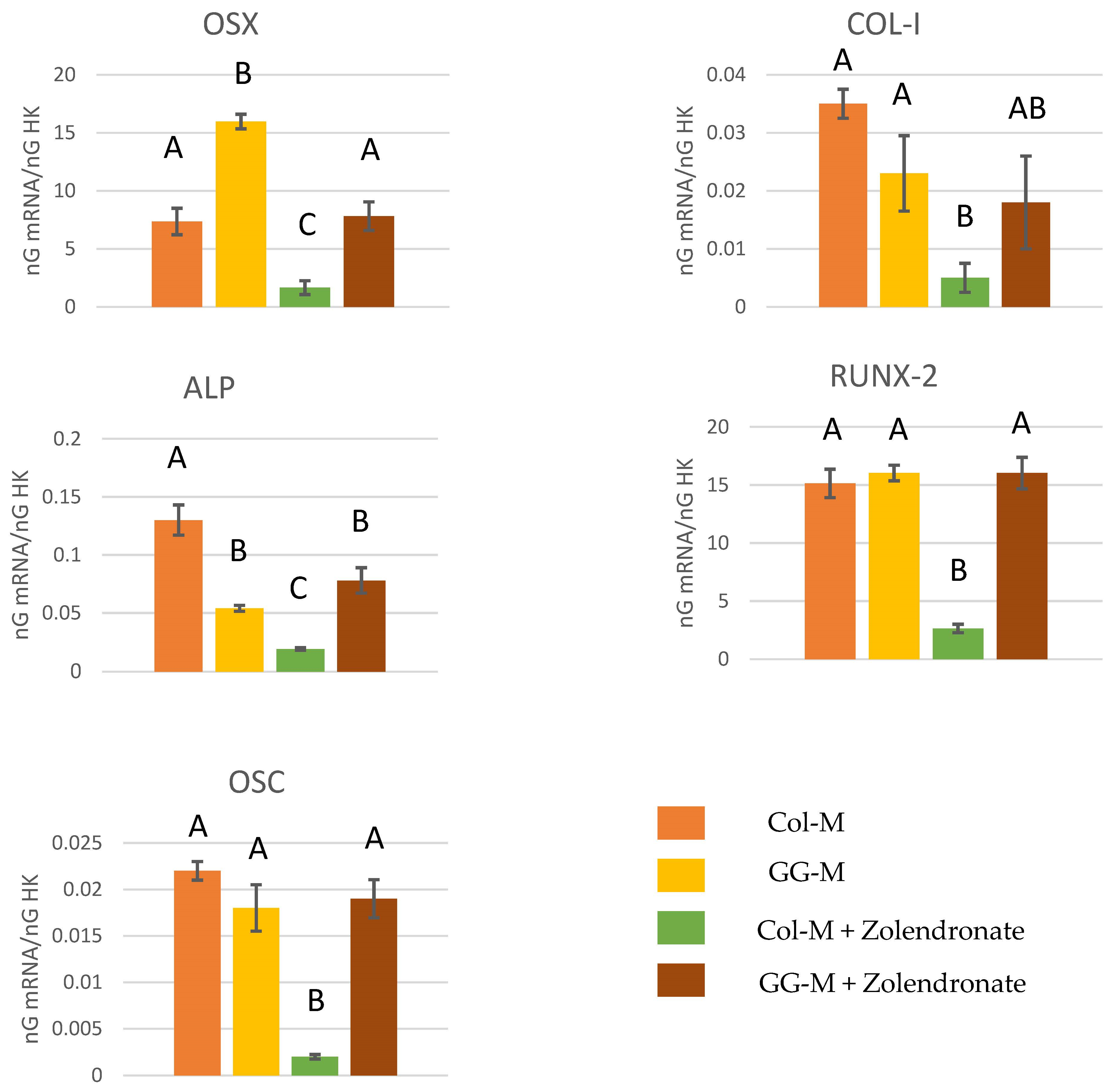
3.2.3. Effect of BPs on the Gene Expression of Runx2, ALP, Col-I, OSX, and OSC
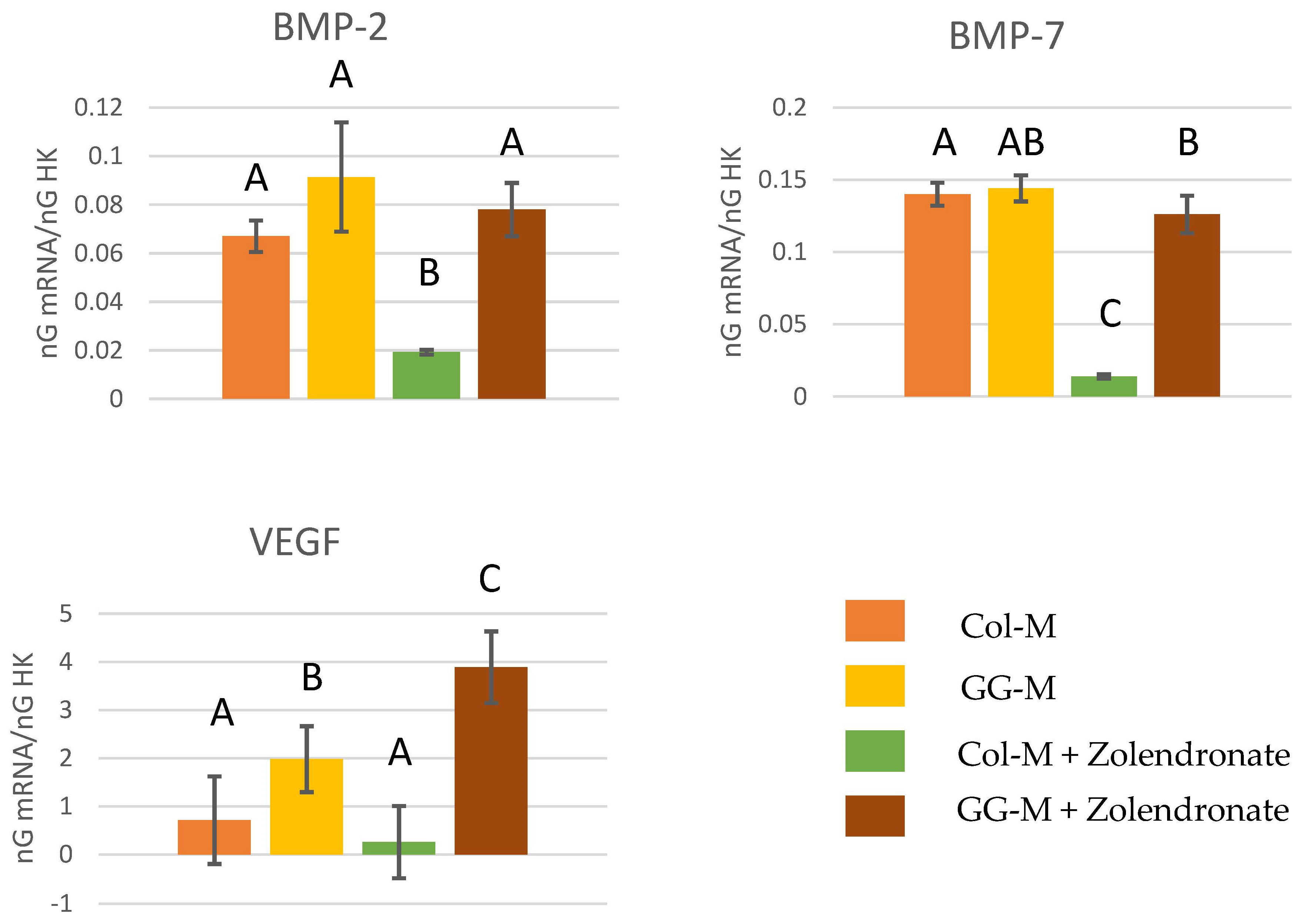
3.2.4. Gene Expression of BMP-2, BMP-7, and VEGF
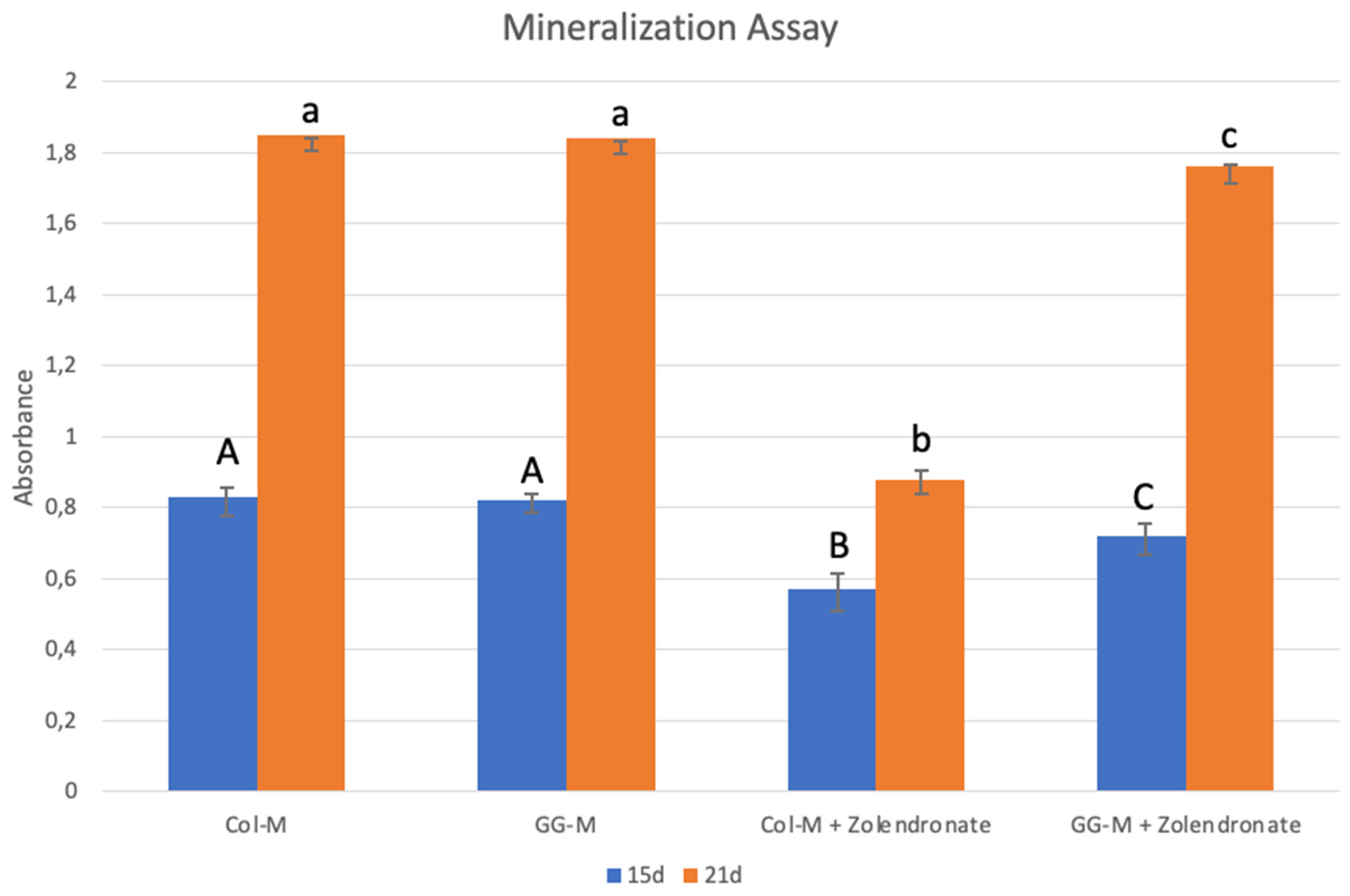
3.3. Nodules Formation and Matrix Mineralization
3.4. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zagho, M.M.; Hussein, E.A.; Elzatahry, A.A. Recent Overviews in Functional Polymer Composites for Biomedical Applications. Polymers 2018, 10, 739. [Google Scholar] [CrossRef]
- Li, H.; Xu, M.; Shi, R.; Zhang, A.; Zhang, J. Advances in Electrostatic Spinning of Polymer Fibers Functionalized with Metal-Based Nanocrystals and Biomedical Applications. Molecules 2022, 27, 5548. [Google Scholar] [CrossRef] [PubMed]
- Abou Neel, E.A.; Bozec, L.; Knowles, J.C.; Syed, O.; Mudera, V.; Day, R.; Hyun, J.K. Collagen—Emerging Collagen Based Therapies Hit the Patient. Adv. Drug Deliv. Rev. 2013, 65, 429–456. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Liang, Y.; Liu, X. Biomaterials for Periodontal Regeneration. Dent. Clin. N. Am. 2022, 66, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Radu, E.R.; Semenescu, A.; Voicu, S.I. Recent Advances in Stimuli-Responsive Doxorubicin Delivery Systems for Liver Cancer Therapy. Polymers 2022, 14, 5249. [Google Scholar] [CrossRef] [PubMed]
- Abbasnezhad, N.; Kebdani, M.; Shirinbayan, M.; Champmartin, S.; Tcharkhtchi, A.; Kouidri, S.; Bakir, F. Development of a Model Based on Physical Mechanisms for the Explanation of Drug Release: Application to Diclofenac Release from Polyurethane Films. Polymers 2021, 13, 1230. [Google Scholar] [CrossRef] [PubMed]
- Bornert, F.; Herber, V.; Sandgren, R.; Witek, L.; Coelho, P.G.; Pippenger, B.E.; Shahdad, S. Comparative Barrier Membrane Degradation over Time: Pericardium versus Dermal Membranes. Clin. Exp. Dent. Res. 2021, 7, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Toledano-Osorio, M.; de Luna-Bertos, E.; Toledano, M.; Manzano-Moreno, F.J.; García-Recio, E.; Ruiz, C.; Osorio, R.; Sanz, M. Doxycycline-Doped Collagen Membranes Accelerate in Vitro Osteoblast Proliferation and Differentiation. J. Periodontal Res. 2023, 58, 296–307. [Google Scholar] [CrossRef]
- Toledano-Osorio, M.; Vallecillo, C.; Vallecillo-Rivas, M.; Manzano-Moreno, F.-J.; Osorio, R. Antibiotic-Loaded Polymeric Barrier Membranes for Guided Bone/Tissue Regeneration: A Mini-Review. Polymers 2022, 14, 840. [Google Scholar] [CrossRef]
- Lane, J.M.; Khan, S.N.; O’Connor, W.J.; Nydick, M.; Hommen, J.P.; Schneider, R.; Tomin, E.; Brand, J.; Curtin, J. Bisphosphonate Therapy in Fibrous Dysplasia. Clin. Orthop. Relat. Res. 2001, 382, 6–12. [Google Scholar] [CrossRef]
- Lala, R.; Matarazzo, P.; Bertelloni, S.; Buzi, F.; Rigon, F.; de Sanctis, C. Pamidronate Treatment of Bone Fibrous Dysplasia in Nine Children with McCune-Albright Syndrome. Acta Paediatr. 2000, 89, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic Approaches to Bone Diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Patntirapong, S.; Singhatanadgit, W.; Chanruangvanit, C.; Lavanrattanakul, K.; Satravaha, Y. Zoledronic Acid Suppresses Mineralization through Direct Cytotoxicity and Osteoblast Differentiation Inhibition. J. Oral. Pathol. Med. 2012, 41, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Reszka, A.A.; Rodan, G.A. Nitrogen-Containing Bisphosphonate Mechanism of Action. Mini Rev. Med. Chem. 2004, 4, 711–719. [Google Scholar] [PubMed]
- Russell, R.G.G. Bisphosphonates: Mode of Action and Pharmacology. Pediatrics 2007, 119 (Suppl. S2), S150–S162. [Google Scholar] [CrossRef] [PubMed]
- Silverman, S.L.; Maricic, M. Recent Developments in Bisphosphonate Therapy. Semin. Arthritis Rheum. 2007, 37, 1–12. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Ramos-Torrecillas, J.; De Luna-Bertos, E.; Ruiz, C.; García-Martínez, O. High Doses of Bisphosphonates Reduce Osteoblast-like Cell Proliferation by Arresting the Cell Cycle and Inducing Apoptosis. J. Cranio-Maxillofac. Surg. 2015, 43, 396–401. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Ramos-Torrecillas, J.; De Luna-Bertos, E.; Reyes-Botella, C.; Ruiz, C.; García-Martínez, O. Nitrogen-Containing Bisphosphonates Modulate the Antigenic Profile and Inhibit the Maturation and Biomineralization Potential of Osteoblast-like Cells. Clin. Oral. Investig. 2015, 19, 895–902. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Ramos-Torrecillas, J.; Melguizo-Rodríguez, L.; Illescas-Montes, R.; Ruiz, C.; García-Martínez, O. Bisphosphonate Modulation of the Gene Expression of Different Markers Involved in Osteoblast Physiology: Possible Implications in Bisphosphonate-Related Osteonecrosis of the Jaw. Int. J. Med. Sci. 2018, 15, 359–367. [Google Scholar] [CrossRef]
- Marx, R.E. Pamidronate (Aredia) and Zoledronate (Zometa) Induced Avascular Necrosis of the Jaws: A Growing Epidemic. J. Oral. Maxillofac. Surg. 2003, 61, 1115–1117. [Google Scholar] [CrossRef]
- Ruggiero, S.L.; Mehrotra, B.; Rosenberg, T.J.; Engroff, S.L. Osteonecrosis of the Jaws Associated with the Use of Bisphosphonates: A Review of 63 Cases. J. Oral. Maxillofac. Surg. 2004, 62, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Melcher, A.H. On the Repair Potential of Periodontal Tissues. J. Periodontol. 1976, 47, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Dahlin, C.; Apatzidou, D.; Artzi, Z.; Bozic, D.; Calciolari, E.; De Bruyn, H.; Dommisch, H.; Donos, N.; Eickholz, P.; et al. Biomaterials and Regenerative Technologies Used in Bone Regeneration in the Craniomaxillofacial Region: Consensus Report of Group 2 of the 15th European Workshop on Periodontology on Bone Regeneration. J. Clin. Periodontol. 2019, 46 (Suppl. S21), 82–91. [Google Scholar] [CrossRef] [PubMed]
- Otto, M.; Lux, C.; Schlittenbauer, T.; Halling, F.; Ziebart, T. Geranyl-Geraniol Addition Affects Potency of Bisphosphonates-a Comparison in Vitro Promising a Therapeutic Approach for Bisphosphonate-Associated Osteonecrosis of the Jaw and Oral Wound Healing. Oral. Maxillofac. Surg. 2022, 26, 321–332. [Google Scholar] [CrossRef]
- Patntirapong, S.; Chanruangvanit, C.; Lavanrattanakul, K.; Satravaha, Y. Assessment of Bisphosphonate Treated-Osteoblast Behaviors by Conventional Assays and a Simple Digital Image Analysis. Acta Histochem. 2021, 123, 151659. [Google Scholar] [CrossRef]
- Thavornyutikarn, B.; Sungkhaphan, P.; Kaewkong, P.; Pornsuwan, S.; Risangud, N.; Singhatanadgit, W.; Janvikul, W. Biodegradable Dual-Function Nanocomposite Hydrogels for Prevention of Bisphosphonate-Related Osteonecrosis of the Jaw. ACS Appl. Bio Mater. 2023, 6, 1658–1675. [Google Scholar] [CrossRef]
- PubChem Geranylgeraniol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281365 (accessed on 31 May 2023).
- Latifi, L.; Sohrabnezhad, S.H.; Hadavi, M. Mesoporous Silica as a Support for Poorly Soluble Drug: Influence of pH and Amino Group on the Drug Release. Microporous Mesoporous Mater. 2017, 250, 148–157. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Ramos-Torrecillas, J.; De Luna-Bertos, E.; Reyes-Botella, C.; García-Martínez, O.; Ruiz, C. Effect of Clodronate on Antigenic Profile, Growth, and Differentiation of Osteoblast-Like Cells. J. Oral. Maxillofac. Surg. 2016, 74, 1765–1770. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Rodríguez-Martínez, J.B.; Ramos-Torrecillas, J.; Vallecillo-Capilla, M.F.; Ruiz, C.; García-Martínez, O.; Reyes-Botella, C. Proliferation and Osteogenic Differentiation of Osteoblast-like Cells Obtained from Two Techniques for Harvesting Intraoral Bone Grafts. Clin. Oral. Investig. 2013, 17, 1349–1356. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Ragni, E.; Viganò, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What Is beyond a qRT-PCR Study on Mesenchymal Stem Cell Differentiation Properties: How to Choose the Most Reliable Housekeeping Genes. J. Cell. Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, E.; Morris, C.; Chiesa, R.; Cigada, A.; Santin, M. In Vitro Assessment of the Osteointegrative Potential of a Novel Multiphase Anodic Spark Deposition Coating for Orthopaedic and Dental Implants. J. Biomed. Mater. Res. Part B Appl. Biomater. 2005, 73, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Toledano-Osorio, M.; Manzano-Moreno, F.J.; Toledano, M.; Osorio, R.; Medina-Castillo, A.L.; Costela-Ruiz, V.J.; Ruiz, C. Doxycycline-Doped Membranes Induced Osteogenic Gene Expression on Osteoblastic Cells. J. Dent. 2021, 109, 103676. [Google Scholar] [CrossRef] [PubMed]
- Rothamel, D.; Schwarz, F.; Fienitz, T.; Smeets, R.; Dreiseidler, T.; Ritter, L.; Happe, A.; Zöller, J. Biocompatibility and Biodegradation of a Native Porcine Pericardium Membrane: Results of in Vitro and in Vivo Examinations. Int. J. Oral. Maxillofac. Implant. 2012, 27, 146–154. [Google Scholar]
- Braga-Vilela, A.S.; Pimentel, E.R.; Marangoni, S.; Toyama, M.H.; de Campos Vidal, B. Extracellular Matrix of Porcine Pericardium: Biochemistry and Collagen Architecture. J. Membr. Biol. 2008, 221, 15–25. [Google Scholar] [CrossRef]
- Ng, H.W.; Zhang, Y.; Naffa, R.; Prabakar, S. Monitoring the Degradation of Collagen Hydrogels by Collagenase Clostridium Histolyticum. Gels 2020, 6, 46. [Google Scholar] [CrossRef]
- Toledano-Osorio, M.; Manzano-Moreno, F.J.; Ruiz, C.; Toledano, M.; Osorio, R. Testing Active Membranes for Bone Regeneration: A Review. J. Dent. 2021, 105, 103580. [Google Scholar] [CrossRef]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In Search of an Osteoblast Cell Model for in Vitro Research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. The RANKL/RANK/OPG Pathway. Curr. Osteoporos. Rep. 2007, 5, 98–104. [Google Scholar] [CrossRef]
- Schmidt, C.; Kaspar, D.; Sarkar, M.R.; Claes, L.E.; Ignatius, A.A. A Scanning Electron Microscopy Study of Human Osteoblast Morphology on Five Orthopedic Metals. J. Biomed. Mater. Res. 2002, 63, 252–261. [Google Scholar] [CrossRef]
- Zafar, S.; Coates, D.E.; Cullinan, M.P.; Drummond, B.K.; Milne, T.; Seymour, G.J. Effects of Zoledronic Acid and Geranylgeraniol on the Cellular Behaviour and Gene Expression of Primary Human Alveolar Osteoblasts. Clin. Oral. Investig. 2016, 20, 2023–2035. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-Y.; Ekeuku, S.O.; Trias, A. The Role of Geranylgeraniol in Managing Bisphosphonate-Related Osteonecrosis of the Jaw. Front. Pharmacol. 2022, 13, 878556. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Wei, A.; Whittaker, S.; Williams, L.A.; Tao, H.; Ma, D.D.F.; Diwan, A.D. The Role of BMP-7 in Chondrogenic and Osteogenic Differentiation of Human Bone Marrow Multipotent Mesenchymal Stromal Cells in Vitro. J. Cell. Biochem. 2010, 109, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Franceschi, R.T. Fibroblast Growth Factor 2 Induction of the Osteocalcin Gene Requires MAPK Activity and Phosphorylation of the Osteoblast Transcription Factor, Cbfa1/Runx2. J. Biol. Chem. 2002, 277, 36181–36187. [Google Scholar] [CrossRef]
- Portal-Núñez, S.; Lozano, D.; Esbrit, P. Role of Angiogenesis on Bone Formation. Histol. Histopathol. 2012, 27, 559–566. [Google Scholar]
- Street, J.; Lenehan, B. Vascular Endothelial Growth Factor Regulates Osteoblast Survival—Evidence for an Autocrine Feedback Mechanism. J. Orthop. Surg. Res. 2009, 4, 19. [Google Scholar] [CrossRef]
- Horner, A.; Bord, S.; Kelsall, A.W.; Coleman, N.; Compston, J.E. Tie2 Ligands Angiopoietin-1 and Angiopoietin-2 Are Coexpressed with Vascular Endothelial Cell Growth Factor in Growing Human Bone. Bone 2001, 28, 65–71. [Google Scholar] [CrossRef]
- Saadeh, P.B.; Mehrara, B.J.; Steinbrech, D.S.; Dudziak, M.E.; Greenwald, J.A.; Luchs, J.S.; Spector, J.A.; Ueno, H.; Gittes, G.K.; Longaker, M.T. Transforming Growth Factor-Beta1 Modulates the Expression of Vascular Endothelial Growth Factor by Osteoblasts. Am. J. Physiol. 1999, 277, C628–C637. [Google Scholar] [CrossRef]
- Evans, K.D.; Oberbauer, A.M. Alendronate Inhibits VEGF Expression in Growth Plate Chondrocytes by Acting on the Mevalonate Pathway. Open Orthop. J. 2009, 3, 83–88. [Google Scholar] [CrossRef]
- Jia, J.; Yao, W.; Amugongo, S.; Shahnazari, M.; Dai, W.; Lay, Y.-A.E.; Olvera, D.; Zimmermann, E.A.; Ritchie, R.O.; Li, C.-S.; et al. Prolonged Alendronate Treatment Prevents the Decline in Serum TGF-Β1 Levels and Reduces Cortical Bone Strength in Long-Term Estrogen Deficiency Rat Model. Bone 2013, 52, 424–432. [Google Scholar] [CrossRef]
- Sims, N.A.; Vrahnas, C. Regulation of Cortical and Trabecular Bone Mass by Communication between Osteoblasts, Osteocytes and Osteoclasts. Arch. Biochem. Biophys. 2014, 561C, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cao, X. BMP Signaling and Skeletogenesis. Ann. N. Y. Acad. Sci. 2006, 1068, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Zhang, L.; Jin, T.; Rutherford, R.B. Identification of Potential Modifiers of Runx2/Cbfa1 Activity in C2C12 Cells in Response to Bone Morphogenetic Protein-7. Cells Tissues Organs (Print) 2004, 176, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.-J.; Shirakawa, H.; Giriwono, P.E.; Ito, A.; Komai, M. A Novel Function of Geranylgeraniol in Regulating Testosterone Production. Biosci. Biotechnol. Biochem. 2018, 82, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, Y.; Nakahama, K.; Fujita, H.; Morita, I. Vitamin K2 and Geranylgeraniol, Its Side Chain Component, Inhibited Osteoclast Formation in a Different Manner. Biochem. Biophys. Res. Commun. 2004, 314, 24–30. [Google Scholar] [CrossRef] [PubMed]
- van Beek, E.; Pieterman, E.; Cohen, L.; Löwik, C.; Papapoulos, S. Farnesyl Pyrophosphate Synthase Is the Molecular Target of Nitrogen-Containing Bisphosphonates. Biochem. Biophys. Res. Commun. 1999, 264, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.E.; Rogers, M.J.; Halasy, J.M.; Luckman, S.P.; Hughes, D.E.; Masarachia, P.J.; Wesolowski, G.; Russell, R.G.; Rodan, G.A.; Reszka, A.A. Alendronate Mechanism of Action: Geranylgeraniol, an Intermediate in the Mevalonate Pathway, Prevents Inhibition of Osteoclast Formation, Bone Resorption, and Kinase Activation in Vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Reszka, A.A.; Halasy-Nagy, J.M.; Masarachia, P.J.; Rodan, G.A. Bisphosphonates Act Directly on the Osteoclast to Induce Caspase Cleavage of Mst1 Kinase during Apoptosis. A Link between Inhibition of the Mevalonate Pathway and Regulation of an Apoptosis-Promoting Kinase. J. Biol. Chem. 1999, 274, 34967–34973. [Google Scholar] [CrossRef]
- Jawad, M.J.; Ibrahim, S.; Kumar, M.; Burgert, C.; Li, W.-W.; Richardson, A. Identification of Foods That Affect the Anti-Cancer Activity of Pitavastatin in Cells. Oncol. Lett. 2022, 23, 73. [Google Scholar] [CrossRef]
- Retzepi, M.; Donos, N. Guided Bone Regeneration: Biological Principle and Therapeutic Applications. Clin. Oral. Implant. Res. 2010, 21, 567–576. [Google Scholar] [CrossRef]
- Xie, Y.; Chu, Z.; Jin, W. Beyond Separation: Membranes towards Medicine. J. Membr. Sci. Lett. 2022, 2, 100020. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense Primer (5′–3′) | Antisense Primer |
|---|---|---|
| ALP | CCAACGTGGCTAAGAATGTCATC | TGGGCATTGGTGTTGTACGTC |
| BMP-2 | TCGAAATTCCCCGTGACCAG | CCACTTCCACCACGAATCCA |
| BMP-7 | CTGGTCTTTGTCTGCAGTGG | GTACCCCTCAACAAGGCTTC |
| Col-1 | AGAACTGGTACATCAGCAAG | GAGTTTACAGGAAGCAGACA |
| OSX | TGCCTAGAAGCCCTGAGAAA | TTTAACTTGGGGCCTTGAGA |
| OPG | ATGCAACACAGCACAACATA | GTTGCCGTTTTATCCTCTCT |
| OSC | CCATGAGAGCCCTCACACTCC | GGTCAGCCAACTCGTCACAGTC |
| RANKL | ATACCCTGATGAAAGGAGGA | GGGGCTCAATCTATATCTCG |
| Runx-2 | TGGTTAATCTCCGCAGGTCAC | ACTGTGCTGAAGAGGCTGTTTG |
| TGFβ1 | TGAACCGGCCTTTCCTGCTTCTCATG | GCGGAAGTCAATGTACAGCTGCCGC |
| TGFβ-R1 | ACTGGCAGCTGTCATTGCTGGACCAG | CTGAGCCAGAACCTGACGTTGTCATATCA |
| TGFβ-R2 | GGCTCAACCACCAGGGCATCCAGAT | CTCCCCGAGAGCCTGTCCAGATGCT |
| TGFβ-R3 | ACCGTGATGGGCATTGCGTTTGCA | GTGCTCTGCGTGCTGCCGA TGCTGT |
| VEGF | CCTTGCTGCTCTACCTCCAC | CACACAGGATGGCTTGAAGA |
| UBC | TGGGATGCAAATCTTCGTGAAGACCCTGAC | ACCAAGTGCAGAGTGGACTCTTTCTGGATG |
| PPIA | CCATGGCAAATGCTGGACCCAACACAAATG | TCCTGAGCTACAGAAGGAATGATCTGGTGG |
| RPS13 | GGTGTTGCACAAGTACGTTTTGTGACAGGC | TCATATTTCCAATTGGGAGGGAGGACTCGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzano-Moreno, F.J.; de Luna-Bertos, E.; Toledano-Osorio, M.; Urbano-Arroyo, P.; Ruiz, C.; Toledano, M.; Osorio, R. Biomimetic Collagen Membranes as Drug Carriers of Geranylgeraniol to Counteract the Effect of Zoledronate. Biomimetics 2024, 9, 4. https://doi.org/10.3390/biomimetics9010004
Manzano-Moreno FJ, de Luna-Bertos E, Toledano-Osorio M, Urbano-Arroyo P, Ruiz C, Toledano M, Osorio R. Biomimetic Collagen Membranes as Drug Carriers of Geranylgeraniol to Counteract the Effect of Zoledronate. Biomimetics. 2024; 9(1):4. https://doi.org/10.3390/biomimetics9010004
Chicago/Turabian StyleManzano-Moreno, Francisco Javier, Elvira de Luna-Bertos, Manuel Toledano-Osorio, Paula Urbano-Arroyo, Concepción Ruiz, Manuel Toledano, and Raquel Osorio. 2024. "Biomimetic Collagen Membranes as Drug Carriers of Geranylgeraniol to Counteract the Effect of Zoledronate" Biomimetics 9, no. 1: 4. https://doi.org/10.3390/biomimetics9010004