The Interdependence between Schistosome Transmission and Protective Immunity


{kind=link}
Abstract
:1. Required Immunological Parameters
2. Delayed Concomitant Immunity
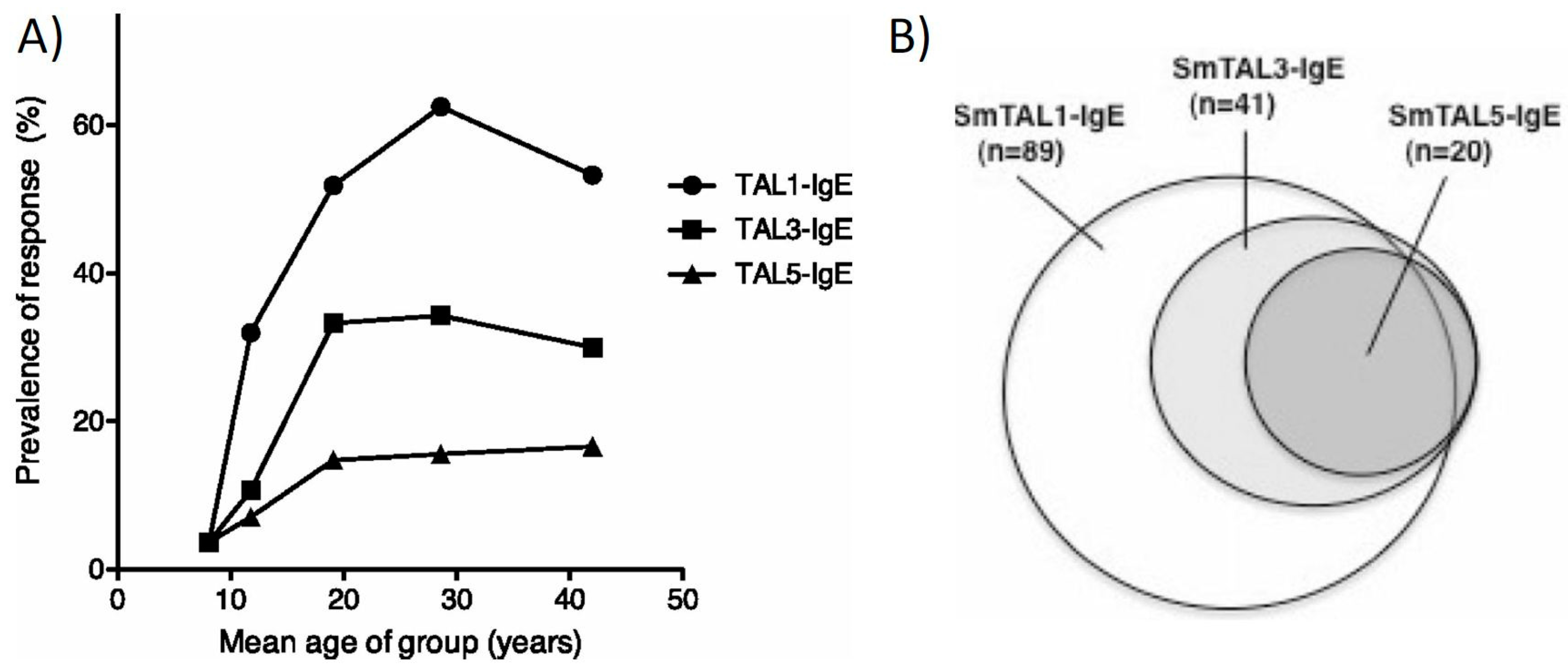
3. IgE Responses and Praziquantel Treatment
4. Rate of Decay of Protective Immunity
5. Variation in Estimates of Immune Parameters
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kabatereine, N.B.; Vennervald, B.J.; Ouma, J.H.; Kemijumbi, J.; Butterworth, A.E.; Dunne, D.W.; Fulford, A.J. Adult resistance to schistosomiasis mansoni: Age-dependence of reinfection remains constant in communities with diverse exposure patterns. Parasitology 1999, 118, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.; Hagan, P. Seeking the ghost of worms past. Nat. Med. 1999, 5, 1225–1227. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Taylor, P.; Matanhire, D.; Chandiwana, S.K. Acquired immunity and epidemiology of Schistosoma haematobium. Nature 1991, 351, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Fulford, A.; Butterworth, A.E.; Sturrock, R.F.; Ouma, J.H. On the use of age-intensity data to detect immunity to parasitic infections, with special reference to Schistosoma mansoni in Kenya. Parasitology 1992, 105 Pt 2, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.S.; Anderson, R.M.; Medley, G.F.; Bundy, D.A.P. Dynamic aspects of morbidity and acquired immunity in schistosomiasis control. Acta Trop. 1996, 62, 105–117. [Google Scholar] [CrossRef]
- Mutapi, F.; Ndhlovu, P.D.; Hagan, P.; Spicer, J.T.; Mduluza, T.; Turner, C.M.; Chandiwana, S.K.; Woolhouse, M.E. Chemotherapy accelerates the development of acquired immune responses to Schistosoma haematobium infection. J. Infect. Dis. 1998, 178, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R.M. Herd immunity to helminth infection and implications for parasite control. Nature 1985, 315, 493–496. [Google Scholar] [CrossRef]
- De Moira, A.P.; Fulford, A.J.C.; Kabatereine, N.B.; Kazibwe, F.; Ouma, J.H.; Dunne, D.W.; Booth, M. Microgeographical and tribal variations in water contact and Schistosoma mansoni exposure within a Ugandan fishing community. Trop. Med. Int. Health 2007, 12, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E. On the application of mathematical models of schistosome transmission dynamics. I. Natural transmission. Acta Trop. 1991, 49, 241–270. [Google Scholar] [CrossRef]
- Woolhouse, M.E.J. On the application of mathematical models of schistosome transmission dynamics. II. Control. Acta Trop. 1992, 50, 189–204. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Etard, J.F.; Dietz, K.; Ndhlovu, P.D.; Chandiwana, S.K. Heterogeneities in schistosome transmission dynamics and control. Parasitology 1998, 117, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, H.; Anderson, R.; Andreasen, V.; Bansal, S.; De, D.; Dye, C.; Eames, K.; Edmunds, J.; Frost, S.; Funk, S.; et al. Modeling infectious disease dynamics in the complex landscape of global health. Science 2015, 347, aaa4339. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Turner, H.C.; Farrell, S.H.; Yang, J.; Truscott, J.E. What is required in terms of mass drug administration to interrupt the transmission of schistosome parasites in regions of endemic infection? Parasit Vectors 2015, 8, 553. [Google Scholar] [CrossRef] [PubMed]
- Gurarie, D.; King, C.H.; Yoon, N.; Li, E. Refined stratified-worm-burden models that incorporate specific biological features of human and snail hosts provide better estimates of Schistosoma diagnosis, transmission, and control. Parasit Vectors 2016, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Truscott, J.E.; Gurarie, D.; Alsallaq, R.; Toor, J.; Yoon, N.; Farrell, S.H.; Turner, H.C.; Phillips, A.E.; Aurelio, H.O.; Ferro, J.; et al. A comparison of two mathematical models of the impact of mass drug administration on the transmission and control of schistosomiasis. Epidemics 2017, 18, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Restif, O.; Hayman, D.T.S.; Pulliam, J.R.C.; Plowright, R.K.; George, D.B.; Luis, A.D.; Cunningham, A.A.; Bowen, R.A.; Fooks, A.R.; O’Shea, T.J.; et al. Model-guided fieldwork: Practical guidelines for multidisciplinary research on wildlife ecological and epidemiological dynamics. Ecol. Lett. 2012, 15, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.J.; Victory, H.D. Modelling and simulation of a schistosomiasis infection with biological control. Acta Trop. 2003, 87, 251–267. [Google Scholar] [CrossRef]
- Hackett, L.W. Malaria in Europe: An Ecological Study; Oxford University Press: London, UK, 1937. [Google Scholar]
- Anderson, R.M.; Turner, H.C.; Farrell, S.H.; Truscott, J.E. Studies of the transmission dynamics, mathematical model development and the control of schistosome parasites by mass drug administration in human communities. Adv. Parasitol. 2016, 94, 199–246. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.M.; Mutapi, F.; Mduluza, T.; Midzi, N.; Savill, N.J.; Woolhouse, M.E.J. Predicted impact of mass drug administration on the development of protective immunity against Schistosoma haematobium. PLoS Negl. Trop. Dis. 2014, 8, e3059. [Google Scholar] [CrossRef] [PubMed]
- Gurarie, D.; Yoon, N.; Li, E.; Ndeffo-mbah, M.; Durham, D.; Phillips, A.E.; Aurelio, H.O.; Ferro, J.; Galvani, A.P.; King, C.H. Modelling control of Schistosoma haematobium infection: Predictions of the long-term impact of mass drug administration in Africa. Parasit. Vectors 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Coutinho, F.A.; Massad, E. Acquired immunity on a schistosomiasis transmission model—Fitting the data. J. Theor. Biol. 1997, 188, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Gurarie, D.; King, C.H. Heterogeneous model of schistosomiasis transmission and long-term control: The combined influence of spatial variation and age-dependent factors on optimal allocation of drug therapy. Parasitology 2005, 130, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Gurarie, D.; King, C.H. Population biology of Schistosoma mating, aggregation, and transmission breakpoints: More reliable model analysis for the end-game in communities at risk. PLoS ONE 2014, 9, e115875. [Google Scholar] [CrossRef] [PubMed]
- Mutapi, F.; Maizels, R.; Fenwick, A.; Woolhouse, M. Human schistosomiasis in the post-mass drug administration era. Lancet Infect. Dis. 2017, 17, e42–e48. [Google Scholar] [CrossRef]
- Woolhouse, M.E.J. A theoretical framework for the immunoepidemiology of helminth infection. Parasite Immunol. 1992, 14, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Jiz, M.; Friedman, J.F.; Leenstra, T.; Jarilla, B.; Pablo, A.; Langdon, G.; Pond-Tor, S.; Wu, H.-W.; Manalo, D.; Olveda, R.; et al. Immunoglobulin E (IgE) responses to paramyosin predict resistance to reinfection with Schistosoma japonicum and are attenuated by IgG4. Infect. Immun. 2009, 77, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Hagan, P.; Blumenthal, U.J.; Dunn, D.; Simpson, A.J.G.; Wilkins, H.A. Human IgE, IgG4 and resistance to reinfection with Schistosoma haematobium. Nature 1991, 349, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Dunne, D.W.; Butterworth, A.E.; Fulford, A.J.C.; Curtis Kariuki, H.; Langley, J.G.; Ouma, J.H.; Capron, A.; Pierce, R.J.; Sturrock, R.F. Immunity after treatment of human schistosomiasis: Association between IgE antibodies to adult worm antigens and resistance to reinfection. Eur. J. Immunol. 1992, 22, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, A.E.; Bensted-Smith, R.; Capron, A.; Capron, M.; Dalton, P.R.; Dunne, D.W.; Grzych, J.M.; Kariuki, H.C.; Khalife, J.; Koech, D. Immunity in human schistosomiasis mansoni: Prevention by blocking antibodies of the expression of immunity in young children. Parasitology 1987, 94, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Demeure, C.E.; Rihet, P.; Abel, L.; Ouattara, M.; Bourgois, A.; Dessein, A.J. Resistance to Schistosoma mansoni in humans: Influence of the IgE/IgG4 balance and IgG2 in immunity to reinfection after chemotherapy. J. Infect. Dis. 1993, 168, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, C.M.; Jones, F.M.; Pinot De Moira, A.; Protasio, A.V.; Khalife, J.; Dickinson, H.A.; Tukahebwa, E.M.; Dunne, D.W. Progressive cross-reactivity in IgE responses: An explanation for the slow development of human immunity to schistosomiasis? Infect. Immun. 2012, 80, 4264–4270. [Google Scholar] [CrossRef] [PubMed]
- Smithers, S.R.; Terry, R.J. Resistance to experimental infection with Schistosoma mansoni in rhesus monkeys induced by the transfer of adult worms. Trans. R. Soc. Trop. Med. Hyg. 1967, 61, 517–533. [Google Scholar] [CrossRef]
- Wilkins, H.A.; Goll, P.H.; Marshall, T.F.; Moore, P.J. Dynamics of Schistosoma haematobium infection in a Gambian community. III. Acquisition and loss of infection. Trans. R. Soc. Trop. Med. Hyg. 1984, 78, 227–232. [Google Scholar] [CrossRef]
- Fulford, A.J.; Butterworth, A.E.; Ouma, J.H.; Sturrock, R.F. A statistical approach to schistosome population dynamics and estimation of the life-span of Schistosoma mansoni in man. Parasitology 1995, 110 Pt 3, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.; Fulford, A.J.C.; Braun, G.; Ouma, J.H.; Kariuki, H.C.; Havercroft, J.C.; Gachuhi, K.; Sturrock, R.F.; Butterworth, A.E.; Dunne, D.W. Human immunoglobulin E responses to a recombinant 22.6-kilodalton antigen from Schistosoma mansoni adult worms are associated with low intensities of reinfection after treatment. Infect. Immun. 1996, 64, 4042–4046. [Google Scholar] [PubMed]
- Fitzsimmons, C.M.; Jones, F.M.; Stearn, A.; Chalmers, I.W.; Hoffmann, K.F.; Wawrzyniak, J.; Wilson, S.; Kabatereine, N.B.; Dunne, D.W.; Oliveira, S.C. The Schistosoma mansoni tegumental-allergen-like (TAL) protein family: Influence of developmental expression on human IgE responses. PLoS Negl. Trop. Dis. 2012, 6, e1593. [Google Scholar] [CrossRef] [PubMed]
- Pinot De Moira, A.; Jones, F.M.; Wilson, S.; Tukahebwa, E.; Fitzsimmons, C.M.; Mwatha, J.K.; Bethony, J.M.; Kabatereine, N.B.; Dunne, D.W. Effects of treatment on IgE responses against parasite allergen-like proteins and immunity to reinfection in childhood schistosome and hookworm coinfections. Infect. Immun. 2013, 81, 23–32. [Google Scholar] [CrossRef] [PubMed]
- De Moira, A.P.; Sousa-Figueiredo, J.C.; Jones, F.M.; Fitzsimmons, C.M.; Betson, M.; Kabatereine, N.B.; Stothard, J.R.; Dunne, D.W. Schistosoma mansoni infection in preschool-aged children: Development of immunoglobulin E and immunoglobulin G4 responses to parasite allergen-like proteins. J. Infect. Dis. 2012, 207, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.C.; Aynsley, S.A.; Turner, J.D.; Jenkins, G.R.; Van Rooijen, N.; Leeto, M.; Brombacher, F.; Mountford, A.P. Multiple helminth infection of the skin causes lymphocyte hypo-responsiveness mediated by Th2 conditioning of dermal myeloid cells. PLoS Pathog. 2011, 7, e1001323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganley-Leal, L.M.; Mwinzi, P.N.M.; Cetre-Sossah, C.B.; Andove, J.; Hightower, A.W.; Karanja, D.M.S.; Colley, D.G.; Secor, W.E. Higher percentages of circulating mast cell precursors correlate with susceptibility to reinfection with Schistosoma mansoni. Am. J. Trop. Med. Hyg. 2006, 75, 1053–1057. [Google Scholar] [PubMed]
- Soussi Gounni, A.; Lamkhioued, B.; Ochiai, K.; Tanaka, Y.; Delaporte, E.; Capron, A.; Kinet, J.-P.; Capron, M. High-affinity IgE receptor on eosinophils is involved in defence against parasites. Nature 1994, 367, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Hagan, P.; Wilkins, H.A.; Blumenthal, U.J.; Hayes, R.J.; Greenwood, B.M. Eosinophilia and resistance to Schistosoma haematobium in man. Parasite Immunol. 1985, 7, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Ganley-Leal, L.M.; Mwinzi, P.N.; Cetre-Sossah, C.B.; Andove, J.; Hightower, A.W.; Karanja, D.M.S.; Colley, D.G.; Secor, W.E. Correlation between eosinophils and protection against reinfection with Schistosoma mansoni and the effect of human immunodeficiency virus type 1 coinfection in humans. Infect. Immun. 2006, 74, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Karanja, D.M.; Hightower, A.W.; Colley, D.G.; Mwinzi, P.N.; Galil, K.; Andove, J.; Secor, W.E. Resistance to reinfection with Schistosoma mansoni in occupationally exposed adults and effect of HIV-1 co-infection on susceptibility to schistosomiasis: A longitudinal study. Lancet 2002, 360, 592–596. [Google Scholar] [CrossRef]
- Black, C.L.; Mwinzi, P.N.M.; Muok, E.M.O.; Abudho, B.; Fitzsimmons, C.M.; Dunne, D.W.; Karanja, D.M.S.; Secor, W.E.; Colley, D.G. Influence of exposure history on the immunology and development of resistance to human schistosomiasis mansoni. PLoS Negl. Trop. Dis. 2010, 4, e637. [Google Scholar] [CrossRef] [PubMed]
- Mwinzi, P.N.M.; Ganley-Leal, L.; Black, C.L.; Secor, W.E.; Karanja, D.M.S.; Colley, D.G. Circulating CD23+ B cell subset correlates with the development of resistance to Schistosoma mansoni reinfection in occupationally exposed adults who have undergone multiple treatments. J. Infect. Dis. 2009, 199, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Black, C.L.; Muok, E.M.O.; Mwinzi, P.N.M.; Carter, J.M.; Karanja, D.M.S.; Secor, W.E.; Colley, D.G. Increases in levels of Schistosome-specific immunoglobulin E and CD23+ B cells in a cohort of Kenyan children undergoing repeated treatment and reinfection with Schistosoma mansoni. J. Infect. Dis. 2010, 202, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Pinot De Moira, A.; Fulford, A.J.C.; Kabatereine, N.B.; Ouma, J.H.; Booth, M.; Dunne, D.W.; Quinnell, R.J. Analysis of complex patterns of human exposure and immunity to Schistosomiasis mansoni: The influence of age, sex, ethnicity and IgE. PLoS Negl. Trop. Dis. 2010, 4, e820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinot De Moira, A.; Fitzsimmons, C.M.; Jones, F.M.; Wilson, S.; Cahen, P.; Tukahebwa, E.; Mpairwe, H.; Mwatha, J.K.; Bethony, J.M.; Skov, P.S.; et al. Suppression of basophil histamine release and other IgE-dependent responses in childhood Schistosoma mansoni/hookworm coinfection. J. Infect. Dis. 2014, 210, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Colley, D.G.; Barsoum, I.S.; Dahawi, H.S.; Gamil, F.; Habib, M.; el Alamy, M.A. Immune responses and immunoregulation in relation to human schistosomiasis in Egypt. III. Immunity and longitudinal studies of in vitro responsiveness after treatment. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 952–957. [Google Scholar] [CrossRef]
- Medhat, A.; Shehata, M.; Bucci, K.; Mohamed, S.; Dief, A.D.; Badary, S.; Galal, H.; Nafeh, M.; King, C.L. Increased interleukin-4 and interleukin-5 production in response to Schistosoma haematobium adult worm antigens correlates with lack of reinfection after treatment. J. Infect. Dis. 1998, 178, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Jones, F.M.; Walter, K.; Fulford, A.J.; Kimani, G.; Mwatha, J.K.; Kamau, T.; Kariuki, H.C.; Kazibwe, F.; Tukahebwa, E.; et al. Increases in human T helper 2 cytokine responses to Schistosoma mansoni worm and worm-tegument antigens are induced by treatment with praziquantel. J. Infect. Dis. 2004, 190, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Van den Biggelaar, A.H.J.; Borrmann, S.; Kremsner, P.; Yazdanbakhsh, M. Immune responses induced by repeated treatment do not result in protective immunity to Schistosoma haematobium: Interleukin (IL)–5 and IL-10 responses. J. Infect. Dis. 2002, 186, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.; But1terworth, A.E.; Kimani, G.; Kamau, T.; Fulford, A.J.C.; Dunne, D.W.; Ouma, J.H.; Sturrock, R.F. Immunity after treatment of human schistosomiasis: Association between cellular responses and resistance to reinfection. Infect. Immun. 1993, 61, 4984–4993. [Google Scholar] [PubMed]
- Walter, K.; Fulford, A.J.C.; McBeath, R.; Joseph, S.; Jones, F.M.; Kariuki, H.C.; Mwatha, J.K.; Kimani, G.; Kabatereine, N.B.; Vennervald, B.J.; et al. Increased human IgE induced by killing Schistosoma mansoni in vivo is associated with pretreatment Th2 cytokine responsiveness to worm antigens. J. Immunol. 2006, 177, 5490–5498. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, C.M.; Joseph, S.; Jones, F.M.; Reimert, C.M.; Hoffmann, K.F.; Kazibwe, F.; Kimani, G.; Mwatha, J.K.; Ouma, J.H.; Tukahebwa, E.M.; et al. Chemotherapy for schistosomiasis in Ugandan fishermen: Treatment can cause a rapid increase in interleukin-5 levels in plasma but decreased levels of eosinophilia and worm-specific immunoglobulin E. Infect. Immun. 2004, 72, 4023–4030. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Jones, F.M.; Fofana, H.K.M.; Doucouré, A.; Landouré, A.; Kimani, G.; Mwatha, J.K.; Sacko, M.; Vennervald, B.J.; Dunne, D.W. Rapidly boosted plasma IL-5 induced by treatment of human schistosomiasis haematobium is dependent on antigen dose, IgE and eosinophils. PLoS Negl. Trop. Dis. 2013, 7, e2149. [Google Scholar] [CrossRef] [PubMed]
- French, M.D.; Churcher, T.S.; Gambhir, M.; Fenwick, A.; Webster, J.P.; Kabatereine, N.B.; Basáñez, M.G. Observed reductions in Schistosoma mansoni transmission from large-scale administration of praziquantel in Uganda: A mathematical modelling study. PLoS Negl. Trop. Dis. 2010, 4, e897. [Google Scholar] [CrossRef] [PubMed]
- French, M.D.; Churcher, T.S.; Webster, J.P.; Fleming, F.M.; Fenwick, A.; Kabatereine, N.B.; Sacko, M.; Garba, A.; Toure, S.; Nyandindi, U.; et al. Estimation of changes in the force of infection for intestinal and urogenital schistosomiasis in countries with schistosomiasis control initiative-assisted programmes. Parasit. Vectors 2015, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Agnew, A.; Fulford, A.J.; Mwanje, M.T.; Gachuhi, K.; Gutsmann, V.; Krijger, F.W.; Sturrock, R.F.; Vennervald, B.J.; Ouma, J.H.; Butterworth, A.E.; et al. Age-dependent reduction of schistosome fecundity in Schistosoma haematobium but not Schistosoma mansoni infections in humans. Am. J. Trop. Med. Hyg. 1996, 55, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Jones, F.M.; Van Dam, G.J.; Corstjens, P.L.; Riveau, G.; Fitzsimmons, C.M.; Sacko, M.; Vennervald, B.J.; Dunne, D.W. Human Schistosoma haematobium antifecundity immunity is dependent on transmission intensity and associated with immunoglobulin G1 to worm-derived antigens. J. Infect. Dis. 2014, 210, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- He, J.-S.; Narayanan, S.; Subramaniam, S.; Ho, W.Q.; Lafaille, J.J.; Curotto de Lafaille, M.A. Biology of IgE production: IgE cell differentiation and the memory of IgE responses. Curr. Top. Microbiol. Immunol. 2015, 388, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Jones, F.M.; Kenty, L.-C.; Mwatha, J.K.; Kimani, G.; Kariuki, H.C.; Dunne, D.W. Posttreatment changes in cytokines induced by Schistosoma mansoni egg and worm antigens: Dissociation of immunity- and morbidity-associated Type 2 responses. J. Infect. Dis. 2014, 209, 1792–1800. [Google Scholar] [CrossRef] [PubMed]
- Haniuda, K.; Fukao, S.; Kodama, T.; Hasegawa, H.; Kitamura, D. Autonomous membrane IgE signaling prevents IgE-memory formation. Nat. Immunol. 2016, 17, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Saiz, R.; Chu, D.K.; Mandur, T.S.; Walker, T.D.; Gordon, M.E.; Chaudhary, R.; Koenig, J.; Saliba, S.; Galipeau, H.J.; Utley, A.; et al. Lifelong memory responses perpetuate humoral TH 2 immunity and anaphylaxis in patients with food allergy. J. Allergy Clin. Immunol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.J.; Timbrell, V.; Xi, Y.; Upham, J.W.; Collins, A.M.; Davies, J.M. IgE+ B cells are scarce, but allergen-specific B cells with a memory phenotype circulate in patients with allergic rhinitis. Allergy 2015, 70, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.S.; Mutapi, F.; Woolhouse, M.E.; Isham, V.S. Stochastic simulation and the detection of immunity to schistosome infections. Parasitology 2000, 120 Pt 2, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.S.; Guyatt, H.L.; Bundy, D.A.; Booth, M.; Fulford, A.J.; Medley, G.F. The development of an age structured model for schistosomiasis transmission dynamics and control and its validation for Schistosoma mansoni. Epidemiol. Infect. 1995, 115, 325–344. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oettle, R.C.; Wilson, S. The Interdependence between Schistosome Transmission and Protective Immunity. Trop. Med. Infect. Dis. 2017, 2, 42. https://doi.org/10.3390/tropicalmed2030042
Oettle RC, Wilson S. The Interdependence between Schistosome Transmission and Protective Immunity. Tropical Medicine and Infectious Disease. 2017; 2(3):42. https://doi.org/10.3390/tropicalmed2030042
Chicago/Turabian StyleOettle, Rebecca C., and Shona Wilson. 2017. "The Interdependence between Schistosome Transmission and Protective Immunity" Tropical Medicine and Infectious Disease 2, no. 3: 42. https://doi.org/10.3390/tropicalmed2030042
APA StyleOettle, R. C., & Wilson, S. (2017). The Interdependence between Schistosome Transmission and Protective Immunity. Tropical Medicine and Infectious Disease, 2(3), 42. https://doi.org/10.3390/tropicalmed2030042