Phylogeography of Bulinus truncatus (Audouin, 1827) (Gastropoda: Planorbidae) in Selected African Countries

 ,
, 



Abstract
:1. Background
2. Materials and Methods
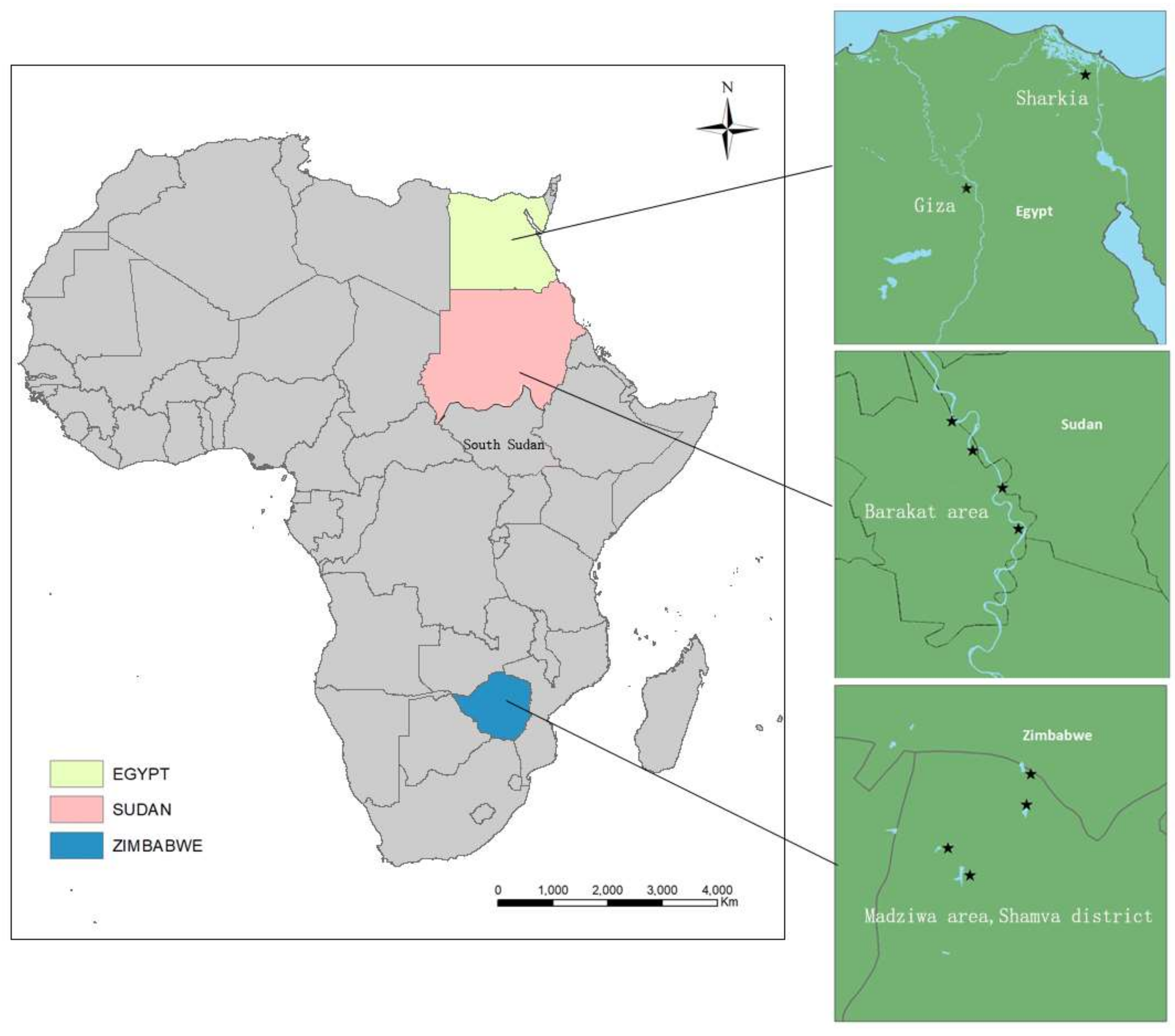
2.1. Sample Collection
2.2. Sample Preparation and DNA Extraction
2.3. Polymerase Chain Reaction (PCR) Amplification of COI and ITS 1 Fragments
2.4. Phylogenetic Analysis of Sequence Data
2.5. Determination of Haplotype and Nucleotide Diversity
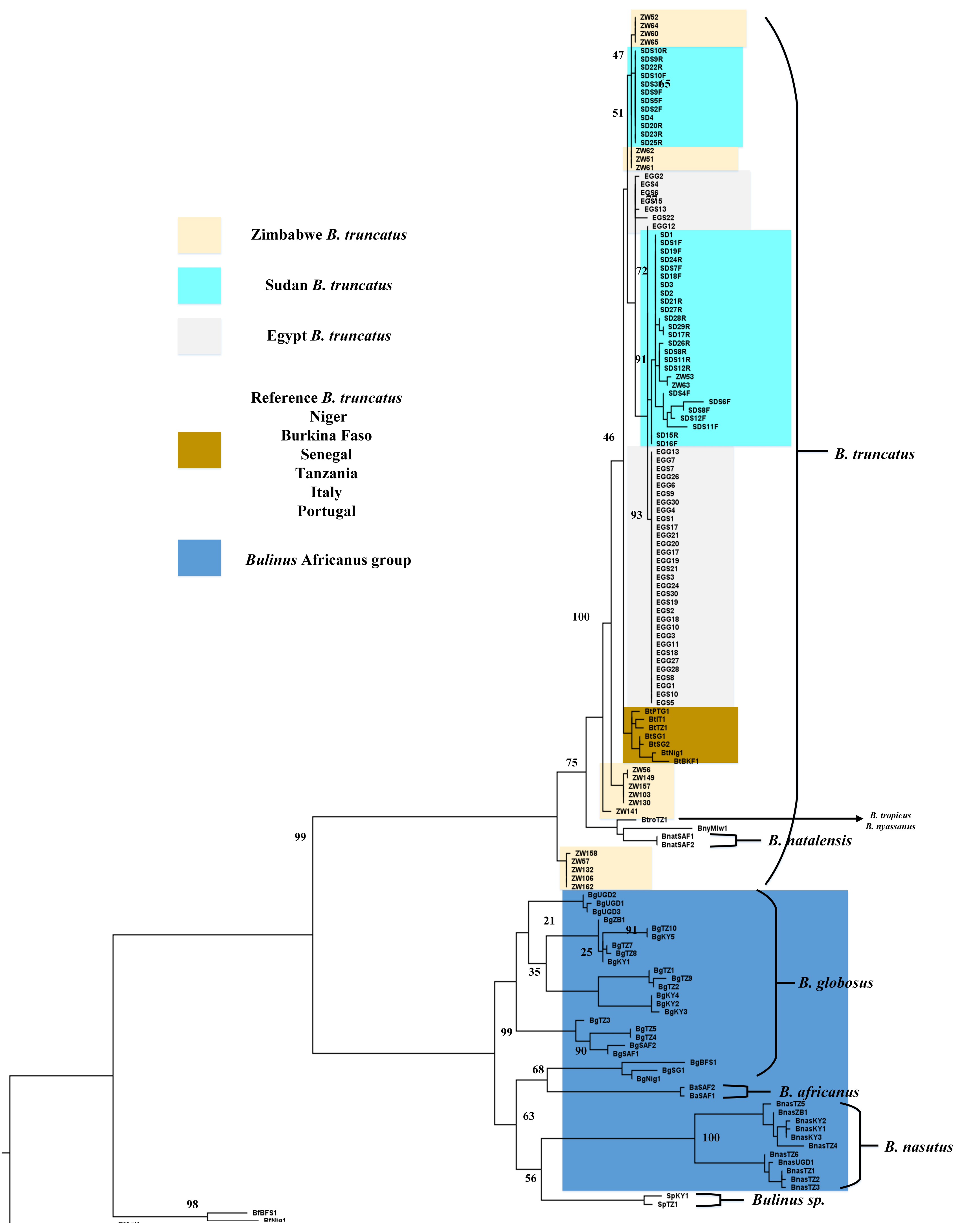
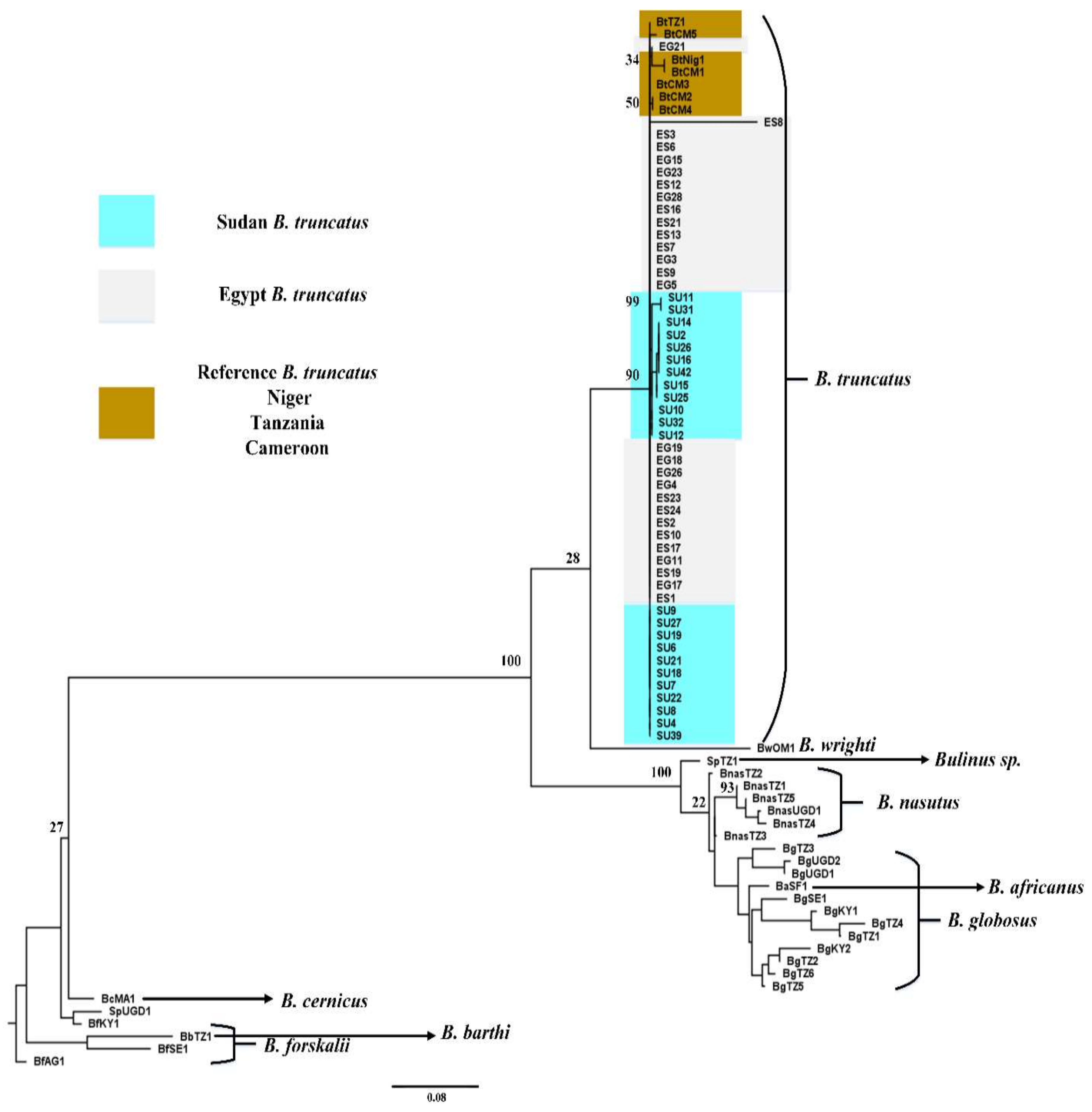
3. Results
3.1. Phylogeny
3.2. Haplotype and Nucleotide Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Country Profile: Preventive Chemotherapy and Transmission Control; World Health Organization: Geneva, Switzerland, 2010; Available online: http://www.who.int/neglected_diseases/preventive_chemotherapy/databank/CP (accessed on 10 February 2018).
- Hanington, P.C.; Forys, M.A.; Loker, E.S. A somatically diversified defense factor, FREP3, is a determinant of snail resistance to schistosome infection. PLoS Negl. Trop. Dis. 2012. [Google Scholar] [CrossRef] [PubMed]
- Gordy, M.A.; Kish, L.; Tarrabain, M.; Hanington, P.C. A comprehensive survey of larval digenean trematodes and their snail hosts in central Alberta, Canada. Parasitol. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rollinson, D.; Stothard, J.R.; Southgate, V.R. Interactions between intermediate snail hosts of the genus Bulinus and schistosomes of the Schistosoma haematobium group. Parasitology 2001, 123, S245–S260. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.S. Fresh Water Snails of Africa and Their Medical Importance, 2nd ed.; Taylor and Francis: London, UK, 1994; p. 609. [Google Scholar]
- Akinwale, O.P.; Kane, R.A.; Rollinson, D.; Stothard, J.R.; Ajayi, M.B.; Akande, D.O.; Ogungbemi, M.O.; Duker, C.; Gyang, P.V.; Adeleke, M.A. Molecular approaches to the identification of Bulinus species in south-west Nigeria and observations on natural snail infections with schistosomes. J. Helminthol. 2010, 85, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Jarne, P. Mating system, bottlenecks and genetic-polymorphism in hermaphroditic animals. Genet. Res. 1995, 65, 193–207. [Google Scholar] [CrossRef]
- Djuikwo-Teukenga, F.F.; Da Silva, A.; Njiokou, F.; Kamgang, B.; Same Ekobo, A.; Dreyfuss, G. Significant population genetic structure of the Cameroonian freshwater snail, Bulinus globosus, (Gastropoda: Planorbidae) revealed by nuclear microsatellite loci analysis. Acta Trop. 2014, 137, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.S. Freshwater Snails of Africa and Their Medical Importance; Taylor and Francis: London, UK, 1980; p. 487. [Google Scholar]
- Jarne, P.; Charlesworth, D. The evolution of the selfing rate in functionally hermaphrodite plants and animals. Annu. Rev. Ecol. Syst. 1993, 24, 441–466. [Google Scholar] [CrossRef]
- Zein-Eddine, R.; Djuikwo-Teukeng, F.F.; Dar, Y.; Dreyfuss, G.; Van den Broeck, F. Population genetics of the Schistosoma snail host Bulinus truncatus in Egypt. Acta Trop. 2017, 172, 36–43. [Google Scholar] [CrossRef]
- Kane, R.A.; Stothard, J.R.; Emery, A.M.; Rollinson, D. Molecular characterization of freshwater snails in the genus Bulinus: A role for bar codes? Parasites Vectors 2008, 1, 15. [Google Scholar] [CrossRef]
- Standley, C.J.; Goodacre, S.L.; Wade, C.M.; Stothard, J.R. The population genetic structure of Biomphalaria choanomphala in Lake Victoria, East Africa: Implications for schistosomiasis transmission. Parasites Vectors 2014, 7, 524. [Google Scholar] [CrossRef]
- Zein-Eddine, R.; Djuikwo-Teukeng, F.F.; Al-Jawhari, M.; Senghor, B.; Huyse, T.; Dreyfuss, G. Phylogeny of seven Bulinus species originating from endemic areas in three African countries, in relation to the human blood fluke Schistosoma haematobium. BMC Evol. Biol. 2014, 14, 271. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.H.; Sharaf El-Din, A.T.; Mohamed, A.M.; Habib, M.R. The relationship between genetic variability and the susceptibility of Biomphalaria alexandrina snails to Schistosoma mansoni infection. Mem. Inst. Oswaldo Cruz 2012, 107, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, Z.A.; Rawi, S.M.; Arafa, A.F. Identification of a genetic marker associated with the resistance to Schistosoma mansoni infection using random amplified polymorphic DNA analysis. Mem. Inst. Oswaldo Cruz 2006, 101, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Emery, A.M.; Loxton, N.J.; Stothard, J.R.; Jones, C.S.; Spinks, J.; Llewellyn-Hughes, J.; Noble, L.R.; Rollinson, D. Microsatellites in the freshwater snail Bulinus globosus (Gastropoda: Planorbidae) from Zanzibar. Mol. Ecol. Notes 2003, 3, 108–110. [Google Scholar] [CrossRef]
- Abe, E.M.; Guan, W.; Guo, Y.H.; Kassegne, K.; Qin, Z.Q.; Xu, J.; Chen, J.H.; Ekpo, U.F.; Li, S.Z.; Zhou, X.N. Differentiating snail intermediate hosts of Schistosoma spp. using molecular approaches: Fundamental to successful integrated control mechanism in Africa. Infect. Dis. Poverty 2018. [Google Scholar] [CrossRef] [PubMed]
- Djuikwo-Teukeng, F.F.; Njiokou, F.; Nkengazong, L.; De Meeus, T.; SameEkobo, A.; Dreyfuss, G. Strong genetic structure in Cameroonian populations of Bulinus truncatus (Gastropoda: Planorbidae), intermediate host of Schistosoma haematobium. Infect. Genet. Evol. 2001, 11, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Viard, F.; Bremond, P.; Labbo, R.; Justy, F.; Delay, B.; Jarne, P. Microsatellites and the genetics of highly selfing populations in the freshwater snail Bulinus truncatus. Genetics 1996, 142, 1237–1247. [Google Scholar]
- Viard, F.; Justy, F.; Jarne, P. Population dynamics inferred from temporal variation at microsatellite loci in the selfing snail Bulinus truncatus. Genetics 1997, 146, 973–982. [Google Scholar]
- Chlyeh, G.; Henry, P.Y.; Jarne, P. Spatial and temporal variation of life-history traits documented using capture-mark-recapture methods in the vector snail Bulinus truncatus. Parasitology 2003, 127, 243–251. [Google Scholar] [CrossRef]
- Jarne, P.; Viard, F.; Delay, B.; Cuny, G. Variable microsatellites in the highly selfing snail Bulinus truncatus (Basommatophora: Planorbidae). Mol. Ecol. 1994, 3, 527–528. [Google Scholar] [CrossRef]
- Brown, D.S.; Kristensen, T.K. A Fish Guide to Fresh Water Snails; Danish Bilharziasis Laboratory: DK-2920 Charlottenlund, Denmark, 1993. [Google Scholar]
- Emery, A.M.; Allan, F.E.; Rabone, M.E.; Rollinson, D. Schistosomiasis collection at NHM (SCAN). Parasit Vectors 2012, 5, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Avres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Evolution and phylogenetics software. FigTree version 1.4.3. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 8 December 2018).
- Polzin, T.; Daneshmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin ver.3.1: An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Goodall-Copestake, W.P.; Tarling, G.A.; Murphy, E.J. On the comparison of population-level estimates of haplotype and nucleotide diversity: A case study using the gene cox1 in animals. Heredity 2012, 109, 50–56. [Google Scholar] [CrossRef]
- Nalugwa, A.; Jørgensen, A.; Nyakaana, S.; Kristensen, TK. Molecular phylogeny of Bulinus (Gastropoda: Planorbidae) reveals the presence of three species complexes in the Albertine Rift freshwater bodies. Int. J. Genet. Mol. Biol. 2010, 2, 130–139. [Google Scholar]
- Brown, D.S.; Shaw, K.M. Freshwater snails of thee Bulinus truncatus/tropicus complex in Kenya: Tetraploid species. J. Mollus. Stud. 1989, 55, 509–532. [Google Scholar] [CrossRef]
- Allan, F.; Sousa-Figueiredo, J.C.; Emery, A.M.; Paulo, R.; Mirante, C.; Sebastião, A.; Brito, M.; Rollinson, D. Mapping freshwater snails in north-western Angola: Distribution, identity and molecular diversity of medically important taxa. Parasites Vectors 2017, 10, 460. [Google Scholar] [CrossRef] [PubMed]

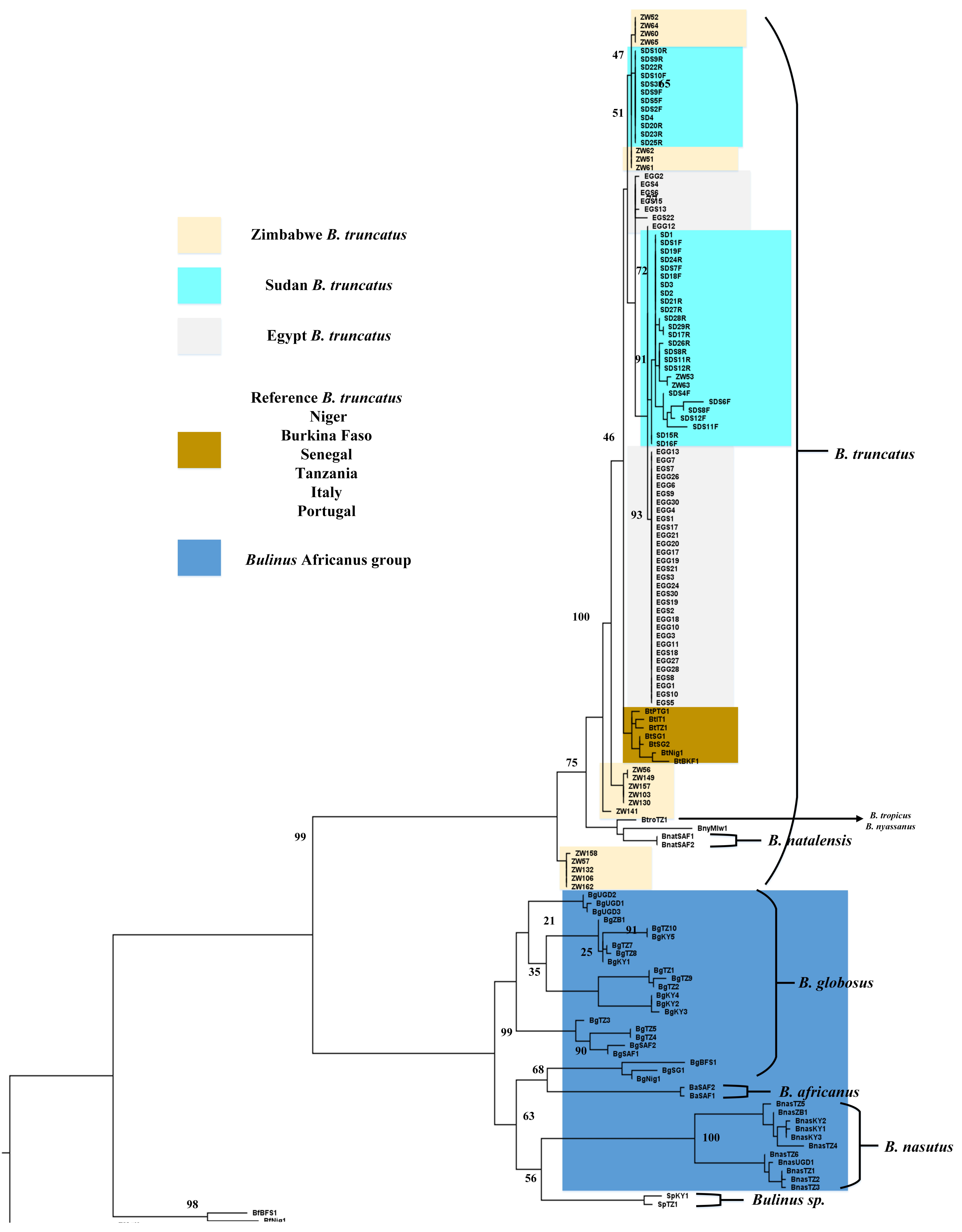


{kind=link}
{kind=link}
{kind=link}
| Country | Location | No. of Samples Collected | Time of Collection | Type of Water Body | Latitude | Longitude |
|---|---|---|---|---|---|---|
| Egypt | Giza (El-Nile river, Gezerite El-Warrak) | 25 | October, 2016 | River | 30.102 | 31.243 |
| Sharkia (El-Salam canal, El-Hesenia district) | 30 | November, 2016 | Canal | 31.258 | 32.270 | |
| Sudan | Barakat area, Wad Madani | 14 | July, 2016 | Canal | 14.33673 | 33.52736 |
| Barakat area, Wad Madani | 22 | August, 2016 | Canal | 14.31780 | 33.53167 | |
| Barakat area, Wad Madani | 5 | August, 2016 | Canal | 14.29210 | 33.55261 | |
| Barakat area, Wad Madani | 8 | August, 2016 | Canal | 14.25122 | 33.59070 | |
| Zimbabwe | Madziwa, Shamva District | 11 | March, 2016 | River | 16.93642 | 31.44603 |
| Madziwa | 6 | March, 2016 | River | 16.91498 | 31.42868 | |
| Madziwa | 10 | June, 2016 | River | 16.85695 | 31.49413 | |
| Madziwa | 3 | June, 2016 | River | 16.88070 | 31.49083 |
| N | H | Hd | π | S.D.π | ΘS | s.d.S | Tajima’s D | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|
| COI | Egypt | 38 | 6 | 0.334 | 0.00205 | 0.001445 | 0.002916 | 0.001299 | −0.85621 | 0.24 |
| Sudan | 36 | 11 | 0.768 | 0.009359 | 0.005067 | 0.009602 | 0.003341 | −0.08761 | 0.538 | |
| Zimbabwe | 20 | 6 | 0.784 | 0.014701 | 0.007859 | 0.011655 | 0.004442 | 1.01745 | 0.898 | |
| ITS 1 | Egypt | 28 | 3 | 0.14 | 0.005589 | 0.003362 | 0.020107 | 0.006909 | −2.69592 | 0 |
| Sudan | 23 | 2 | 0.443 | 0.00169 | 0.001367 | 0.001034 | 0.000769 | 1.41416 | 0.923 |
| Egypt | Sudan | |
|---|---|---|
| Sudan | 0.34860 * | |
| Zimbabwe | 0.59653 * | 0.36160 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abe, E.M.; Guo, Y.-H.; Shen, H.; Mutsaka-Makuvaza, M.J.; Habib, M.R.; Xue, J.-B.; Midzi, N.; Xu, J.; Li, S.-Z.; Zhou, X.-N. Phylogeography of Bulinus truncatus (Audouin, 1827) (Gastropoda: Planorbidae) in Selected African Countries. Trop. Med. Infect. Dis. 2018, 3, 127. https://doi.org/10.3390/tropicalmed3040127
Abe EM, Guo Y-H, Shen H, Mutsaka-Makuvaza MJ, Habib MR, Xue J-B, Midzi N, Xu J, Li S-Z, Zhou X-N. Phylogeography of Bulinus truncatus (Audouin, 1827) (Gastropoda: Planorbidae) in Selected African Countries. Tropical Medicine and Infectious Disease. 2018; 3(4):127. https://doi.org/10.3390/tropicalmed3040127
Chicago/Turabian StyleAbe, Eniola M., Yun-Hai Guo, Haimo Shen, Masceline J. Mutsaka-Makuvaza, Mohamed R. Habib, Jing-Bo Xue, Nicholas Midzi, Jing Xu, Shi-Zhu Li, and Xiao-Nong Zhou. 2018. "Phylogeography of Bulinus truncatus (Audouin, 1827) (Gastropoda: Planorbidae) in Selected African Countries" Tropical Medicine and Infectious Disease 3, no. 4: 127. https://doi.org/10.3390/tropicalmed3040127
APA StyleAbe, E. M., Guo, Y.-H., Shen, H., Mutsaka-Makuvaza, M. J., Habib, M. R., Xue, J.-B., Midzi, N., Xu, J., Li, S.-Z., & Zhou, X.-N. (2018). Phylogeography of Bulinus truncatus (Audouin, 1827) (Gastropoda: Planorbidae) in Selected African Countries. Tropical Medicine and Infectious Disease, 3(4), 127. https://doi.org/10.3390/tropicalmed3040127