Molecular Evidence of a Broad Range of Pathogenic Bacteria in Ctenocephalides spp.: Should We Re-Examine the Role of Fleas in the Transmission of Pathogens?

 ,
, 
 and
and 
Abstract
:1. Introduction
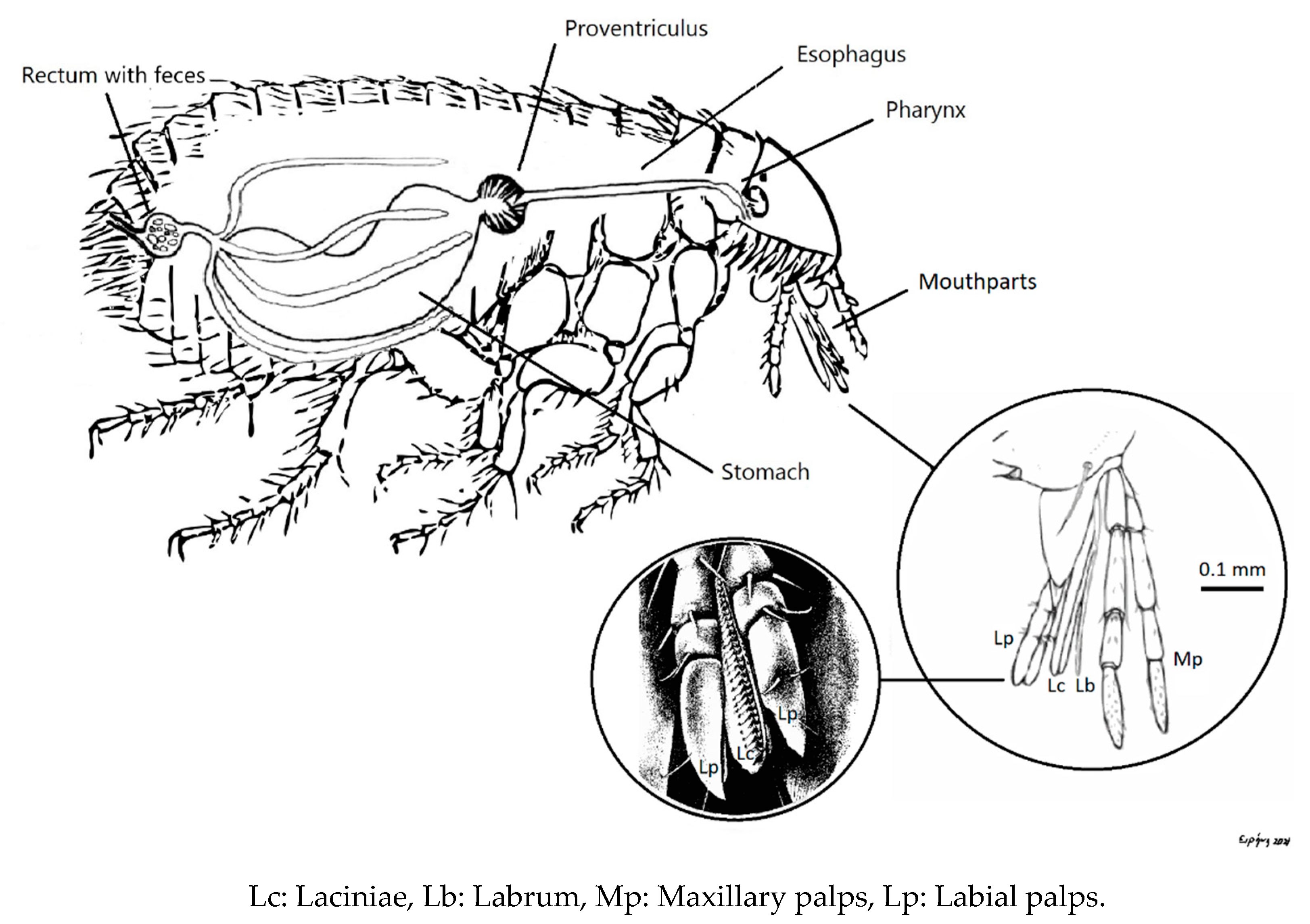
2. Materials and Methods
3. Results
3.1. Flea Samples
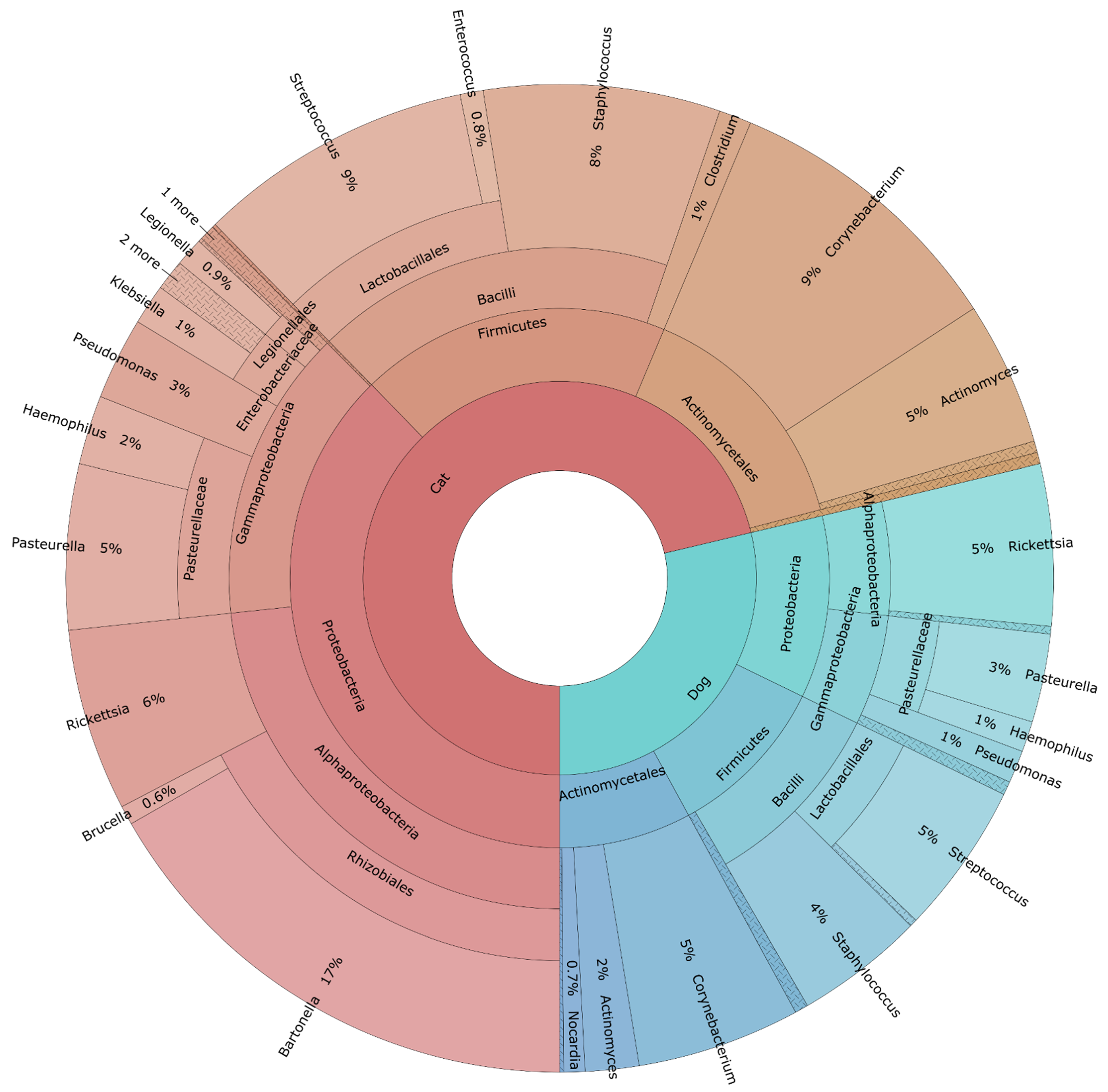
3.2. Flea Internal Microbiome
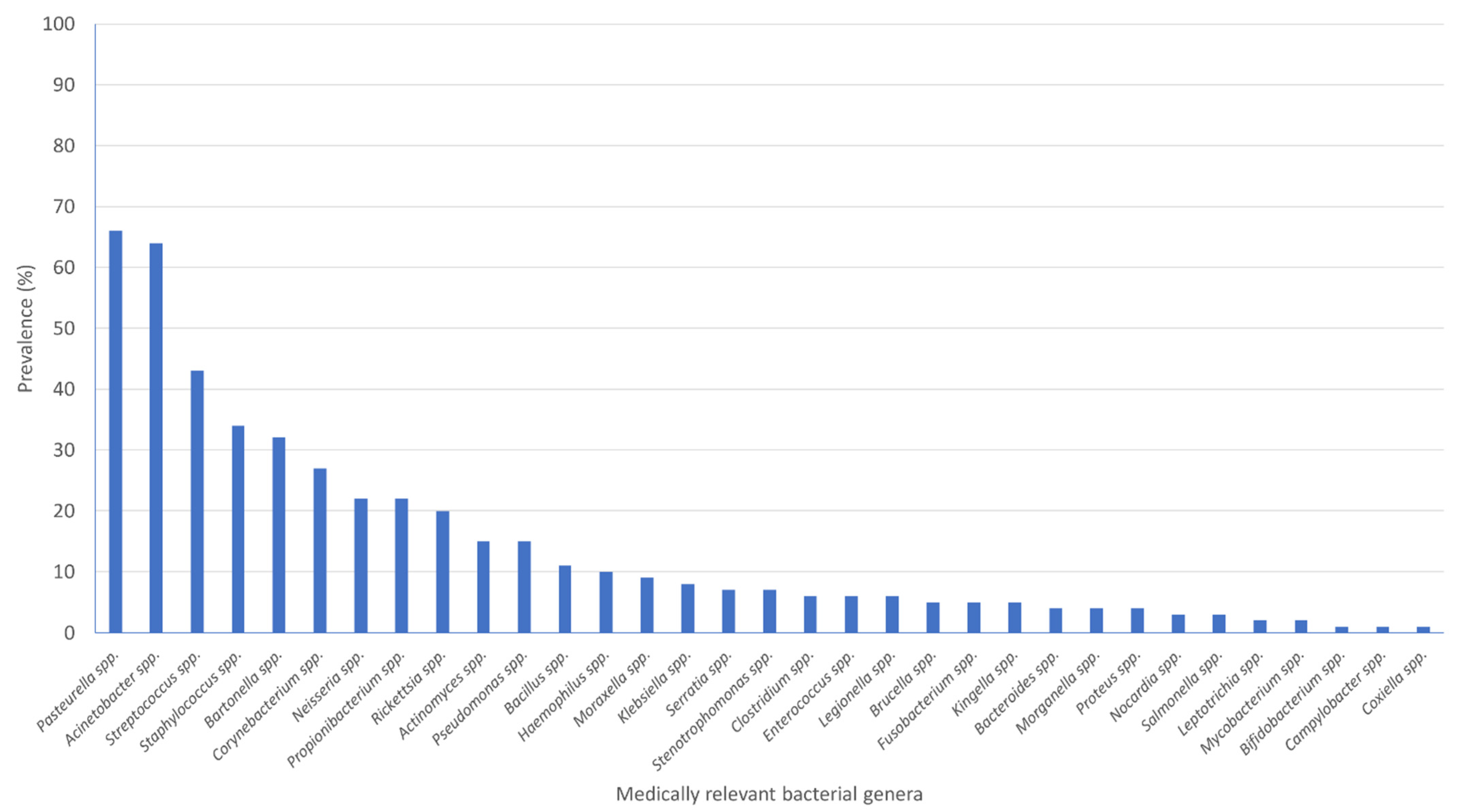
3.3. Medically Relevant Bacterial Genera
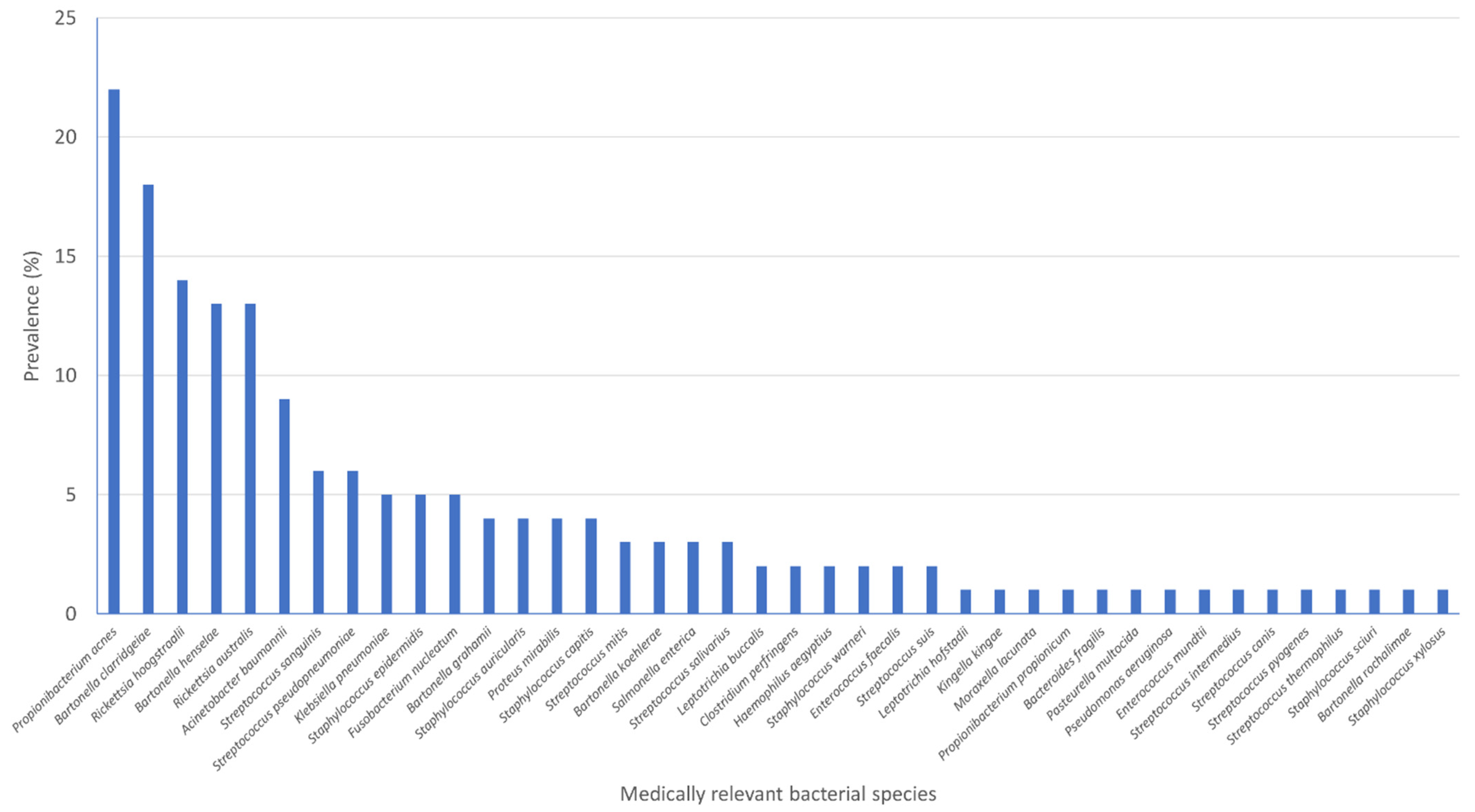
3.4. Medically Relevant Bacterial Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clark, N.J.; Seddon, J.M.; Šlapeta, J.; Wells, K. Parasite spread at the domestic animal—Wildlife interface: Anthropogenic habitat use, phylogeny and body mass drive risk of cat and dog flea (Ctenocephalides spp.) infestation in wild mammals. Parasit. Vectors 2018, 11, 8. [Google Scholar] [CrossRef]
- Bitam, I.; Dittmar, K.; Parola, P.; Whiting, M.F.; Raoult, D. Fleas and flea-borne diseases. Int. J. Infect. Dis. 2010, 14, e667–e676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenn, H.W.; Aspöck, H. Form, function and evolution of the mouthparts of blood-feeding Arthropoda. Arthropod Struct. Dev. 2012, 41, 101–118. [Google Scholar] [CrossRef]
- Legendre, K.P.; Macaluso, K.R. Rickettsia felis: A review of transmission mechanisms of an emerging pathogen. Trop. Med. Infect. Dis. 2017, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- Gilles, J.; Just, F.T.; Silaghi, C.; Pradel, I.; Lengauer, H.; Hellmann, K.; Pfister, K. Rickettsia felis in fleas, France. Emerg. Infect. Dis. 2008, 14, 684–686. [Google Scholar] [CrossRef]
- Zouari, S.; Khrouf, F.; M’ghirbi, Y.; Bouattour, A. First molecular detection and characterization of zoonotic Bartonella species in fleas infesting domestic animals in Tunisia. Parasit. Vectors 2017, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Cheslock, M.A.; Embers, M.E. Human bartonellosis: An underappreciated public health problem? Trop. Med. Infect. Dis. 2019, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, E.J.R.; Billeter, S.A.; Jett, L.A.; Meinersmann, R.J.; Barr, M.C.; Diniz, P.V.P.P.; Oakley, B.B. Assessing cat flea microbiomes in Northern and Southern California by 16S rRNA next-generation sequencing. Vector Borne Zoonotic Dis. 2018, 18, 491–499. [Google Scholar] [CrossRef]
- Takhampunya, R.; Korkusol, A.; Pongpichit, C.; Yodin, K.; Rungrojn, A.; Chanarat, N.; Promsathaporn, S.; Monkanna, T.; Thaloengsok, S.; Tippayachai, B.; et al. Metagenomic approach to characterizing disease epidemiology in a disease-endemic environment in Northern Thailand. Front. Microbiol. 2019, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, A.L.; Hii, S.F.; Chong, R.; Webb, C.E.; Traub, R.; Brown, G.; Šlapeta, J. Evaluation of the bacterial microbiome of two flea species using different DNA-isolation techniques provides insights into flea host ecology. FEMS Microbiol. Ecol. 2015, 91, fiv134. [Google Scholar] [CrossRef] [Green Version]
- Pratt, H.D.; Stojanovich, C.H.J. Pictorial Keys Arthropods, Reptiles, Birds and Mammals of Public Health Significance; Public Health Service, Communicable Disease Center: Atlanta, GA, USA, 1966; pp. 171–174. [Google Scholar]
- Andrews, E.S. Analyzing arthropods for the presence of bacteria. Curr. Protoc. Microbiol. 2013, 28, 1E.6.1–1E.6.14. [Google Scholar] [CrossRef] [Green Version]
- Regional typologies. In Eurostat Methodological Manual on Territorial Typologies, 2018th ed.; Publications Office of the European Union: Luxemburg, 2019; p. 77. Available online: https://ec.europa.eu/eurostat/web/products-manuals-and-guidelines/-/KS-GQ-18-008 (accessed on 7 January 2021). [CrossRef]
- Mediannikov, O.; Aubadie-Ladrix, M.; Raoult, D. Candidatus ‘Rickettsia senegalensis’ in cat fleas in Senegal. New Microbes New Infect. 2014, 3, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Utsuki, T.; Husseneder, C.; Chouljenko, V.; Azad, A.F.; Macaluso, K.R. Genomic diversification in strains of Rickettsia felis Isolated from different arthropods. Genome Biol. Evol. 2014, 7, 35–56. [Google Scholar] [CrossRef]
- Silaghi, C.; Knaus, M.; Rapti, D.; Shukullari, E.; Pfister, K.; Rehbein, S. Rickettsia felis and Bartonella spp. in fleas from cats in Albania. Vector Borne Zoonotic Dis. 2012, 12, 76–77. [Google Scholar] [CrossRef]
- Boudebouch, N.; Sarih, M.; Beaucournu, J.C.; Amarouch, H.; Hassar, M.; Raoult, D.; Parola, P. Bartonella clarridgeiae, B. Henselae and Rickettsia felis in fleas from Morocco. Ann. Trop. Med. Parasitol. 2011, 105, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H.; Wall, R. Pathogens in fleas collected from cats and dogs: Distribution and prevalence in the UK. Parasit. Vectors 2019, 12, 71. [Google Scholar] [CrossRef]
- Rossi, E.; Paroni, M.; Landini, P. Biofilm and motility in response to environmental and host-related signals in Gram negative opportunistic pathogens. J. Appl. Microbiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, B.J.; Erickson, D.L. Yersinia pestis biofilm in the flea vector and its role in the transmission of plague. Curr. Top. Microbiol. Immunol. 2008, 322, 229–248. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.D.; Banajee, K.H.; Foil, L.D.; Macaluso, K.R. Transmission mechanisms of an emerging insect-borne rickettsial pathogen. Parasit. Vectors 2016, 9, 237. [Google Scholar] [CrossRef] [Green Version]
- Peniche Lara, G.; Dzul-Rosado, K.R.; Velázquez, J.E.Z.; Zavala-Castro, J. Murine Typhus: Clinical and epidemiological aspects. Colomb Med. 2012, 43, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Viscoli, C. Bloodstream infections: The peak of the iceberg. Virulence 2016, 7, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Courjon, J.; Demonchy, E.; Degand, N.; Risso, K.; Ruimy, R.; Roger, P.M. Patients with community-acquired bacteremia of unknown origin: Clinical characteristics and usefulness of microbiological results for therapeutic issues: A single-center cohort study. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 40. [Google Scholar] [CrossRef] [Green Version]
- Lassmann, B.; Gustafson, D.R.; Wood, C.M.; Rosenblatt, J.E. Reemergence of anaerobic bacteremia. Clin. Infect. Dis. 2007, 44, 895–900. [Google Scholar] [CrossRef]
- Trell, K.; Nilson, B.; Petersson, A.C.; Rasmussen, M. Clinical and microbiological features of bacteremia with Streptococcus equi. Diagn. Microbiol. Infect. Dis. 2017, 87, 196–198. [Google Scholar] [CrossRef]
- Lin, J.N.; Lai, C.H.; Yang, C.H.; Huang, Y.H. Elizabethkingia infections in humans: From genomics to clinics. Microorganisms 2019, 7, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García López, E.; Martín-Galiano, A.J. The versatility of opportunistic infections caused by Gemella Isolates is supported by the carriage of virulence factors from multiple origins. Front. Microbiol. 2020, 11, 524. [Google Scholar] [CrossRef]
- Watanabe, T.; Hara, Y.; Yoshimi, Y.; Fujita, Y.; Yokoe, M.; Noguchi, Y. Clinical characteristics of bloodstream infection by Parvimonas micra: Retrospective case series and literature review. BMC Infect. Dis. 2020, 20, 578. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.B.; Borchert, J.N.; Black, W.C.; Atiku, L.A.; Mpanga, J.T.; Boegler, K.A.; Moore, S.M.; Gage, K.L.; Eisen, R.J. Blood meal identification in off-host cat fleas (Ctenocephalides felis) from a plague-endemic region of Uganda. Am. J. Trop. Med. Hyg. 2013, 88, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef] [PubMed]
- Allos, B.M.; Moore, M.R.; Griffin, P.M.; Tauxe, R.V. Surveillance for sporadic foodborne disease in the 21st century: The FoodNet perspective. Clin. Infect. Dis. 2004, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougas, G.; Tsakris, A.; Beleri, S.; Patsoula, E.; Billinis, C.; Papaparaskevas, J. Evidence of brucella melitensis DNA in the microbiome of ctenocephalides felis from pet cats in Greece. Vector Borne Zoonotic Dis. 2020, 20, 390–392. [Google Scholar] [CrossRef]
- Cadiergues, M.C.; Santamarta, D.; Mallet, X.; Franc, M. First blood meal of Ctenocephalides canis (Siphonaptera: Pulicidae) on dogs: Time to initiation of feeding and duration. J. Parasitol. 2001, 87, 214–215. [Google Scholar] [CrossRef]
- Wang, C.; Mount, J.; Butler, J.; Gao, D.; Jung, E.; Blagburn, B.L.; Kaltenboeck, B. Real-Time PCR of the mammalian hydroxymethylbilane synthase (HMBS) gene for analysis of flea (Ctenocephalides felis) feeding patterns on dogs. Parasit. Vectors 2012, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Winand, R.; Bogaerts, B.; Hoffman, S.; Lefevre, L.; Delvoye, M.; Braekel, J.V.; Fu, Q.; Roosens, N.H.; Keersmaecker, S.C.; Vanneste, K. Targeting the 16S rRNA gene for bacterial identification in complex mixed samples: Comparative evaluation of second (illumina) and third (oxford nanopore technologies) generation sequencing technologies. Int. J. Mol. Sci. 2019, 21, 298. [Google Scholar] [CrossRef] [Green Version]
- Werno, A.M.; Christner, M.; Anderson, T.P.; Murdoch, D.R. Differentiation of Streptococcus pneumoniae from nonpneumococcal streptococci of the Streptococcus mitis group by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2012, 50, 2863–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | Host Age-Group | Host Gender | Host Status | Flea Species | Flea-Pools |
|---|---|---|---|---|---|
| Cat | 0−1 years | Female | Stray | C. felis | 24 |
| Owned | C. felis | 2 | |||
| Male | Stray | C. felis | 15 | ||
| Unknown | C. felis | 1 | |||
| 1−5 years | Female | Stray | C. felis | 9 | |
| Owned | C. felis | 1 | |||
| Male | Stray | C. felis | 5 | ||
| Owned | C. felis | 1 | |||
| >5 years | Female | Stray | C. felis | 2 | |
| Owned | C. felis | 4 | |||
| Male | Stray | C. felis | 1 | ||
| Unknown | Female | Stray | C. felis | 2 | |
| Dog | 0−1 years | Female | Stray | C. felis | 2 |
| Owned | C. felis | 1 | |||
| P. irritans | 1 | ||||
| Male | Stray | C. felis | 3 | ||
| C. canis | 1 | ||||
| Owned | C. felis | 4 | |||
| C. canis | 1 | ||||
| Unknown | Stray | P. irritans | 1 | ||
| 1−5 years | Female | Stray | C. felis | 3 | |
| Owned | C. felis | 7 | |||
| Male | Stray | C. felis | 2 | ||
| Owned | C. felis | 2 | |||
| >5 years | Female | Owned | C. felis | 4 | |
| Male | Owned | C. felis | 1 |
| MR Bacterial Genera | MR Bacterial Species | No of Flea-Pools | MR Bacterial Genera | MR Bacterial Species | No of Flea-Pools |
|---|---|---|---|---|---|
| Acinetobacter | baumannii | 9 | Mycobacterium | Species unidentified | 2 |
| Species unidentified | 58 | Neisseria | Species unidentified | 15 | |
| Actinomyces | Species unidentified | 12 | Nocardia | Species unidentified | 3 |
| Bacillus | Species unidentified | 10 | Pasteurella | multocida | 1 |
| Bacteroides | fragilis | 1 | Species unidentified | 65 | |
| Species unidentified | 1 | Propionibacterium | acnes | 22 | |
| Bartonella | clarridgeiae | 18 | propionicum | 1 | |
| grahamii | 4 | Species unidentified | 0 | ||
| henselae | 13 | Proteus | mirabilis | 4 | |
| koehlerae | 3 | Species unidentified | 0 | ||
| rochalimae | 1 | Pseudomonas | aeruginosa | 1 | |
| Species unidentified | 35 | Species unidentified | 15 | ||
| Bifidobacterium | Species unidentified | 1 | Rickettsia | australis | 13 |
| Brucella | Species unidentified | 5 | hoogstraalii | 14 | |
| Campylobacter | Species unidentified | 1 | Species unidentified | 20 | |
| Clostridium | perfringens | 2 | Salmonella | enterica | 3 |
| Species unidentified | 6 | Species unidentified | 0 | ||
| Corynebacterium | Species unidentified | 24 | Serratia | Species unidentified | 7 |
| Coxiella | Species unidentified | 1 | Staphylococcus | auricularis | 4 |
| Enterococcus | faecalis | 2 | capitis | 4 | |
| mundtii | 1 | epidermidis | 5 | ||
| Species unidentified | 4 | sciuri | 1 | ||
| Fusobacterium | nucleatum | 5 | warneri | 2 | |
| Species unidentified | 1 | xylosus | 1 | ||
| Haemophilus | aegyptius | 2 | Species unidentified | 31 | |
| Species unidentified | 6 | Stenotrophomonas | Species unidentified | 3 | |
| Kingella | kingae | 1 | Streptococcus | canis | 1 |
| Species unidentified | 1 | intermedius | 1 | ||
| Klebsiella | pneumoniae | 5 | mitis | 3 | |
| Species unidentified | 3 | pseudopneumoniae | 6 | ||
| Legionella | Species unidentified | 6 | pyogenes | 1 | |
| Leptotrichia | buccalis | 2 | salivarius | 3 | |
| hofstadii | 1 | sanguinis | 6 | ||
| Species unidentified | 0 | suis | 2 | ||
| Moraxella | lacunata | 1 | thermophilus | 1 | |
| Species unidentified | 5 | Species unidentified | 43 | ||
| Morganella | Species unidentified | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dougas, G.; Tsakris, A.; Beleri, S.; Patsoula, E.; Linou, M.; Billinis, C.; Papaparaskevas, J. Molecular Evidence of a Broad Range of Pathogenic Bacteria in Ctenocephalides spp.: Should We Re-Examine the Role of Fleas in the Transmission of Pathogens? Trop. Med. Infect. Dis. 2021, 6, 37. https://doi.org/10.3390/tropicalmed6010037
Dougas G, Tsakris A, Beleri S, Patsoula E, Linou M, Billinis C, Papaparaskevas J. Molecular Evidence of a Broad Range of Pathogenic Bacteria in Ctenocephalides spp.: Should We Re-Examine the Role of Fleas in the Transmission of Pathogens? Tropical Medicine and Infectious Disease. 2021; 6(1):37. https://doi.org/10.3390/tropicalmed6010037
Chicago/Turabian StyleDougas, Georgios, Athanassios Tsakris, Stavroula Beleri, Eleni Patsoula, Maria Linou, Charalambos Billinis, and Joseph Papaparaskevas. 2021. "Molecular Evidence of a Broad Range of Pathogenic Bacteria in Ctenocephalides spp.: Should We Re-Examine the Role of Fleas in the Transmission of Pathogens?" Tropical Medicine and Infectious Disease 6, no. 1: 37. https://doi.org/10.3390/tropicalmed6010037
APA StyleDougas, G., Tsakris, A., Beleri, S., Patsoula, E., Linou, M., Billinis, C., & Papaparaskevas, J. (2021). Molecular Evidence of a Broad Range of Pathogenic Bacteria in Ctenocephalides spp.: Should We Re-Examine the Role of Fleas in the Transmission of Pathogens? Tropical Medicine and Infectious Disease, 6(1), 37. https://doi.org/10.3390/tropicalmed6010037