
Predominance of Multidrug Resistant Escherichia coli of Environmental Phylotype in Different Environments of Dhaka, Bangladesh

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Identification of Escherichia coli
2.3. Antibiotic Susceptibility Pattern Determination
2.4. Determination of Phylogenetic Group
2.5. Statistical Analysis
3. Results
3.1. Identification of the E. coli Isolates
3.2. Antimicrobial Resistance Evaluation of E. coli
3.3. Phylogroup Diversity of the E. coli Isolates
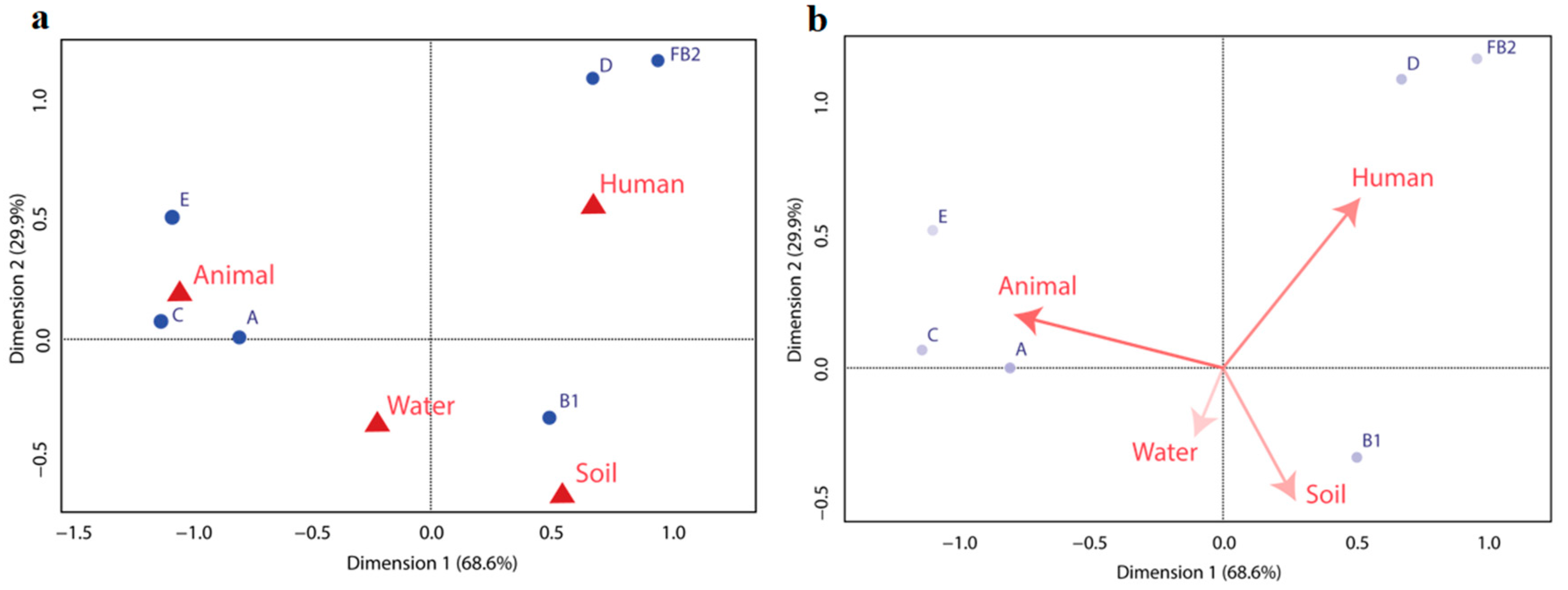
3.4. Correspondence Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Wang, W.; Xu, W.; Wang, X.; Zhao, S. Mercury Removals by Existing Pollutants Control Devices of Four Coal-Fired Power Plants in China. J. Environ. Sci. 2011, 23, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- Montealegre, M.C.; Roy, S.; Böni, F.; Hossain, M.I.; Navab-Daneshmand, T.; Caduff, L.; Faruque, A.S.G.; Islam, M.A.; Julian, T.R. Risk Factors for Detection, Survival, and Growth of Antibiotic-Resistant and Pathogenic Escherichia coli in Household Soils in Rural Bangladesh. Appl. Environ. Microbiol. 2018, 84, e01978-18. [Google Scholar] [CrossRef] [PubMed]
- Byappanahalli, M.N.; Fujioka, R.S. Evidence That Tropical Soil Environment Can Support the Growth of Escherichia coli. Water Sci. Technol. 1998, 38, 171–174. [Google Scholar] [CrossRef]
- Fujioka, R.S. Monitoring Coastal Marine Waters for Spore-Forming Bacteria of Faecal and Soil Origin to Determine Point from Non-Point Source Pollution. Water Sci. Technol. 2001, 44, 181. [Google Scholar] [CrossRef]
- Solo-Gabriele, H.M.; Wolfert, M.A.; Desmarais, T.R.; Palmer, C.J. Sources of Escherichia coli in a Coastal Subtropical Environment. Appl. Environ. Microbiol. 2000, 66, 230–237. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Whitman, R.L.; Shively, D.A.; Sadowsky, M.J.; Ishii, S. Population Structure, Persistence, and Seasonality of Autochthonous Escherichia coli in Temperate, Coastal Forest Soil from a Great Lakes Watershed. Environ. Microbiol. 2006, 8, 504–513. [Google Scholar] [CrossRef]
- Ishii, S.; Ksoll, W.B.; Hicks, R.E.; Sadowsky, M.J. Presence and Growth of Naturalized Escherichia coli in Temperate Soils from Lake Superior Watersheds. Appl. Environ. Microbiol. 2006, 72, 612–621. [Google Scholar] [CrossRef]
- Topp, E.; Welsh, M.; Tien, Y.-C.; Dang, A.; Lazarovits, G.; Conn, K.; Zhu, H. Strain-Dependent Variability in Growth and Survival of Escherichia coli in Agricultural Soil. FEMS Microbiol. Ecol. 2003, 44, 303–308. [Google Scholar] [CrossRef]
- Ishii, S.; Sadowsky, M.J. Escherichia coli in the Environment: Implications for Water Quality and Human Health. Microbes Environ. 2008, 23, 101–108. [Google Scholar] [CrossRef]
- Lee, C.C.Y. Genotyping Escherichia coli Isolates from Duck, Goose, and Gull Fecal Samples with Phylogenetic Markers Using Multiplex Polymerase Chain Reaction for Application in Microbial Source Tracking. J. Exp. Microbiol. Immunol. 2011, 15, 130–135. [Google Scholar]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The C Lermont Escherichia coli Phylo-typing Method Revisited: Improvement of Specificity and Detection of New Phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.K.; Pinyon, J.L.; Anantham, S.; Hall, R.M. Distribution of Human Commensal Escherichia coli Phylogenetic Groups. J. Clin. Microbiol. 2010, 48, 3455–3456. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The Link between Phylogeny and Virulence in Escherichia coli Extraintestinal Infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Walk, S.T.; Alm, E.W.; Calhoun, L.M.; Mladonicky, J.M.; Whittam, T.S. Genetic Diversity and Population Structure of Escherichia coli Isolated from Freshwater Beaches. Environ. Microbiol. 2007, 9, 2274–2288. [Google Scholar] [CrossRef]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.Z.; Gomes, T.A.T.; Amaral, L.A.; Ottoboni, L.M.M. Escherichia coli Phylogenetic Group Determination and Its Application in the Identification of the Major Animal Source of Fecal Contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef]
- Coura, F.M.; de Araújo Diniz, S.; Silva, M.X.; Mussi, J.M.S.; Barbosa, S.M.; Lage, A.P.; Heinemann, M.B. Phylogenetic Group Determination of Escherichia coli Isolated from Animals Samples. Sci. World J. 2015, 2015, 258424. [Google Scholar]
- Clermont, O.; Dixit, O.V.A.; Vangchhia, B.; Condamine, B.; Dion, S.; Bridier-Nahmias, A.; Denamur, E.; Gordon, D. Characterization and Rapid Identification of Phylogroup G in Escherichia coli, a Lineage with High Virulence and Antibiotic Resistance Potential. Environ. Microbiol. 2019, 21, 3107–3117. [Google Scholar] [CrossRef]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The Population Genetics of Commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Bhowmik, A.; Goswami, S.; Ahsan, S. Microbiological Quality of Prawns Collected from Local Markets of Dhaka Metropolis. Bangladesh J. Microbiol. 2018, 35, 13–16. [Google Scholar] [CrossRef]
- Bhowmik, A.; Ahsan, S. Isolation and Enumeration of Escherichia coli from Soil and Water. Bangladesh J. Microbiol. 2019, 36, 75–77. [Google Scholar] [CrossRef]
- Godambe, L.P.; Bandekar, J.; Shashidhar, R. Species Specific PCR Based Detection of Escherichia coli from Indian Foods. 3 Biotech 2017, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Kohler, B.; Oswald, E.; Beutin, L.; Karch, H.; Morabito, S.; Caprioli, A.; Suerbaum, S.; Schmidt, H. Genetic Diversity of Intimin Genes of Attaching and Effacing Escherichia coli Strains. J. Clin. Microbiol. 2002, 40, 4486–4492. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef]
- Pakbin, B.; Mahmoudi, R.; Mousavi, S.; Allahyari, S.; Amani, Z.; Peymani, A.; Qajarbeygi, P.; Hoseinabadi, Z. Genotypic and Antimicrobial Resistance Characterizations of Cronobacter Sakazakii Isolated from Powdered Milk Infant Formula: A Comparison between Domestic and Imported Products. Food Sci. Nutr. 2020, 8, 6708–6717. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. Ecological Diversity; John Wiley and Sons: New York, NY, USA, 1975. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Escobar-Páramo, P.; Le Menac’h, A.; Le Gall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of Forces Shaping the Commensal Escherichia coli Genetic Structure by Comparing Animal and Human Isolates. Environ. Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef]
- Lee, S.; Yu, J.K.; Park, K.; Oh, E.-J.; Kim, S.-Y.; Park, Y.-J. Phylogenetic Groups and Virulence Factors in Pathogenic and Commensal Strains of Escherichia coli and Their Association with BlaCTX-M. Ann. Clin. Lab. Sci. 2010, 40, 361–367. [Google Scholar] [PubMed]
- Skurnik, D.; Bonnet, D.; Bernède-Bauduin, C.; Michel, R.; Guette, C.; Becker, J.; Balaire, C.; Chau, F.; Mohler, J.; Jarlier, V. Characteristics of Human Intestinal Escherichia coli with Changing Environments. Environ. Microbiol. 2008, 10, 2132–2137. [Google Scholar] [CrossRef]
- Anastasi, E.M.; Matthews, B.; Stratton, H.M.; Katouli, M. Pathogenic Escherichia coli Found in Sewage Treatment Plants and Environmental Waters. Appl. Environ. Microbiol. 2012, 78, 5536–5541. [Google Scholar] [CrossRef]
- Escobar-Páramo, P.; Grenet, K.; Le Menac’h, A.; Rode, L.; Salgado, E.; Amorin, C.; Gouriou, S.; Picard, B.; Rahimy, M.C.; Andremont, A. Large-Scale Population Structure of Human Commensal Escherichia coli Isolates. Appl. Environ. Microbiol. 2004, 70, 5698–5700. [Google Scholar] [CrossRef]
- Stoppe, N.d.C.; Silva, J.S.; Carlos, C.; Sato, M.I.Z.; Saraiva, A.M.; Ottoboni, L.M.M.; Torres, T.T. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front. Microbiol. 2017, 8, 2512. [Google Scholar] [CrossRef] [PubMed]
- Son, I.; Van Kessel, J.A.S.; Karns, J.S. Genotypic Diversity of Escherichia coli in a Dairy Farm. Foodborne Pathog. Dis. 2009, 6, 837–847. [Google Scholar] [CrossRef] [PubMed]
- NandaKafle, G.; Seale, T.; Flint, T.; Nepal, M.; Venter, S.N.; Brözel, V.S. Distribution of Diverse Escherichia coli between Cattle and Pasture. Microbes Environ. 2017, 32, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Bergholz, P.W.; Noar, J.D.; Buckley, D.H. Environmental Patterns Are Imposed on the Population Structure of Escherichia coli after Fecal Deposition. Appl. Environ. Microbiol. 2011, 77, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Tymensen, L.D.; Pyrdok, F.; Coles, D.; Koning, W.; McAllister, T.A.; Jokinen, C.C.; Dowd, S.E.; Neumann, N.F. Comparative Accessory Gene Fingerprinting of Surface Water Escherichia coli Reveals Genetically Diverse Naturalized Population. J. Appl. Microbiol. 2015, 119, 263–277. [Google Scholar] [CrossRef]
- Berthe, T.; Ratajczak, M.; Clermont, O.; Denamur, E.; Petit, F. Evidence for Coexistence of Distinct Escherichia coli Populations in Various Aquatic Environments and Their Survival in Estuary Water. Appl. Environ. Microbiol. 2013, 79, 4684–4693. [Google Scholar] [CrossRef]
- Méric, G.; Kemsley, E.K.; Falush, D.; Saggers, E.J.; Lucchini, S. Phylogenetic Distribution of Traits Associated with Plant Colonization in Escherichia coli. Environ. Microbiol. 2013, 15, 487–501. [Google Scholar] [CrossRef]
- Abreham, S.; Teklu, A.; Cox, E.; Sisay Tessema, T. Escherichia coli O157: H7: Distribution, Molecular Characterization, Antimicrobial Resistance Patterns and Source of Contamination of Sheep and Goat Carcasses at an Export Abattoir, Mojdo, Ethiopia. BMC Microbiol. 2019, 19, 215. [Google Scholar] [CrossRef]
- Johnsen, G.; Wasteson, Y.; Heir, E.; Berget, O.I.; Herikstad, H. Escherichia coli O157: H7 in Faeces from Cattle, Sheep and Pigs in the Southwest Part of Norway during 1998 and 1999. Int. J. Food Microbiol. 2001, 65, 193–200. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source | Collection Sites |
|---|---|
| Human (n = 60) | Clinical samples from Medinova Diagnostic Centre, Dhaka, Bangladesh |
| Animal (n = 80) | Fecal specimen of animals |
| Prawn (n = 65) | Collected from local markets of Dhaka city |
| Soil (n = 85) | Different locations in Dhaka, Bangladesh |
| Water (n = 93) | Surface (n = 33) and tap water (n = 60), Dhaka, Bangladesh |
| Antibiotic Class | Antibiotic Agent | n (%) a | |||||
|---|---|---|---|---|---|---|---|
| Human (n = 60) | Animal (n = 54) | Prawn (n = 32) | Soil (n = 26) | Water (n = 2) | Total (n = 197) | ||
| Nitroheterocyclincs | Nitrofurantoin | 2 (3.3) | 15 (27.7) | 8 (25.0) | 0 (0) | 1 (4.0) | 26 (13.2) |
| Phenicols | Chloramphenicol | 2 (3.3) | 23 (42.5) | 0 (0) | 0 (0) | 3 (12.0) | 28 (14.2) |
| Tetracyclines | Tetracycline | 22 (36.6) | 35 (64.8) | 7 (21.8) | 4 (15.3) | 3 (12.0) | 71 (36.0) |
| B-lactams | Amoxicillin-Clavulanic acid | 13 (21.6) | 31 (57.4) | 4 (12.5) | 1 (3.8) | 4 (16.0) | 53 (26.9) |
| Ceftriaxone | 23 (38.3) | 2 (3.7) | 2 (6.2) | 3 (11.5) | 7 (28.0) | 37 (18.7) | |
| Quinolones | Ciprofloxacin | 34 (56.6) | 21 (38.8) | 5 (15.6) | 1 (3.8) | 5 (20.0) | 66 (33.5) |
| Aminoglycosides | Gentamicin | 21 (35.0) | 1 (1.8) | 5 (15.6) | 2 (7.6) | 6 (24.0) | 35 (17.7) |
| Macrolides | Azithromycin | 36 (60.0) | 10 (18.5) | 8 (25.0) | 4 (15.3) | 6 (24.0) | 64 (32.4) |
| Sample Source | Multi-Drug Resistance | Association | ||
|---|---|---|---|---|
| Resistant to ≥Three Classes of Drugs | Resistant to <Three Classes of Drugs | χ2 | p-Value | |
| Human | 31 (51.67) | 29 (48.33) | 29.853 | 0.001 |
| Animal | 28 (51.85) | 26 (48.15) | ||
| Prawn | 5 (15.63) | 27 (84.38) | ||
| Soil | 2 (7.69) | 24 (92.31) | ||
| Water | 5 (20.0) | 20 (80.0) | ||
| Total | 71 (36.0) | 126 (64.0) | ||
| Phylogenetic Groups | Frequency n (%) a | Multidrug-Resistant E. coli n (%) b |
|---|---|---|
| A | 53 (26.9) | 12 (22.6) |
| B1 | 95 (48.22) | 28 (29.5) |
| B2 | 4 (2.03) | 1 (25.0) |
| C | 17 (8.63) | 11 (64.7) |
| D | 18 (9.14) | 11 (61.1) |
| E | 6 (3.05) | 4 (66.7) |
| F | 4 (2.03) | 4 (100.0) |
| Samples | Phylogenetic Groups, N (%) | Diversity Indexes | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A | B1 | B2 | C | D | E | F | Shannon | Simpson | |
| Human (n = 60) | 4 (7) | 32 (53) * | 3 (5) | 0 | 16 (27) * | 1 (2) | 4 (7) * | 1.27 | 0.64 |
| Animal (n = 54) | 30 (56) * | 4 (7) | 0 | 13 (24) * | 2 (4) | 5 (9) * | 0 | 1.2 | 0.63 |
| Prawn (n = 32) | 5 (16) | 25 (78) * | 0 | 2 (6) | 0 | 0 | 0 | 0.66 | 0.37 |
| Soil (n = 26) | 4 (15) | 21 (81) * | 1 (4) | 0 | 0 | 0 | 0 | 0.27 | 0.15 |
| Water (n = 25) | 10 (40) | 13 (52) | 0 | 2 (8) | 0 | 0 | 0 | 0.91 | 0.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhowmik, A.; Shah, S.T.; Goswami, S.; Sirajee, A.S.; Ahsan, S. Predominance of Multidrug Resistant Escherichia coli of Environmental Phylotype in Different Environments of Dhaka, Bangladesh. Trop. Med. Infect. Dis. 2023, 8, 226. https://doi.org/10.3390/tropicalmed8040226
Bhowmik A, Shah ST, Goswami S, Sirajee AS, Ahsan S. Predominance of Multidrug Resistant Escherichia coli of Environmental Phylotype in Different Environments of Dhaka, Bangladesh. Tropical Medicine and Infectious Disease. 2023; 8(4):226. https://doi.org/10.3390/tropicalmed8040226
Chicago/Turabian StyleBhowmik, Anindita, SM Tanjil Shah, Sharmistha Goswami, Ahmad Salman Sirajee, and Sunjukta Ahsan. 2023. "Predominance of Multidrug Resistant Escherichia coli of Environmental Phylotype in Different Environments of Dhaka, Bangladesh" Tropical Medicine and Infectious Disease 8, no. 4: 226. https://doi.org/10.3390/tropicalmed8040226
APA StyleBhowmik, A., Shah, S. T., Goswami, S., Sirajee, A. S., & Ahsan, S. (2023). Predominance of Multidrug Resistant Escherichia coli of Environmental Phylotype in Different Environments of Dhaka, Bangladesh. Tropical Medicine and Infectious Disease, 8(4), 226. https://doi.org/10.3390/tropicalmed8040226