
False Positives in Brucellosis Serology: Wrong Bait and Wrong Pond?




Abstract
1. Introduction
Brucellosis Serology: Catching Fish with the Wrong Bait in the Wrong Pond?
2. Discussion
2.1. The Wrong Bait
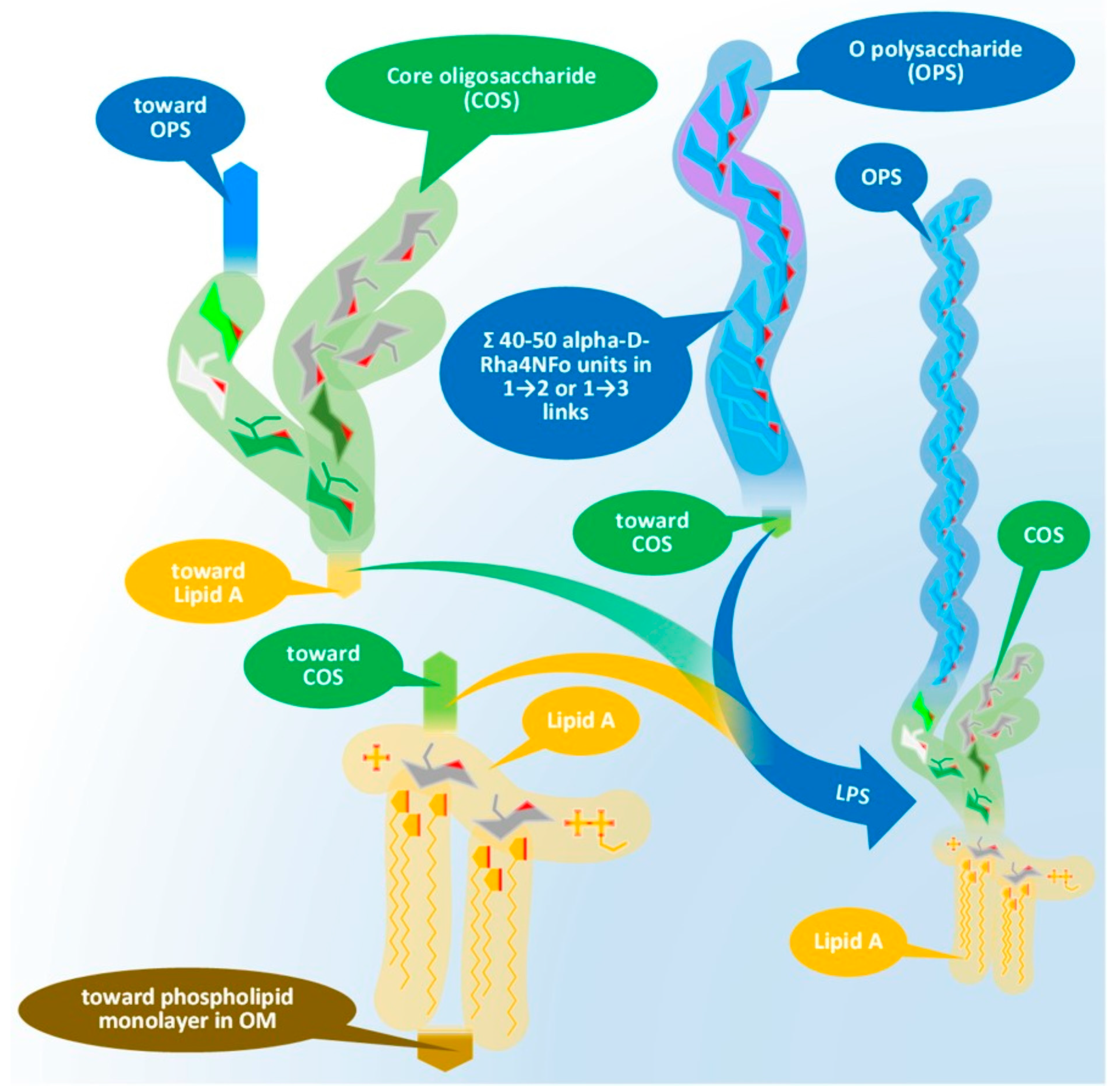
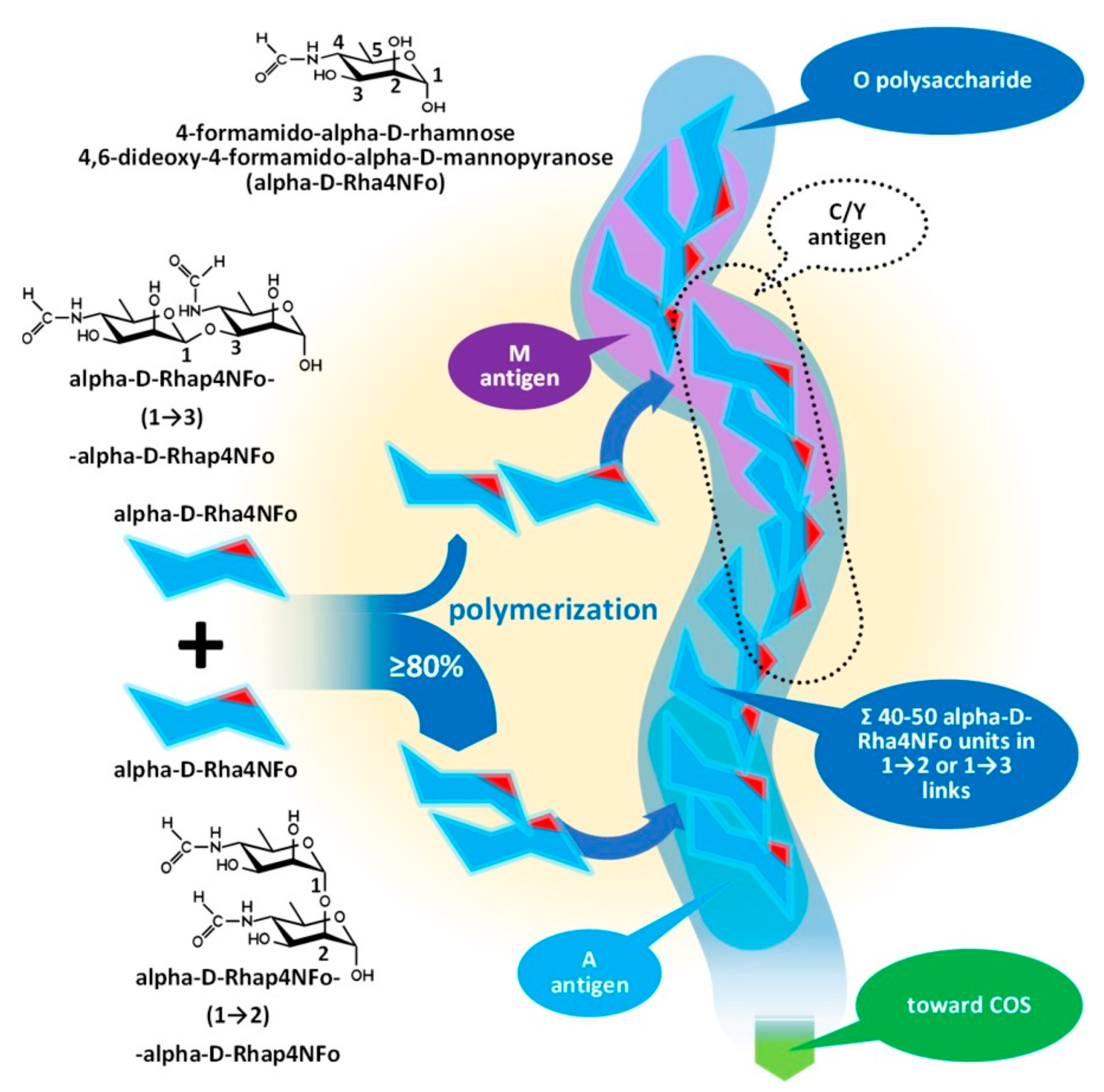
Issues with the Antigenicity of the Brucella Cell Wall
2.2. The Wrong Pond
2.2.1. Muddy Waters: Issues with Cross-Reactive Species and Other Unidentified Agents
2.2.2. Rivaling Schools of Fish: Time Lapse of Immune Reaction
2.3. Finding the Perfect Bait and/or Trying a Better Pond?
2.3.1. Finding the Perfect Bait
2.3.2. Trying a Better Pond
2.4. Stop Fishing and Start Hunting
2.4.1. cfDNA as Analyte
2.4.2. Identification of cfDNA
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herron, J.B.T.; Dunbar, J.A.T. The British Army’s contribution to tropical medicine. Clin. Med. 2018, 18, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Smith, F. On the application of the serum test to the differential diagnosis of typhoid and Malta fever: And on the further application of the method of serum diagnosis to the elucidation of certain problems in connexion with the duration of immunity and the geographical distribution of disease. Lancet 1897, 1, 656–659. [Google Scholar]
- Wright, A.E.; Semple, D. On the employment of dead bacteria in the serum diagnosis of typhoid and Malta fever, and on an easy method of extemporising a blowpipe flame for making capillary sero-sedimentation tubes. Brit. Med. J. 1897, 1, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.C.; Nöckler, K.; Göllner, C.; Bahn, P.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Kämpfer, P.; Cloeckaert, A.; Maquart, M.; et al. Brucella inopinata sp. nov., isolated from a breast implant infection. Int. J. Syst. Evol. Microbiol. 2010, 60, 801–808. [Google Scholar] [CrossRef]
- Brucellosis in Sheep and Goats-European Commission, Health & Consumer Protection Directorate-General. SANCO.C.2/AH/R23/2001. Available online: https://ec.europa.eu/food/system/files/2020-12/sci-com_scah_out59_en.pdf (accessed on 12 July 2001).
- Celli, J. The intracellular life cycle of Brucella spp. Microbiol. Spectr. 2019, 7, 101–111. [Google Scholar] [CrossRef]
- Martirosyan, A.; Moreno, E.; Gorvel, J.P. An evolutionary strategy for a stealthy intracellular Brucella pathogen. Immunol. Rev. 2011, 240, 211–234. [Google Scholar] [CrossRef]
- Barquero-Calvo, E.; Chaves-Olarte, E.; Weiss, D.S.; Guzmán-Verri, C.; Chacón-Díaz, C.; Rucavado, A.; Moriyón, I.; Moreno, E. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS ONE 2007, 2, e631. [Google Scholar] [CrossRef]
- Leong, D.; Diaz, R.; Milner, K.; Rudbach, J.; Wilson, J.B. Some structural and biological properties of Brucella endotoxin. Infect. Immun. 1970, 1, 174–182. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (WOAH). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2022, Brucellosis (Infection with Brucella abortus, B. melitensis and B. suis). Chapter 3.1.4.B.2. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.01.04_BRUCELLOSIS.pdfs (accessed on 25 May 2022).
- Wise, B.; Craig, H.W. The Brucella complement fixation reaction. J. Infect. Dis. 1942, 70, 147–151. [Google Scholar] [CrossRef]
- Gilsdorf, M.J.; Thoen, C.O.; Temple, R.M.; Gidlewski, T.; Ewalt, D.; Martin, B.; Henneger, S.B. Experimental exposure of llamas (Lama glama) to Brucella abortus; humoral antibody response. Vet. Microbiol. 2001, 81, 85–91. [Google Scholar] [CrossRef]
- Ducrotoy, M.J.; Muñoz, P.M.; Conde-Álvarez, R.; Blasco, J.M.; Moriyón, I. A systematic review of current immunological tests for the diagnosis of cattle brucellosis. Prev. Vet. Med. 2018, 151, 57–72. [Google Scholar] [CrossRef]
- Ducrotoy, M.J.; Conde-Álvarez, R.; Blasco, J.M.; Moriyón, I. A review of the basis of the immunological diagnosis of ruminant brucellosis. Vet. Immunol. Immunopathol. 2016, 171, 81–102. [Google Scholar] [CrossRef]
- Muñoz, P.M.; Marín, C.M.; Monreal, D.; González, D.; Garin-Bastuji, B.; Díaz, R.; Mainar-Jaime, R.C.; Moriyón, I.; Blasco, J.M. Efficacy of several serological tests and antigens for diagnosis of bovine brucellosis in the presence of false-positive serological results due to Yersinia enterocolitica O:9. Clin. Diagn. Lab. Immunol. 2005, 2, 141–151. [Google Scholar] [CrossRef]
- El-Tantawy, L.A.; Farahat, L.F.; Ramadan, E.S.; Warda, F.F. Effects of chemically pretreated bovine serum samples on sensitivity of Rose Bengal test for serodiagnosis of bovine brucellosis. J. Appl. Vet. Sci. 2020, 5, 6–12. [Google Scholar] [CrossRef]
- Corbel, M.J. Studies on the mechanism of the Rose Bengal plate test for bovine brucellosis. Br. Vet. J. 1973, 129, 157–166. [Google Scholar] [CrossRef]
- Macmillan, A.P.; Cockrem, D.S. Reduction of non-specific reactions to the Brucella abortus serum agglutination test by the addition of EDTA. Res. Vet. Sci. 1985, 38, 288–291. [Google Scholar] [CrossRef]
- McMahon, K.J. Comparison of the 2-mercaptoethanol and dithiothreitol tests for determining Brucella immunoglobulin G agglutinating antibody in bovine serum. Can. J. Comp. Med. 1983, 47, 370–372. [Google Scholar]
- Bonfini, B.; Chiarenza, G.; Paci, V.; Sacchini, F.; Salini, R.; Vesco, G.; Villari, S.; Zilli, K.; Tittarelli, M. Cross-reactivity in serological tests for brucellosis: A comparison of immune response of Escherichia coli O157:H7 and Yersinia enterocolitica O:9 vs. Brucella spp. Vet. Ital. 2018, 54, 107–114. [Google Scholar] [CrossRef]
- Muñoz, P.M.; Marín, C.M.; Monreal, D.; González, D.; Garin-Bastuji, B.; Díaz, R.; Mainar-Jaime, R.C.; Moriyón, I.; Blasco, J.M. Importance of screening household members of acute brucellosis cases in endemic areas. Epidemiol. Infect. 2004, 132, 533–540. [Google Scholar] [CrossRef]
- Gammoudi, I.; Mathelie-Guinlet, M.; Morote, F.; Beven, L.; Moynet, D.; Grauby-Heywang, C.; Cohen-Bouhacina, T. Morphological and nanostructural surface changes in Escherichia coli over time, monitored by atomic force microscopy. Colloids Surf. B Biointerfaces 2016, 141, 355–364. [Google Scholar] [CrossRef]
- Huber, M.; Kalis, C.; Keck, S.; Jiang, Z.; Georgel, P.; Du, X.; Shamel, L.; Sovath, S.; Mudd, S.; Beutler, B.; et al. R-form LPS, the master key to the activation ofTLR4/MD-2-positive cells. Eur. J. Immunol. 2006, 36, 701–711. [Google Scholar] [CrossRef]
- Han, X.; Tong, Y.; Tian, M.; Sun, X.; Wang, S.; Ding, C.; Yu, S. Characterization of the immunogenicity and pathogenicity of malate dehydrogenase in Brucella abortus. World J. Microbiol. Biotechnol. 2014, 30, 2063–2070. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.R.; Dorneles, E.M.S.; Pires, S.D.F.; de Andrade, H.M.; Lage, A.P. Immunoproteomics of Brucella abortus reveals potential of recombinant antigens for discriminating vaccinated from naturally infected cattle. Microb. Pathog. 2020, 147, 104345. [Google Scholar] [CrossRef] [PubMed]
- Hop, H.T.; Arayan, L.T.; Simborio, H.L.; Reyes, A.W.; Min, W.; Lee, H.J.; Lee, J.J.; Chang, H.H.; Kim, S. An evaluation of ELISA using recombinant Brucella abortus bacterioferritin (Bfr) for bovine brucellosis. Comp. Immunol. Microbiol. Infect. Dis. 2016, 45, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Nagalingam, M.; Basheer, T.J.; Balamurugan, V.; Shome, R.; Kumari, S.S.; Reddy, G.; Shome, B.R.; Rahman, H.; Roy, P.; Kingston, J.J.; et al. Comparative evaluation of the immunodominant proteins of Brucella abortus for the diagnosis of cattle brucellosis. Vet. World 2021, 14, 803–812. [Google Scholar] [CrossRef]
- Bulashev, A.; Akibekov, O.; Syzdykova, A.; Suranshiyev, Z.; Ingirbay, B. Use of recombinant Brucella outer membrane proteins 19, 25, and 31 for serodiagnosis of bovine brucellosis. Vet. World 2020, 13, 1439–1447. [Google Scholar] [CrossRef]
- Vatankhah, M.; Beheshti, N.; Mirkalantari, S.; Khoramabadi, N.; Aghababa, H.; Mahdavi, M. Recombinant Omp2b antigen-based ELISA is an efficient tool for specific serodiagnosis of animal brucellosis. Braz. J. Microbiol. 2019, 50, 979–984. [Google Scholar] [CrossRef]
- Koyuncu, I.; Kocyigit, A.; Ozer, A.; Selek, S.; Kirmit, A.; Karsen, H. Diagnostic potential of Brucella melitensis Rev1 native Omp28 precursor in human brucellosis. Central Eur. J. Immunol. 2018, 43, 81–89. [Google Scholar] [CrossRef]
- Vassen, V.; Valotteau, C.; Feuillie, C.; Formosa-Dague, C.; Dufrêne, Y.F.; De Bolle, X. Localized incorporation of outer membrane components in the pathogen Brucella abortus. EMBO J. 2019, 38, e100323. [Google Scholar] [CrossRef]
- Bohl, H.O.; Aihara, H. Current progress in the structural and biochemical characterization of proteins involved in the assembly of lipopolysaccharide. Int. J. Microbiol. 2018, 2018, 5319146. [Google Scholar] [CrossRef]
- Trent, M.S.; Stead, C.M.; Tran, A.X.; Hankins, J.V. Diversity of endotoxin and its impact on pathogenesis. J. Endotoxin Res. 2006, 12, 205–223. [Google Scholar] [CrossRef]
- Ahmed, W.; Zheng, K.; Liu, Z.F. Establishment of chronic infection: Brucella’s stealth strategy. Front. Cell. Infect. Microbiol. 2016, 6, 30. [Google Scholar] [CrossRef]
- Ferguson, G.P.; Datta, A.; Baumgartner, J.; Roop, R.M.; Carlson, R.W.; Walker, G.C. Similarity to peroxisomal-membrane protein family reveals that Sinorhizobium and Brucella BacA affect lipid-A fatty acids. Proc. Natl. Acad. Sci. USA 2004, 101, 5012–5017. [Google Scholar] [CrossRef]
- Moreno, E.; Stackebrandt, E.; Dorsch, M.; Wolters, J.; Busch, M.; Mayer, H. Brucella abortus 16S rRNA and lipid A reveal a phylogenetic relationship with members of the alpha-2 subdivision of the class Proteobacteria. J. Bacteriol. 1990, 172, 3569–3576. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Macedo, G.C.; Azevedo, V.; Oliveira, S.C. Brucella spp. noncanonical LPS: Structure, biosynthesis, and interaction with host immune system. Microb. Cell. Factories 2006, 5, 13. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Young, G.M.; Solnick, J.V.; Bäumler, A.J. From bench to bedside: Stealth of enteroinvasive pathogens. Nat. Rev. Microbiol. 2008, 6, 883–892. [Google Scholar] [CrossRef]
- Caroff, M.; Novikov, A. Lipopolysaccharides: Structure, function and bacterial identification. OCL 2020, 27, 31. [Google Scholar] [CrossRef]
- Lapaque, N.; Moriyon, I.; Moreno, E.; Gorvel, J.P. Brucella lipopolysaccharide acts as a virulence factor. Curr. Opin. Microbiol. 2005, 8, 60–66. [Google Scholar] [CrossRef]
- Forestier, C.; Moreno, E.; Méresse, S.; Phalipon, A.; Olive, D.; Sansonetti, P.; Gorvel, J.-P. Interaction of Brucella abortus lipopolysaccharide with major histocompatibility complex class II molecules in B lymphocytes. Infect. Immun. 1999, 67, 4048–4054. [Google Scholar] [CrossRef]
- Wilkinson, S.G. Bacterial lipopolysaccharides—Themes and variations. Prog. Lipid Res. 1996, 35, 283–343. [Google Scholar] [CrossRef]
- Frirdich, E.; Whitfield, C. Lipopolysaccharide inner core oligosaccharide structure and outer membrane stability in human pathogens belonging to the Enterobacteriaceae. J. Endotoxin Res. 2005, 11, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ramírez, Y.; Conde-Álvarez, R.; Palacios-Chaves, L.; Zúñiga-Ripa, A.; Grilló, M.J.; Arce-Gorvel, V.; Hanniffy, S.; Moriyón, I.; Iriarte, M. The identification of wadB, a new glycosyltransferase gene, confirms the branched structure and the role in virulence of the lipopolysaccharide core of Brucella abortus. Microb. Pathog. 2014, 73, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.; Conde-Álvarez, R.; Ståhle, J.; Holst, O.; Iriarte, M.; Zhao, Y.; Arce-Gorvel, V.; Hanniffy, S.; Gorvel, J.-P.; Moriyón, I.; et al. Structural studies of lipopolysaccharide-defective mutants from Brucella melitensis identify a core oligosaccharide critical in virulence. J. Biol. Chem. 2016, 291, 7727–7741. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.; Conde-Álvarez, R.; Ståhle, J.; Holst, O.; Iriarte, M.; Zhao, Y.; Arce-Gorvel, V.; Hanniffy, S.; Gorvel, J.-P.; Moriyón, I.; et al. A new Brucella lipopolysaccharide core glycosyltransferase identified by genomic search and phenotypic characterization. Front. Microbiol. 2018, 9, 2293. [Google Scholar] [CrossRef]
- Qureshi, N.; Takayama, K.; Mascagni, P.; Honovich, J.; Wong, R.; Cotter, R.J. Complete structural determination of lipopolysaccharide obtained from deep rough mutant of Escherichia coli. Purification by high performance liquid chromatography and direct analysis by plasma desorption mass spectrometry. J. Biol. Chem. 1988, 263, 11971–11976. [Google Scholar] [CrossRef]
- DebRoy, C.; Fratamico, P.M.; Yan, X.; Baranzoni, G.; Liu, Y.; Needleman, D.S.; Tebbs, R.; O’Connell, C.D.; Allred, A.; Swimley, M.; et al. Comparison of O-antigen gene clusters of all O-serogroups of Escherichia coli and proposal for adopting a new nomenclature for O-typing. PLoS ONE 2016, 11, e0147434. [Google Scholar] [CrossRef]
- Stenutz, R.; Weintraub, A.; Widmalm, G. The structures of Escherichia coli O-polysaccharide antigens. FEMS Microbiol. Rev. 2006, 30, 382–403. [Google Scholar] [CrossRef]
- McGiven, J.; Howells, L.; Duncombe, L.; Stack, J.; Ganesh, N.V.; Guiard, J.; Bundle, D.R. Improved serodiagnosis of bovine brucellosis by novel synthetic oligosaccharide antigens representing the capping m epitope elements of Brucella O-polysaccharide. J. Clin. Microbiol. 2015, 53, 1204–1210. [Google Scholar] [CrossRef]
- Wilson, G.S.; Miles, A.A. The serological differentiation of smooth strains of the Brucella group. Br. J. Exp. Pathol. 1932, 13, 1–13. [Google Scholar]
- Kaufmann, F. Zur serologie der Coli gruppe. Acta Pathol. Microbiol. Scand. 2009, 21, 20–45. [Google Scholar] [CrossRef]
- Kauffmann, F. Ueber einen neuen serologischen formenwechsel der Typhusbacillen. Z. Hyg. Infektionskr. 1935, 116, 617–651. [Google Scholar] [CrossRef]
- Kauffmann, F. Untersuchungen uber die korperantigene in der Salmonella-gruppe. Z. Hyg. Infektionskr. 1936, 117, 778–791. [Google Scholar] [CrossRef]
- Kauffmann, F. On the serology of the Klebsiella group. Acta Pathol. Microbiol. Scand. 1949, 26, 381–406. [Google Scholar] [CrossRef]
- Bundle, D.R.; McGiven, J. Brucellosis: Improved diagnostics and vaccine insights from synthetic glycans. Acc. Chem. Res. 2017, 50, 2958–2967. [Google Scholar] [CrossRef]
- Kubler-Kielb, J.; Vinogradov, E. Reinvestigation of the structure of Brucella O-antigens. Carbohydr. Res. 2013, 378, 144–147. [Google Scholar] [CrossRef]
- Zaccheus, M.V.; Ali, T.; Cloeckaert, A.; Zygmunt, M.S.; Weintraub, A.; Iriarte, M.; Moriyón, I.; Widmalm, G. The epitopic and structural characterization of Brucella suis biovar 2 O-polysaccharide demonstrates the existence of a new M-negative C-negative smooth Brucella serovar. PLoS ONE 2013, 8, e53941. [Google Scholar] [CrossRef]
- Ganesh, N.V.; Sadowska, J.M.; Sarkar, S.; Howells, L.; McGiven, J.; Bundle, D.R. Molecular recognition of Brucella A and M antigens dissected by synthetic oligosaccharide glycoconjugates leads to a disaccharide diagnostic for brucellosis. J. Am. Chem. Soc. 2014, 136, 16260–16269. [Google Scholar] [CrossRef]
- Zygmunt, M.S.; Bundle, D.R.; Ganesh, N.V.; Guiard, J.; Cloeckaert, A. Monoclonal antibody-defined specific C epitope of Brucella O-polysaccharide revisited. Clin. Vaccine Immunol. 2015, 22, 979–982. [Google Scholar] [CrossRef]
- Dieste-Pérez, L.; Blasco, J.M.; de Miguel, M.J.; Moriyón, I.; Muñoz, P.M. Diagnostic performance of serological tests for swine brucellosis in the presence of false positive serological reactions. J. Microbiol. Methods 2015, 111, 57–63. [Google Scholar] [CrossRef]
- Chenais, E.; Bagge, E.; Lambertz, S.T.; Artursson, K. Yersinia enterocolitica serotype O:9 cultured from Swedish sheep showing serologically false-positive reactions for Brucella melitensis. Infect. Ecol. Epidemiol. 2012, 2, 1–7. [Google Scholar] [CrossRef]
- Corbel, M.J. The serological relationship between Brucella spp., Yersinia enterocolitica serotype IX and Salmonella serotypes of Kauffmann-White group N. J. Hyg. 1975, 75, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.C.; Soeurt, N.; Splitter, G. Molecular and cellular interactions between Brucella abortus antigens and host immune responses. Vet. Microbiol. 2002, 90, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.M.; Islam, M.A.; Baek, B.K.; Lee, S.I. Cellular and humoral immune responses and antigen recognition in Sprague-Dawley rats experimentally infected with Brucella abortus biotype 1. Asian J. Anim. Vet. Adv. 2009, 4, 267–277. [Google Scholar] [CrossRef]
- Bercovich, Z. The use of skin delayed-type hypersensitivity as an adjunct test to diagnose brucellosis in cattle: A review. Vet. Q. 2000, 22, 123–130. [Google Scholar] [CrossRef]
- Saegerman, C.; Vo, T.-K.O.; De Waele, L.; Gilson, D.; Bastin, A.; Dubray, G.; Flanagan, P.; Limet, J.N.; Letesson, J.-J.; Godfroid, J. Diagnosis of bovine brucellosis by skin test: Conditions for the test and evaluation of its performance. Vet. Rec. 1999, 145, 214–218. [Google Scholar] [CrossRef]
- Conde-Álvarez, R.; Arce-Gorvel, V.; Iriarte, M.; Manček-Keber, M.; Barquero-Calvo, E.; Palacios-Chaves, L.; Chacón-Díaz, C.; Chaves-Olarte, E.; Martirosyan, A.; von Bargen, K.; et al. The lipopolysaccharide core of Brucella abortus acts as a shield against innate immunity recognition. PLoS Pathog. 2012, 8, e1002675. [Google Scholar] [CrossRef]
- Bienvenu, A.; Martinez, E.; Bonazzi, M. Undercover agents of infection: The stealth strategies of t4ss-equipped bacterial pathogens. Toxins 2021, 13, 713. [Google Scholar] [CrossRef]
- Cornejo, E.; Schlaermann, P.; Mukherjee, S. How to rewire the host cell: A home improvement guide for intracellular bacteria. J. Cell Biol. 2017, 216, 3931–3948. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, H.; Guo, F.; Yuan, L.; Zhang, W.; Wang, Y.; Chen, C. OMP31 of Brucella melitensis 16M impairs the apoptosis of macrophages triggered by TNF-α. Exp. Ther. Med. 2016, 12, 2783–2789. [Google Scholar] [CrossRef]
- Ma, Q.-L.; Liu, A.-C.; Ma, X.-J.; Wang, Y.-B.; Hou, Y.-T.; Wang, Z.-H. Brucella outer membrane protein Omp25 induces microglial cells in vitro to secrete inflammatory cytokines and inhibit apoptosis. Int. J. Clin. Exp. Med. 2015, 8, 17530–17535. [Google Scholar]
- De Jong, M.F.; Rolán, H.G.; Tsolis, R.M. Microreview: Innate immune encounters of the (Type) 4th kind: Brucella. Cell. Microbiol. 2010, 12, 1195–1202. [Google Scholar] [CrossRef]
- Von Bargen, K.; Gorvel, J.-P.; Salcedo, S.P. Internal affairs: Investigating the Brucella intracellular lifestyle. FEMS Microbiol. Rev. 2012, 36, 533–562. [Google Scholar] [CrossRef]
- Byndloss, M.X.; Tsolis, R.M. Brucella spp. virulence factors and immunity. Annu. Rev. Anim. Biosci. 2016, 4, 111–127. [Google Scholar] [CrossRef]
- Jiao, H.; Zhou, Z.; Li, B.; Xiao, Y.; Li, M.; Zeng, H.; Guo, X.; Gu, G. The mechanism of facultative intracellular parasitism of Brucella. Int. J. Mol. Sci. 2021, 22, 3673. [Google Scholar] [CrossRef]
- Bin Im, Y.; Bin Park, W.; Jung, M.; Kim, S.; Yoo, H.S. Evaluation of Th1/Th2-related immune response against recombinant proteins of Brucella abortus infection in mice. J. Microbiol. Biotechnol. 2016, 26, 1132–1139. [Google Scholar] [CrossRef]
- Del Pozo, J.S.G.; Ortuño, S.L.; Navarro, E.; Solera, J. Detection of IgM antibrucella antibody in the absence of IgGs: A challenge for the clinical interpretation of Brucella serology. PLoS Negl. Trop. Dis. 2014, 8, e3390. [Google Scholar] [CrossRef]
- Jindan, R.A.; Saleem, N.; Shafi, A.; Amjad, S.M. Clinical interpretation of detection of IgM anti-Brucella antibody in the absence of IgG and vice versa; a diagnostic challenge for clinicians. Pol. J. Microbiol. 2019, 68, 51–57. [Google Scholar] [CrossRef]
- Kumar, S.; Tuteja, U.; Kumar, A.; Batra, H.V. Expression and purification of the 26 kDa periplasmic protein of Brucella abortus: A reagent for the diagnosis of bovine brucellosis. Biotechnol. Appl. Biochem. 2008, 49, 213–218. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Baucheron, S.; Vizcaino, N.; Zygmunt, M.S.; Cole, A.M.; Tahk, S.; Oren, A.; Yoshioka, D.; Kim, Y.-H.; Park, A.; et al. Use of recombinant BP26 protein in serological diagnosis of Brucella melitensis infection in sheep. Clin. Diagn. Lab. Immunol. 2001, 8, 1064–1069. [Google Scholar] [CrossRef]
- Letesson, J.J.; Tibor, A.; van Eynde, G.; Wansard, V.; Weynants, V.; Denoel, P.; Saman, E. Humoral immune responses of Brucella-infected cattle, sheep, and goats to eight purified recombinant Brucella proteins in an indirect enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol. 1997, 4, 556–564. [Google Scholar] [CrossRef]
- Bulashev, A.K.; Ingirbay, B.K.; Mukantayev, K.N.; Syzdykova, A.S. Evaluation of chimeric proteins for serological diagnosis of brucellosis in cattle. Vet. World 2021, 14, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Bai, Q.; Wu, X.; Li, H.; Shao, J.; Sun, M.; Zhang, J. A multi-epitope fusion protein-based p-ELISA method for diagnosing bovine and goat brucellosis. Front. Vet. Sci. 2021, 8, 708008. [Google Scholar] [CrossRef] [PubMed]
- Corrente, M.; Desario, C.; Parisi, A.; Grandolfo, E.; Scaltrito, D.; Vesco, G.; Colao, V.; Buonavoglia, D. Serological diagnosis of bovine brucellosis using B. melitensis strain B115. J. Microbiol. Methods 2015, 119, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Trotta, A.; Marinaro, M.; Cirilli, M.; Sposato, A.; Adone, R.; Beverelli, M.; Buonavoglia, D.; Corrente, M. Brucella melitensis B115-based ELISA to unravel false positive serologic reactions in bovine brucellosis: A field study. BMC Vet. Res. 2020, 16, 50. [Google Scholar] [CrossRef]
- Degner, N.R.; Castillo-Galvan, R.; Alexander, J.; Arun, A.; de Vries, C.R.; Macintyre, A.; Perkins, B.; Ahmed, A.A.; Smollin, M. Following the hoof prints: Detecting Coxiella and Brucella infections with a plasma-based microbial cell-free DNA next-generation sequencing test. Open Forum Infect. Dis. 2021, 8 (Suppl. S1), S604. [Google Scholar] [CrossRef]
- Kowarsky, M.; Camunas-Soler, J.; Kertesz, M.; De Vlaminck, I.; Koh, W.; Pan, W.; Martin, L.; Neff, N.F.; Okamoto, J.; Wong, R.J.; et al. Numerous uncharacterized and highly divergent microbes which colonize humans are revealed by circulating cell-free DNA. Proc. Natl. Acad. Sci. USA 2017, 114, 9623–9628. [Google Scholar] [CrossRef]
- Blauwkamp, T.A.; Thair, S.; Rosen, M.J.; Blair, L.; Lindner, M.S.; Vilfan, I.D.; Kawli, T.; Christians, F.C.; Venkatasubrahmanyam, S.; Wall, G.D.; et al. Analytical and clinical validation of a microbial cell-free DNA sequencing test for infectious disease. Nat. Microbiol. 2019, 4, 663–674. [Google Scholar] [CrossRef]
- Ramírez, J.D.; Herrera, G.; Muskus, C.; Mendez, C.; Duque, M.C.; Butcher, R. Development of a digital droplet polymerase chain reaction (ddPCR) assay to detect Leishmania DNA in samples from cutaneous leishmaniasis patients. Int. J. Infect. Dis. 2019, 79, 1–3. [Google Scholar] [CrossRef]
- Rossoff, J.; Chaudhury, S.; Soneji, M.; Patel, S.J.; Kwon, S.; Armstrong, A.; Muller, W.J. Noninvasive diagnosis of infection using plasma next-generation sequencing: A single-center experience. Open Forum Infect. Dis. 2019, 6, mofz327. [Google Scholar] [CrossRef]
- Fernández-Carballo, B.L.; Broger, T.; Wyss, R.; Banaei, N.; Denkinger, C.M. Toward the development of a circulating free DNA-based in vitro diagnostic test for infectious diseases: A review of evidence for tuberculosis. J. Clin. Microbiol. 2019, 57, e01234-18. [Google Scholar] [CrossRef]
- Han, D.; Li, R.; Shi, J.; Tan, P.; Zhang, R.; Li, J. Liquid biopsy for infectious diseases: A focus on microbial cell-free DNA sequencing. Theranostics 2020, 10, 5501–5513. [Google Scholar] [CrossRef]
- Peaper, D.R.; Durant, T.S. Can circulating cell-free microbial DNA carry us into the future of culture independent microbiology? Clin. Chem. 2020, 66, 29–32. [Google Scholar] [CrossRef]
- Pan, S.-W.; Su, W.-J.; Chan, Y.-J.; Chuang, F.-Y.; Feng, J.-Y.; Chen, Y.-M. Mycobacterium tuberculosis–derived circulating cell-free DNA in patients with pulmonary tuberculosis and persons with latent tuberculosis infection. PLoS ONE 2021, 16, e0253879. [Google Scholar] [CrossRef]
- Zheng, Y.; Jin, J.; Shao, Z.; Liu, J.; Zhang, R.; Sun, R.; Hu, B. Development and clinical validation of a droplet digital PCR assay for detecting Acinetobacter baumannii and Klebsiella pneumoniae in patients with suspected bloodstream infections. MicrobiologyOpen 2021, 10, e1247. [Google Scholar] [CrossRef]
- Shao, Z.; Zhu, J.; Wei, Y.; Jin, J.; Zheng, Y.; Liu, J.; Zhang, R.; Sun, R.; Hu, B. Pathogen load and species monitored by droplet digital PCR in patients with bloodstream infections: A prospective case series study. BMC Infect. Dis. 2022, 22, 771. [Google Scholar] [CrossRef]
- Hogan, C.A.; Yang, S.; Garner, O.B.; Green, D.A.; Gomez, C.A.; Bard, J.D.; Pinsky, B.A.; Banaei, N. Clinical impact of metagenomic next-generation sequencing of plasma cell-free DNA for the diagnosis of infectious diseases: A multicenter retrospective cohort study. Clin. Infect. Dis. 2021, 72, 239–245. [Google Scholar] [CrossRef]
- Babady, N.E. Clinical metagenomics for bloodstream infections: Is the juice worth the squeeze? Clin. Infect. Dis. 2021, 72, 246–248. [Google Scholar] [CrossRef]
- Muller, W.J.; Chaudhury, S. Utility of metagenomic next-generation sequencing of plasma for infectious pathogens. Clin. Infect. Dis. 2020, 71, 2775. [Google Scholar] [CrossRef]
- Justiz Vaillant, A.A.; Zulfiqar, H.; Ramphul, K. Delayed Hypersensitivity Reactions; StatPearls: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK519023/ (accessed on 8 May 2023).
- Mandel, P.; Metais, P. Nuclear acids in human blood plasma. C. R. Seances Soc. Biol. Fil. 1948, 142, 241–243. [Google Scholar]
- Lo, Y.M.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]
- Urbanova, M.; Plzak, J.; Strnad, H.; Betka, J. Circulating nucleic acids as a new diagnostic tool. Cell. Mol. Biol. Lett. 2010, 15, 242–259. [Google Scholar] [CrossRef] [PubMed]
- Elshimali, Y.I.; Khaddour, H.; Sarkissyan, M.; Wu, Y.; Vadgama, J.V. The clinical utilization of circulating cell free DNA (CCFDNA) in blood of cancer patients. Int. J. Mol. Sci. 2013, 14, 18925–18958. [Google Scholar] [CrossRef] [PubMed]
- El-Heliebi, A.; Heitzer, E. State of the art and future direction for the analysis of cell-free circulating DNA. In Micro and Nano Technologies, Nucleic Acid Nanotheranostics, 1st ed.; Marco Filice, M., Ruiz-Cabello, J., Eds.; Elsevier: Cambridge, MA, USA, 2019; Chapter 5; pp. 133–188. [Google Scholar] [CrossRef]
- Hassan, S.; Shehzad, A.; Khan, S.A.; Miran, W.; Khan, S.; Lee, Y.-S. Diagnostic and therapeutic potential of circulating-free DNA and cell-free RNA in cancer management. Biomedicines 2022, 10, 2047. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zeng, Q.; Wang, Z.; Li, C.; Xu, Y.; Cui, P.; Zhu, X.; Lu, H.; Wang, G.; Cai, S.; et al. Circulating cell-free DNA for cancer early detection. Innovation 2022, 3, 100259. [Google Scholar] [CrossRef] [PubMed]
- Arosemena, M.; Meah, F.A.; Mather, K.J.; Tersey, S.A.; Mirmira, R.G. Cell-free DNA fragments as biomarkers of islet β-cell death in obesity and type 2 diabetes. Int. J. Mol. Sci. 2021, 22, 2151. [Google Scholar] [CrossRef]
- Vajpeyee, A.; Wijatmiko, T.; Vajpeyee, M.; Taywade, O.; Pandey, S.; Chauhan, P.S. Clinical usefulness of cell-free DNA as a prognostic marker in acute ischemic stroke. Neurologist 2020, 2, 11–13. [Google Scholar] [CrossRef]
- Xu, Y.; Song, Y.; Chang, J.; Zhou, X.; Qi, Q.; Tian, X.; Li, M.; Zeng, X.; Xu, M.; Zhang, W.; et al. High levels of circulating cell-free DNA are a biomarker of active SLE. Eur. J. Clin. Investig. 2018, 48, e13015. [Google Scholar] [CrossRef]
- Filho, E.M.R.; Simon, D.; Ikuta, N.; Klovan, C.; Dannebrock, F.A.; de Oliveira, C.O.; Regner, A. Elevated cell-free plasma DNA level as an independent predictor of mortality in patients with severe traumatic brain injury. J. Neurotrauma 2014, 31, 1639–1646. [Google Scholar] [CrossRef]
- Briggs, G.D.; Gelzinnis, S.; Meakes, S.; King, K.L.; Balogh, Z.J. Not all cell-free mitochondrial DNA is equal in trauma patients. Shock 2022, 58, 231–235. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yoshida, K.; Hashiramoto, A.; Matsui, K. Cell-free DNA in rheumatoid arthritis. Int. J. Mol. Sci. 2021, 22, 8941. [Google Scholar] [CrossRef]
- Van der Meer, A.J.; Kroeze, A.; Hoogendijk, A.J.; Soussan, A.A.; van der Schoot, C.E.; Wuillemin, W.A.; Voermans, C.; van der Poll, T.; Zeerleder, S. Systemic inflammation induces release of cell-free DNA from hematopoietic and parenchymal cells in mice and humans. Blood Adv. 2019, 3, 724–728. [Google Scholar] [CrossRef]
- Mondelo-Macía, P.; Castro-Santos, P.; Castillo-García, A.; Muinelo-Romay, L.; Diaz-Peña, R. Circulating Free DNA and its emerging role in autoimmune diseases. J. Pers. Med. 2021, 11, 151. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, C.; Li, X.; Fu, Q.; Li, J.; Su, Q.; Zeng, L.; Liu, Z.; Wang, J.; Huang, H.; et al. Diagnostic performance of donor-derived plasma cell-free DNA fraction for antibody-mediated rejection in post renal transplant recipients: A prospective observational study. Front. Immunol. 2020, 11, 342. [Google Scholar] [CrossRef]
- Khush, K.K.; Patel, J.; Pinney, S.; Kao, A.; Alharethi, R.; DePasquale, E.; Ewald, G.; Berman, P.; Kanwar, M.; Hiller, D.; et al. Noninvasive detection of graft injury after heart transplant using donor-derived cell-free DNA: A prospective multicenter study. Am. J. Transplant. 2019, 19, 2889–2899. [Google Scholar] [CrossRef]
- Oellerich, M.; Budde, K.; Osmanodja, B.; Bornemann-Kolatzki, K.; Beck, J.; Schütz, E.; Walson, P.D. Donor-derived cell-free DNA as a diagnostic tool in transplantation. Front. Genet. 2022, 13, 1031894. [Google Scholar] [CrossRef]
- Afzal, A.; Alam, A.; van Zyl, J.S.; Zafar, H.; Felius, J.; Hall, S.A.; Carey, S.A. Observed elevated donor-derived cell free DNA in orthotopic heart transplant recipients without clinical evidence of rejection. Clin. Transplant. 2022, 36, e14549. [Google Scholar] [CrossRef]
- Hanson, B.; Scotchman, E.; Chitty, L.S.; Chandler, N.J. Non-invasive prenatal diagnosis (NIPD): How analysis of cell-free DNA in maternal plasma has changed prenatal diagnosis for monogenic disorders. Clin. Sci. 2022, 136, 1615–1629. [Google Scholar] [CrossRef]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating cell-free nucleic acids: Main characteristics and clinical application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef]
- Pös, Z.; Pös, O.; Styk, J.; Mocova, A.; Strieskova, L.; Budis, J.; Kadasi, L.; Radvanszky, J.; Szemes, T. Technical and methodological aspects of cell-free nucleic acids analyzes. Int. J. Mol. Sci. 2020, 21, 8634. [Google Scholar] [CrossRef]
- Filho, E.M.R.; Simon, D.; Ikuta, N.; Klovan, C.; Dannebrock, F.A.; de Oliveira, C.O.; Regner, A. Effects of preanalytical factors on the molecular size of cell-free DNA in blood. Clin. Chem. 2005, 51, 781–784. [Google Scholar] [CrossRef]
- Xue, X.; Teare, M.D.; Holen, I.; Zhu, Y.M.; Woll, P.J. Optimizing the yield and utility of circulating cell-free DNA from plasma and serum. Clin. Chim. Acta 2009, 404, 100–104. [Google Scholar] [CrossRef] [PubMed]
- El Messaoudi, S.; Rolet, F.; Mouliere, F.; Thierry, A.R. Circulating cell free DNA: Preanalytical considerations. Clin. Chim. Acta 2013, 424, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Page, K.; Guttery, D.S.; Zahra, N.; Primrose, L.; Elshaw, S.R.; Pringle, J.H.; Blighe, K.; Marchese, S.D.; Hills, A.; Woodley, L.; et al. Influence of plasma processing on recovery and analysis of circulating nucleic acids. PLoS ONE 2013, 8, e77963. [Google Scholar] [CrossRef] [PubMed]
- Devonshire, A.S.; Whale, A.S.; Gutteridge, A.; Jones, G.; Cowen, S.; Foy, C.A.; Huggett, J.F. Towards standardisation of cell-free DNA measurement in plasma: Controls for extraction efficiency, fragment size bias and quantification. Anal. Bioanal. Chem. 2014, 406, 6499–6512. [Google Scholar] [CrossRef]
- Markus, H.; Contente-Cuomo, T.; Farooq, M.; Liang, W.S.; Borad, M.J.; Sivakumar, S.; Gollins, S.; Tran, N.L.; Dhruv, H.D.; Berens, M.E.; et al. Evaluation of pre-analytical factors affecting plasma DNA analysis. Sci. Rep. 2018, 8, 7375. [Google Scholar] [CrossRef]
- Shiotsu, H.; Okada, K.; Shibuta, T.; Kobayashi, Y.; Shirahama, S.; Kuroki, C.; Ueda, S.; Ohkuma, M.; Ikeda, K.; Ando, Y.; et al. The influence of pre-analytical factors on the analysis of circulating microRNA. MicroRNA 2018, 7, 195–203. [Google Scholar] [CrossRef]
- Nesic, M.; Bødker, J.S.; Terp, S.K.; Dybkær, K. Optimization of preanalytical variables for cfDNA processing and detection of ctDNA in archival plasma samples. Biomed Res. Int. 2021, 2021, 5585148. [Google Scholar] [CrossRef]
- Norton, S.E.; Lechner, J.M.; Williams, T.; Fernando, M.R. A stabilizing reagent prevents cell-free DNA contamination by cellular DNA in plasma during blood sample storage and shipping as determined by digital PCR. Clin. Biochem. 2013, 46, 1561–1565. [Google Scholar] [CrossRef]
- Westermeier, R. Electrophoresis in Practice: A Guide to Methods and Applications of DNA and Protein Separations, 5th ed.; Wiley: Weinheim, Germany, 2016; pp. 1–146. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Analysis of DNA by agarose gel electrophoresis. Cold Spring Harb. Protoc. 2019, 2019, 6–15. [Google Scholar] [CrossRef]
- Yu, S.C.Y.; Chan, K.C.A.; Zheng, Y.W.L.; Jiang, P.; Liao, G.J.W.; Sun, H.; Akolekar, R.; Leung, T.Y.; Go, A.T.J.I.; van Vugt, J.M.G.; et al. Size-based molecular diagnostics using plasma DNA for noninvasive prenatal testing. Proc. Natl. Acad. Sci. USA 2014, 111, 8583–8588. [Google Scholar] [CrossRef]
- Huang, Y.; Mu, J.; Qi, L.; Ge, W.; Fang, X.; Song, Y.; Yuan, Y.; Zheng, S. Diverse fragment lengths dismiss size selection for serum cell-free DNA: A comparative study of serum and plasma samples. Clin. Chem. Lab. Med. 2020, 58, 1451–1459. [Google Scholar] [CrossRef]
- Pečnikar, Ž.F.; Buzan, E.V. 20 years since the introduction of DNA barcoding: From theory to application. J. Appl. Genet. 2014, 55, 43–52. [Google Scholar] [CrossRef]
- Rassinger, A. In vivo footprinting analysis in Trichoderma reesei. Methods Mol. Biol. 2021, 2234, 177–189. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Gu, W.; Deng, X.; Lee, M.; Sucu, Y.D.; Arevalo, S.; Stryke, D.; Federman, S.; Gopez, A.; Reyes, K.; Zorn, K.; et al. Rapid pathogen detection by metagenomic next-generation sequencing of infected body fluids. Nat. Med. 2021, 27, 115–124. [Google Scholar] [CrossRef]
- Martignano, F.; Munagala, U.; Crucitta, S.; Mingrino, A.; Semeraro, R.; Del Re, M.; Petrini, I.; Magi, A.; Conticello, S.G. Nanopore sequencing from liquid biopsy: Analysis of copy number variations from cell-free DNA of lung cancer patients. Mol. Cancer 2021, 20, 32. [Google Scholar] [CrossRef]
- Katsman, E.; Orlanski, S.; Martignano, F.; Fox-Fisher, I.; Shemer, R.; Dor, Y.; Zick, A.; Eden, A.; Petrini, I.; Conticello, S.G.; et al. Detecting cell-of-origin and cancer-specific methylation features of cell-free DNA from nanopore sequencing. Genome Biol. 2022, 23, 158. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe | Diagnostic Sensitivity (%) | Diagnostic Specificity (%) |
|---|---|---|
| RBT a [15] | 100 (98.2–100) | 86.4 (79.1–91.9) |
| CFT b [15] | 100 (98.2–100) | 94.4 (88.8–97.7) |
| iELISA c [15] | 100 (98–100) | 58.4 (49.2–67.1) |
| Approach | Antigen/Target | Origin of Antigen | Target Molecule | Detection Technology | Reference |
|---|---|---|---|---|---|
| (i) Non-LPS or non-LPS elements | Other components of the bacterium cells (proteins) | Cytoplasmic | malate dehydrogenase * | NA | [24,25] |
| superoxide dismutase * | iELISA | [25] | |||
| bacterioferritin * | iELISA | [26] | |||
| Periplasmic | protein 26 (BP26) * | iELISA | [27,80,81] | ||
| Inner membrane | ABC transporter * | iELISA | [25] | ||
| Outer membrane | Omp10 * | iELISA | [82] | ||
| Omp16 * | iELISA | [82] | |||
| Omp19 * | iELISA | [28] | |||
| Omp25 * | iELISA | [28] | |||
| Omp28 * | iELISA | [30] | |||
| Omp19-25 **, Omp19-31 **, Omp25-31 ** | iELISA | [83] | |||
| OMP31* | iELISA | [28] | |||
| OMP36 * | iELISA | [82] | |||
| Omp2b * | iELISA | [29] | |||
| Mixed origin | fusion protein of Omp16, Omp25, Omp31, Omp2b, and BP26 epitops (22 epitops) * | pELISA **** | [84] | ||
| (ii) Non-S LPS or non-S LPS elements | R LPS or R LPS element | Outer membrane isolate | NA | iELISA | [85] |
| Processed *** outer membrane isolate | NA | NA | NA | ||
| Killed cells | NA | iELISA | [86] | ||
| Artificial oligosaccharides | Synthetic | free or BSA-conjugated oligosaccharides | iELISA | [50,56,59] | |
| (iii) Alternative solutions | Brucellin | Killed cell isolates | NA | Skin test | [66] |
| Cell-free DNA | Immune processed bacteria in blood plasma | DNA | Cell-free DNA NGS | [87,88,89,90,91,92,93,94,95,96,97,98,99,100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bányász, B.; Antal, J.; Dénes, B. False Positives in Brucellosis Serology: Wrong Bait and Wrong Pond? Trop. Med. Infect. Dis. 2023, 8, 274. https://doi.org/10.3390/tropicalmed8050274
Bányász B, Antal J, Dénes B. False Positives in Brucellosis Serology: Wrong Bait and Wrong Pond? Tropical Medicine and Infectious Disease. 2023; 8(5):274. https://doi.org/10.3390/tropicalmed8050274
Chicago/Turabian StyleBányász, Borbála, József Antal, and Béla Dénes. 2023. "False Positives in Brucellosis Serology: Wrong Bait and Wrong Pond?" Tropical Medicine and Infectious Disease 8, no. 5: 274. https://doi.org/10.3390/tropicalmed8050274
APA StyleBányász, B., Antal, J., & Dénes, B. (2023). False Positives in Brucellosis Serology: Wrong Bait and Wrong Pond? Tropical Medicine and Infectious Disease, 8(5), 274. https://doi.org/10.3390/tropicalmed8050274