



Assessing the Efficacy of Monovalent and Commercialized Antivenoms for Neutralizing Moroccan Cobra Naja haje Venom: A Comparative Study
 ,
,  , and
, and
Abstract
1. Introduction
2. Materials & Methods
2.1. Venoms
2.2. Antivenoms
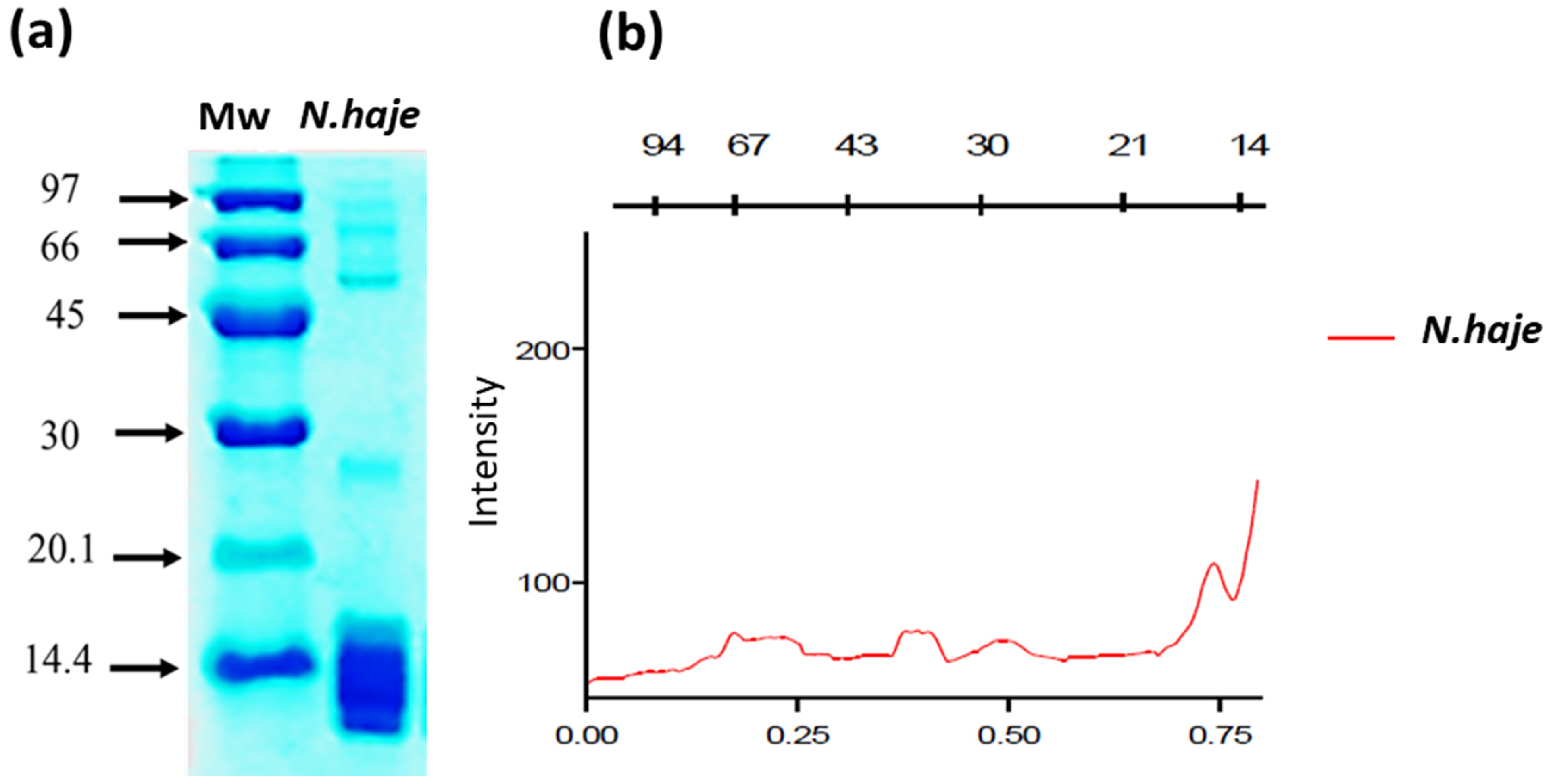
2.3. SDS-Page and Densitometric Analysis
2.4. Toxic Activities of Naja haje Venom
2.4.1. Lethality
2.4.2. Experimental Envenomation
2.5. Neutralization of Toxic Activities
2.5.1. Median Effective Dose (ED50) Determination
2.5.2. Neutralization of the Alterations Induced by Naja haje Venom
2.6. Histological Study
2.7. Animals and Ethics Statement
2.8. Statistical Analysis
3. Results
3.1. SDS-Page Analysis
3.2. Toxic Activities of the Naja haje Venom
3.2.1. Determination of the LD 50
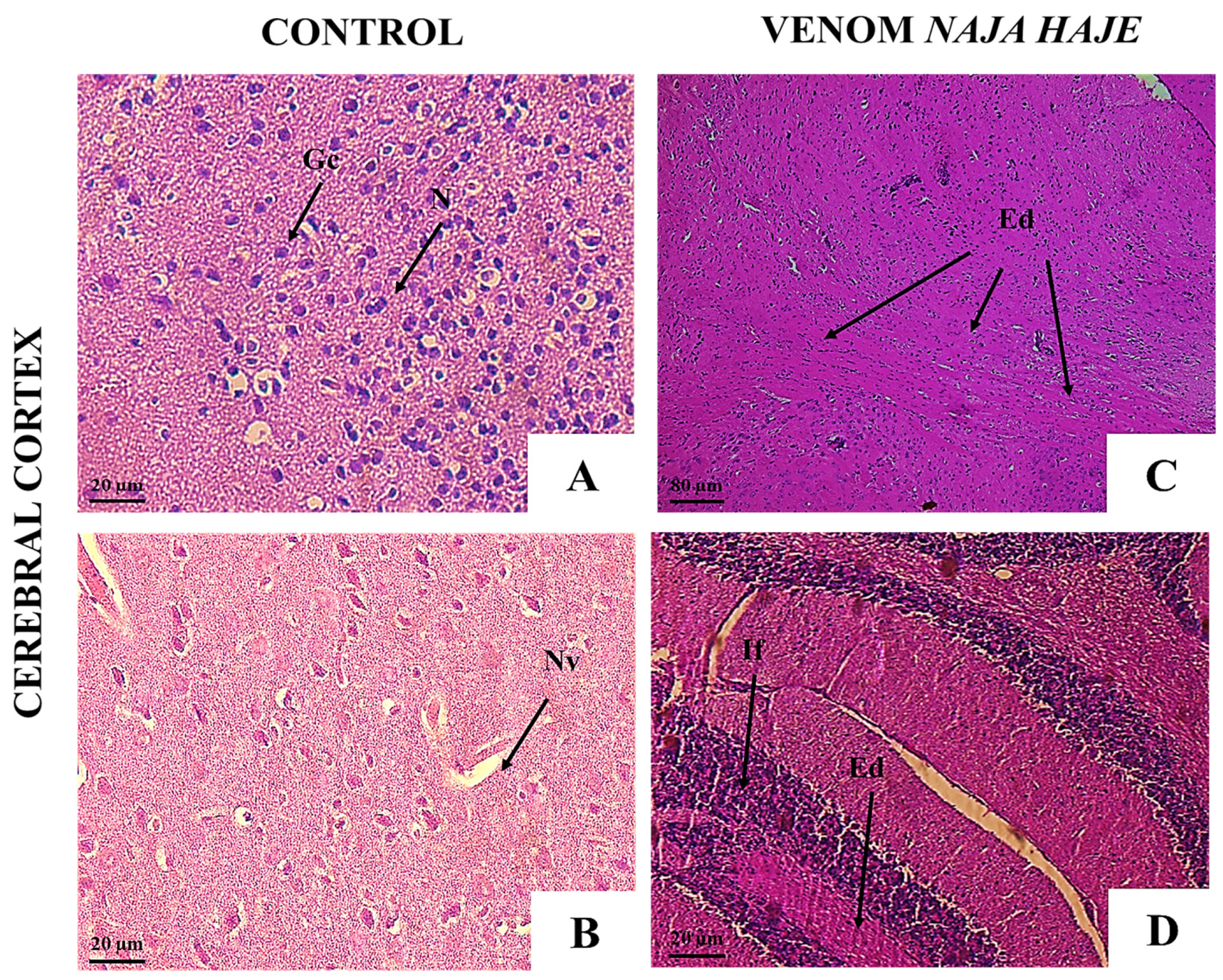
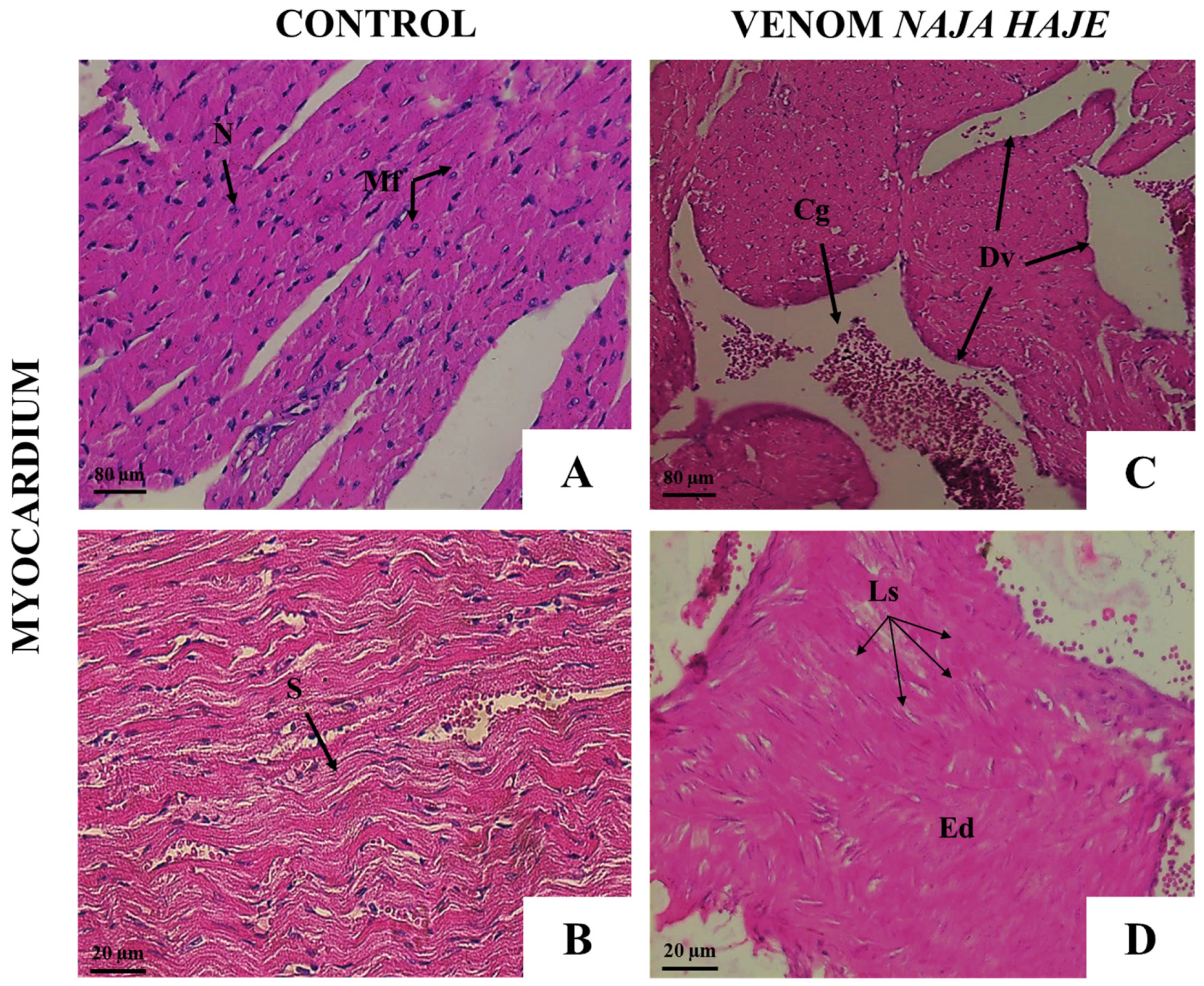
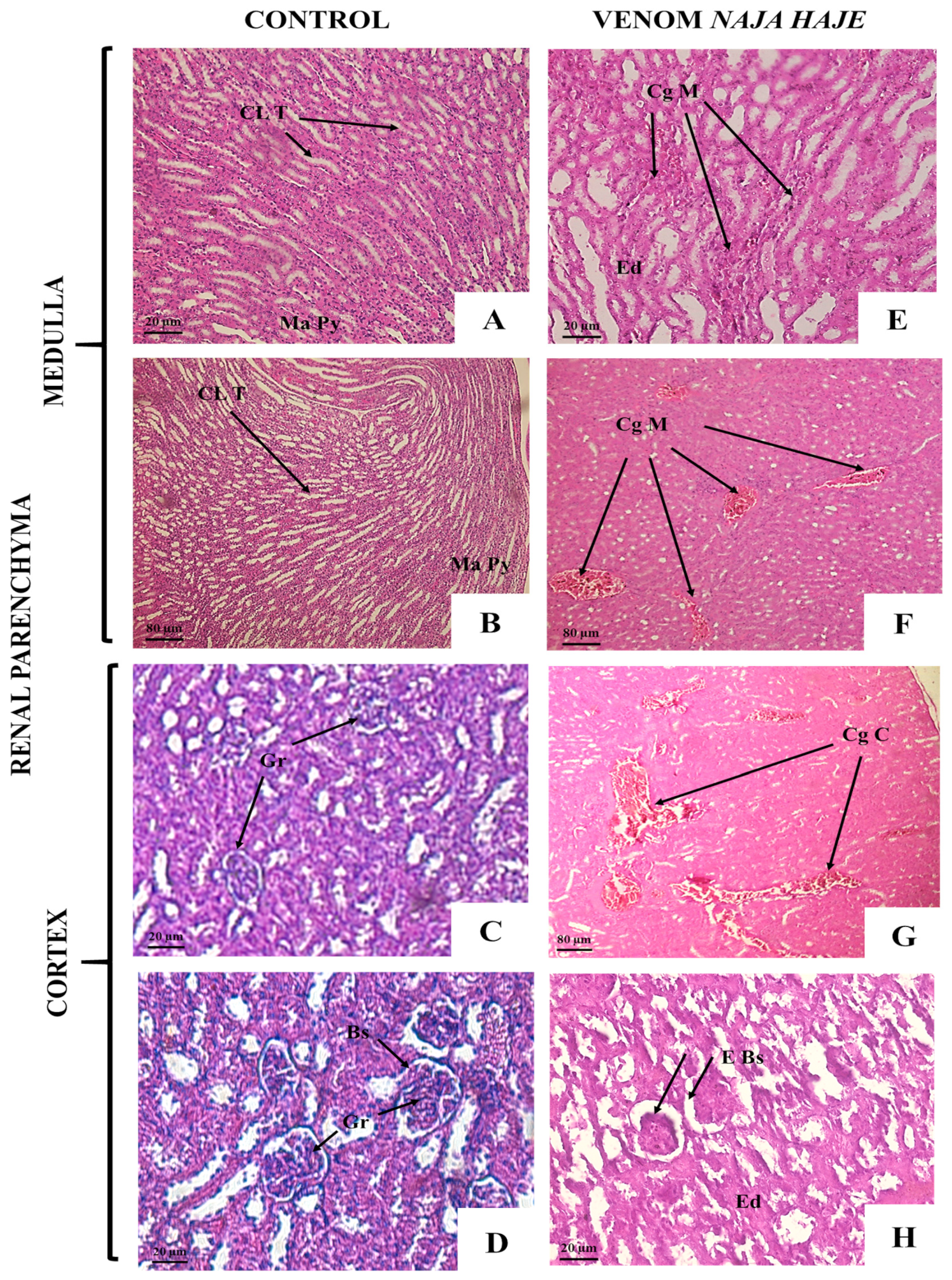
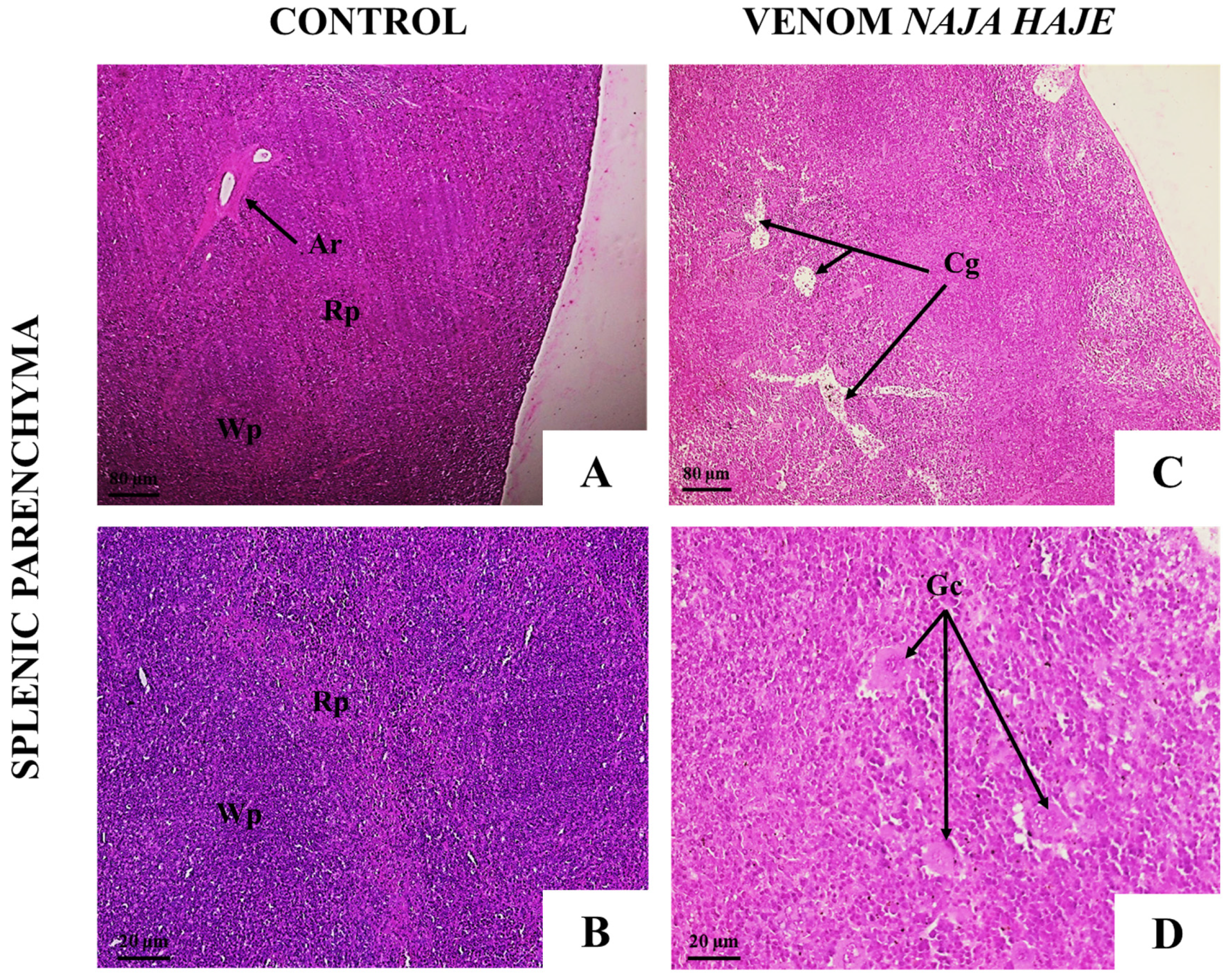
3.2.2. Naja haje Venom Induced Pathological Changes in Mouse Organs
3.3. Neutralization of Toxic Potency
3.3.1. Determination of the Median Effective Dose (DE50)
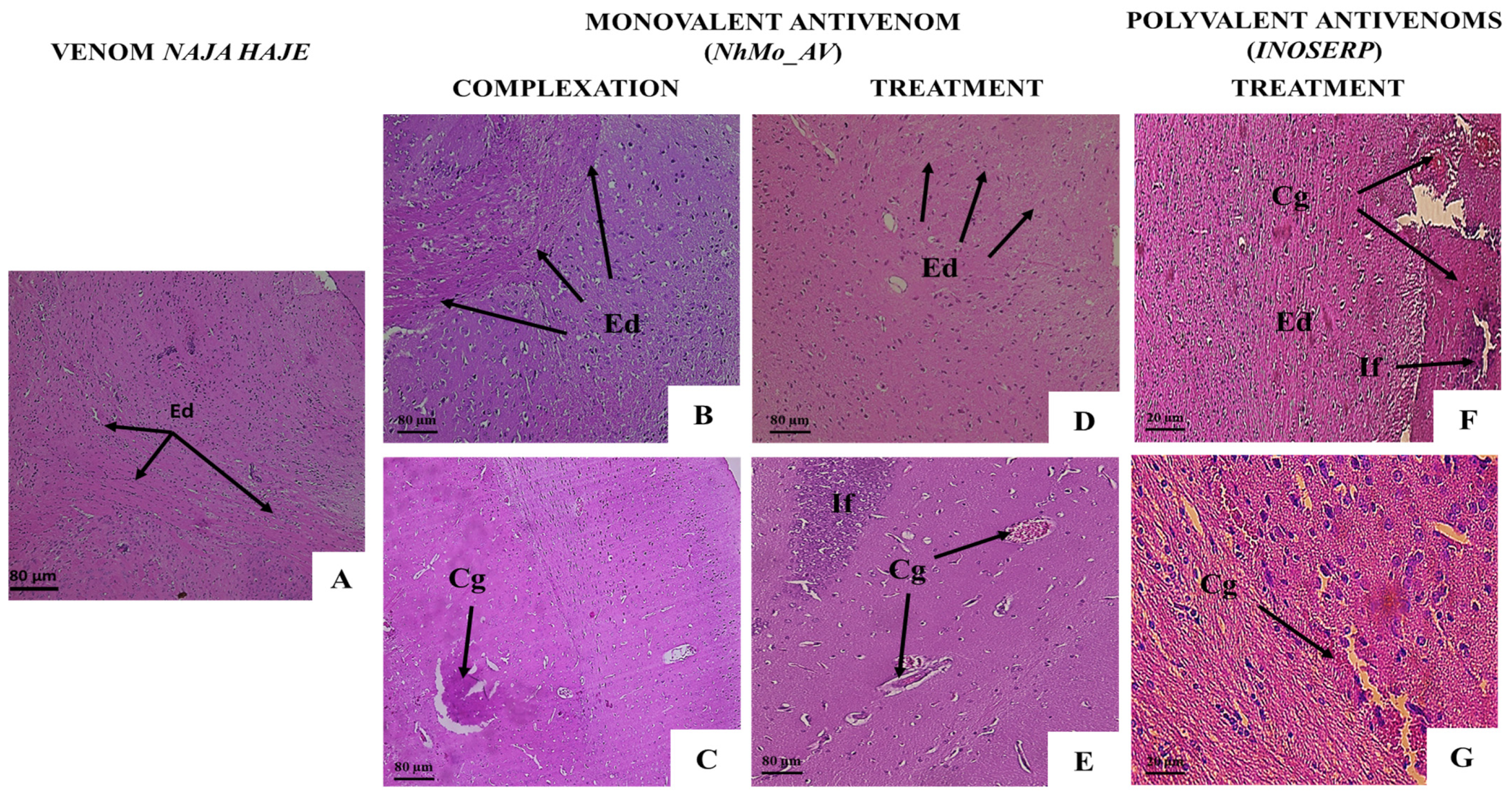
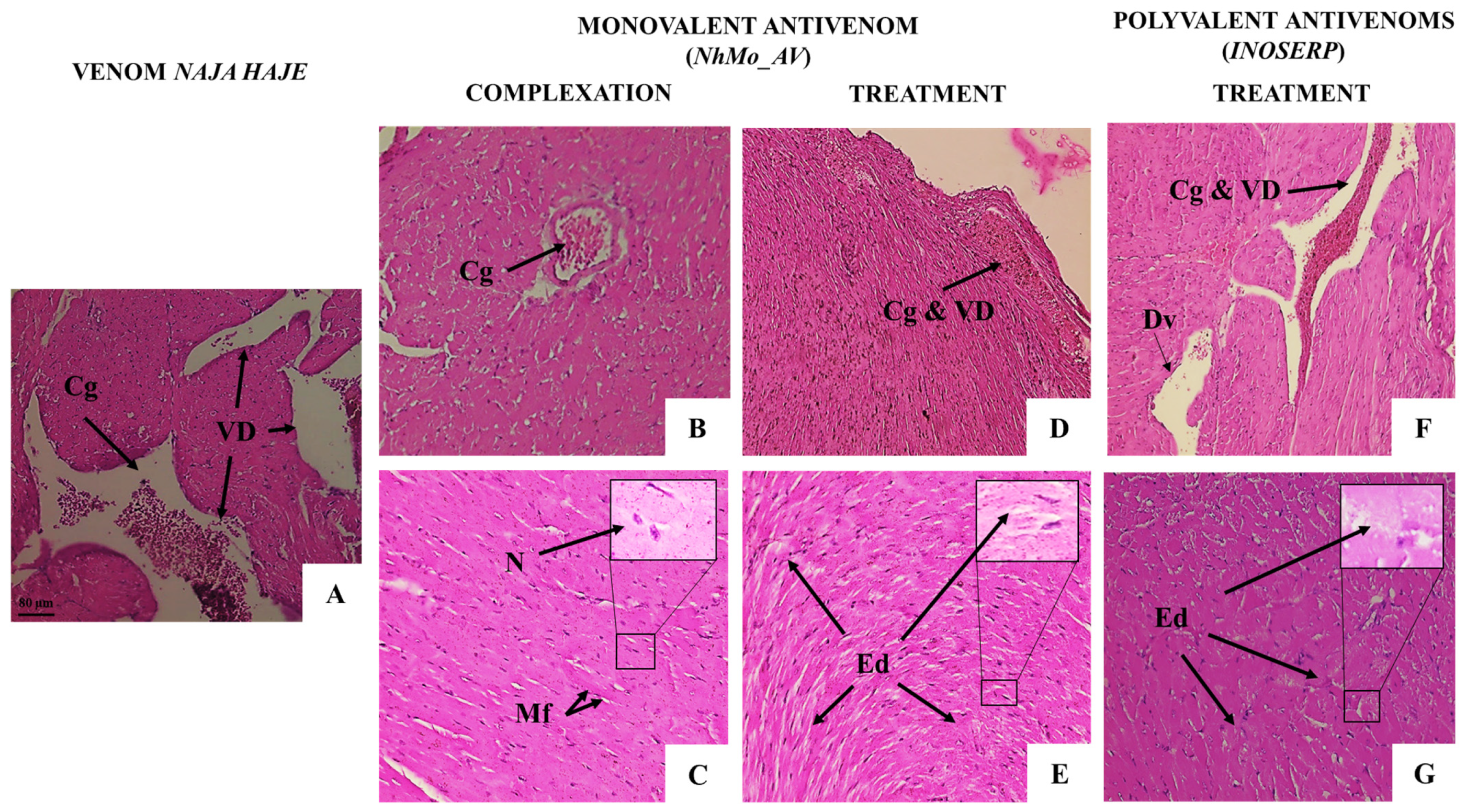
3.3.2. Neutralization of the Pathological Alterations by Monovalent and Polyvalent Antivenoms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Estimate of the burden of snakebites in sub-Saharan Africa: A meta-analytic approach. Toxicon 2011, 57, 586–599. [Google Scholar] [CrossRef]
- Benjamin, J.M.; Abo, B.N.; Brandehoff, N. Review Article: Snake Envenomation in Africa. Curr. Trop. Med. Rep. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Brito, J.C.; Fahd, S.; Geniez, P.; Martínez-Freiría, F.; Pleguezuelos, J.M.; Trape, J.F. Biogeography and conservation of viperids from North-West Africa: An application of ecological niche-based models and GIS. J. Arid. Environ. 2011, 75, 1029–1037. [Google Scholar] [CrossRef]
- Wilkins, D.; Burns, D.S.; Wilson, D.; Warrell, D.A.; Lamb, L.E.M. Snakebites in Africa and Europe: A military perspective and update for contemporary operations. J. R. Army Med. Corps 2018, 164, 370–379. [Google Scholar] [CrossRef]
- Modahl, C.M.; Roointan, A.; Rogers, J.; Currier, K.; Mackessy, S.P. Interspecific and intraspecific venom enzymatic variation among cobras (Naja sp. and Ophiophagus hannah). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 232, 108743. [Google Scholar] [CrossRef]
- Borja, M.; Lazcano, D.; Martínez-Romero, G.; Morlett, J.; Sánchez, E.; Cepeda-Nieto, A.C.; Garza-García, Y.; Zugasti-Cruz, A. Intra-specific Variation in the Protein Composition and Proteolytic Activity of Venom of Crotalus lepidus morulus from the Northeast of Mexico. Copeia 2013, 2013, 707–716. [Google Scholar] [CrossRef]
- Chippaux, J.P.; White, J.; Habib, A.G. African Snakes. In Critical Care Toxicology; Brent, J., Burkhart, K., Dargan, P., Hatten, B., Megarbane, B., Palmer, R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 2319–2342. Available online: http://link.springer.com/10.1007/978-3-319-17900-1_87 (accessed on 20 April 2023).
- Malih, I.; Ahmad Rusmili, M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of Moroccan cobra Naja haje legionis venom using tandem mass spectrometry. J. Proteom. 2014, 96, 240–252. [Google Scholar] [CrossRef]
- Karam, H.; Shaaban, E.; Fahmy, A.; Zaki, H.; Kenawy, S. Improvement of Naja haje snake antivenom production using gamma radiation and a biotechnological technique. Toxin Rev. 2021, 40, 48–58. [Google Scholar] [CrossRef]
- Berling, I.; Isbister, G.K. Hematologic Effects and Complications of Snake Envenoming. Transfus. Med. Rev. 2015, 29, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Križaj, I. Snake Venom Phospholipase A2 Toxins. In Handbook of Venoms and Toxins of Reptiles, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Ajisebiola, B.S.; Adeniji, O.B.; James, A.S.; Ajayi, B.O.; Adeyi, A.O. Naja nigricollis venom altered reproductive and neurological functions via modulation of pro-inflammatory cytokines and oxidative damage in male rats. Metab. Open 2022, 14, 100188. [Google Scholar] [CrossRef] [PubMed]
- Brunda, G.; Sashidhar, R.B.; Sarin, R.K. Use of egg yolk antibody (IgY) as an immunoanalytical tool in the detection of Indian cobra (Naja naja naja) venom in biological samples of forensic origin. Toxicon 2006, 48, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Wong, K.Y.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Elucidating the biogeographical variation of the venom of Naja naja (spectacled cobra) from Pakistan through a venom-decomplexing proteomic study. J. Proteom. 2018, 175, 156–173. [Google Scholar] [CrossRef]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef]
- Palasuberniam, P.; Chan, Y.W.; Tan, K.Y.; Tan, C.H. Snake Venom Proteomics of Samar Cobra (Naja samarensis) from the Southern Philippines: Short Alpha-Neurotoxins as the Dominant Lethal Component Weakly Cross-Neutralized by the Philippine Cobra Antivenom. Front. Pharmacol. 2021, 12, 727756. [Google Scholar] [CrossRef]
- Hansson, E.; Sasa, M.; Mattisson, K.; Robles, A.; Gutiérrez, J.M. Using Geographical Information Systems to Identify Populations in Need of Improved Accessibility to Antivenom Treatment for Snakebite Envenoming in Costa Rica. PLoS Negl. Trop. Dis. 2013, 7, e2009. [Google Scholar] [CrossRef]
- WHO Issues New Recommendation on Antivenom for Snakebites. Available online: https://www.who.int/news/item/19-08-2018-who-issues-new-recommendation-on-antivenom-for-snakebites (accessed on 21 April 2023).
- Becker, E.L. Antigen-antibody reactions in gels. Fed. Proc. 1953, 12, 717–722. [Google Scholar]
- Chavez-Olortegui, C.; Lopes, C.S.; Cordeiro, F.D.; Granier, C.; Diniz, C.R. An enzyme linked immunosorbent assay (ELISA) that discriminates between Bothrops atrox and Lachesis muta muta venoms. Toxicon 1993, 31, 417–426. [Google Scholar] [CrossRef]
- Khoobdel, M.; Fasaei, B.N.; Salehi, T.Z.; Khosravi, M.; Taheri, M.; Koochakzadeh, A.; Masihipour, B.; Motedayen, M.; Akbari, S. The production of monovalent and anti-idiotype antivenom against Mesobuthus eupeus (Scorpionida: Buthidae) venom in rabbits. Toxicon 2013, 76, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; El Jaoudi, R.; Ghalim, N.; Chgoury, F.; Bouhaouala, B.; El Mdaghri, N.; Sabatier, J.-M. Evaluation of the Lethal Potency of Scorpion and Snake Venoms and Comparison between Intraperitoneal and Intravenous Injection Routes. Toxins 2014, 6, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, R.T. Theory and Practice of Histological Techniques. JAMA 1983, 250, 1909. [Google Scholar] [CrossRef]
- Ross, M.H.; Pawlina, W. Histology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006; 934p. [Google Scholar]
- Trevors, J.T. A BASIC Program for Estimating LDso Values Using the IBM-PC. Bull. Environ. Contam. Toxicol. 1986, 37, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Tan, C.H.; Sim, S.M.; Fung, S.Y.; Tan, N.H. Geographical venom variations of the Southeast Asian monocled cobra (Naja kaouthia): Venom-induced neuromuscular depression and antivenom neutralization. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 185–186, 77–86. [Google Scholar] [CrossRef]
- World Health Organization. WHO Expert Committee on Biological Standardization (2016: Geneva S. WHO Expert Committee on Biological Standardization, Sixty-Seventh Report; WHO Technical Report Series, 1004; World Health Organization: Geneva, Switzerland, 2017; 616p, Available online: https://apps.who.int/iris/handle/10665/255657 (accessed on 20 April 2023).
- Mukherjee, A.K.; Maity, C.R. Biochemical composition, lethality and pathophysiology of venom from two cobras—Naja naja and N. kaouthia. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 131, 125–132. [Google Scholar] [CrossRef]
- Lafnoune, A.; Lee, S.-Y.; Heo, J.-Y.; Gourja, I.; Darkaoui, B.; Abdelkafi-Koubaa, Z.; Chgoury, F.; Daoudi, K.; Chakir, S.; Cadi, R.; et al. Anti-Cancer Effect of Moroccan Cobra Naja haje Venom and Its Fractions against Hepatocellular Carcinoma in 3D Cell Culture. Toxins 2021, 13, 402. [Google Scholar] [CrossRef]
- Demirel, M.A.; Alcigir, M.E.; Ozkan, O.; Turkmen, M.B. The effects of antivenom administrations on the brain tissue of experimentally envenomed pregnant rats and their pups with Androctonus crassicauda scorpion venom during organogenesis period. Toxicon 2021, 200, 13–18. [Google Scholar] [CrossRef]
- Naoual, O.; Rachid, E.; Sebastien, L.; Salma, C.; Chafi, Q.F.; Abdelaziz, H.; Georges, M. Snake bites in morocco: Progress and challenges. Adv. Toxicol. Toxic. Eff. 2019, 3, 9–14. [Google Scholar] [CrossRef]
- Dissanayake, D.S.B.; Thewarage, L.D.; Waduge, R.N.; Ranasinghe, J.G.S.; Kularatne, S.A.M.; Rajapakse, R.P.V.J. The Venom of Spectacled Cobra (Elapidae: Naja naja): In Vitro Study from Distinct Geographical Origins in Sri Lanka. J. Toxicol. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Al-Mamun, M.A.; Hakim, M.A.; Zaman, M.K.; Hoque, K.M.F.; Ferdousi, Z.; Reza, M.A. Histopathological Alterations Induced by Naja naja Crude Venom on Renal, Pulmonary and Intestinal Tissues of Mice Model. Biotechnol. J. Int. 2015, 6, 119–125. [Google Scholar] [CrossRef]
- Cher, C.D.N.; Armugam, A.; Zhu, Y.Z.; Jeyaseelan, K. Molecular basis of cardiotoxicity upon cobra envenomation. Cell. Mol. Life Sci. 2005, 62, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Liu, J.-H.; Lee, S.-C.; Hsiao, C.-D.; Wu, W.-G. Glycosphingolipid-facilitated Membrane Insertion and Internalization of Cobra Cardiotoxin. J. Biol. Chem. 2006, 281, 656–667. [Google Scholar] [CrossRef]
- Hung, D.Z.; Liau, M.Y.; Lin-Shiau, S.Y. The clinical significance of venom detection in patients of cobra snakebite. Toxicon 2003, 41, 409–415. [Google Scholar] [CrossRef]
- Adzu, B.; Abubakar, M.S.; Izebe, K.S.; Akumka, D.D.; Gamaniel, K.S. Effect of Annona senegalensis rootbark extracts on Naja nigricotlis nigricotlis venom in rats. J. Ethnopharmacol. 2005, 96, 507–513. [Google Scholar] [CrossRef]
- Al-Ani, I.; Ismail, S.; Maung, K.; Oothuman, P.; Al-Mahmood, S. Histological study on the protective effects of Tamarind seed extract on Cobra venom in mice. Asian J. Pharm. Clin. Res. 2017, 10, 301. [Google Scholar] [CrossRef]
- Al-Sadoon, M.K.; Orabi, G.M.; Badr, G. Toxic Effects of Crude Venom of a Desert Cobra, Walterinnesia aegyptia, on Liver, Abdominal Muscles and Brain of Male Albino Rats. Pak. J. Zool. 2013, 45, 1359–1366. [Google Scholar]
- Tohamy, A.A.; Mohamed, A.F.; Abdel Moneim, A.E.; Diab, M.S.M. Biological effects of Naja haje crude venom on the hepatic and renal tissues of mice. J. King Saud Univ. Sci. 2014, 26, 205–212. [Google Scholar] [CrossRef]
- Ding, Z.-H.; Xu, L.-M.; Wang, S.-Z.; Kou, J.-Q.; Xu, Y.-L.; Chen, C.-X.; Yu, H.-P.; Qin, Z.-H.; Xie, Y. Ameliorating Adriamycin-Induced Chronic Kidney Disease in Rats by Orally Administrated Cardiotoxin from Naja naja atra Venom. Evid. Based Complement. Altern. Med. 2014, 2014, 621756. [Google Scholar] [CrossRef]
- Abdel-Ghani, L.; El-Asmer, M.; Abuzinadah, O.; Abbas, A.; Rahmy, T. Histological and immunohistochemical studies on the nephrotoxic effects of naja nigricollis snake venom. Egypt. J. Nat. Toxins 2010, 7, 29–52. [Google Scholar]
- Rivel, M.; Solano, D.; Herrera, M.; Vargas, M.; Villalta, M.; Segura, Á.; Arias, A.S.; León, G.; Gutiérrez, J.M. Pathogenesis of dermonecrosis induced by venom of the spitting cobra, Naja nigricollis: An experimental study in mice. Toxicon 2016, 119, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Senji Laxme, R.R.; Khochare, S.; De Souza, H.F.; Ahuja, B.; Suranse, V.; Martin, G.; Whitaker, R.; Sunagar, K. Beyond the ‘big four’: Venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl. Trop. Dis. 2019, 13, e0007899. [Google Scholar] [CrossRef]
- Sandesha, V.D.; Darshan, B.; Tejas, C.; Girish, K.S.; Kempaiah, K. A comparative cross-reactivity and paraspecific neutralization study on Hypnale hypnale, Echis carinatus, and Daboia russelii monovalent and therapeutic polyvalent anti-venoms. PLoS Negl. Trop. Dis. 2022, 16, e0010292. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Wong, K.Y.; Tan, N.H.; Tan, C.H. Quantitative proteomics of Naja annulifera (sub-Saharan snouted cobra) venom and neutralization activities of two antivenoms in Africa. Int. J. Biol. Macromol. 2020, 158, 605–616. [Google Scholar] [CrossRef]
- Manson, E.Z.; Kyama, M.C.; Gikunju, J.K.; Kimani, J.; Kimotho, J.H. Evaluation of lethality and cytotoxic effects induced by Naja ashei (large brown spitting cobra) venom and the envenomation-neutralizing efficacy of selected commercial antivenoms in Kenya. Toxicon X 2022, 14, 100125. [Google Scholar] [CrossRef]
- Shashidharamurthy, R.; Kemparaju, K. Region-specific neutralization of Indian cobra (Naja naja) venom by polyclonal antibody raised against the eastern regional venom: A comparative study of the venoms from three different geographical distributions. Int. Immunopharmacol. 2007, 7, 61–69. [Google Scholar] [CrossRef]
- Shaban, E.A.; Hafez, M.N. Ability of Gamma-Irradiated Polyvalent Antivenin to Neutralize the Toxicity of the Egyptian Cobra (Naja haje) Venom. Egypt J. Hosp. Med. 2003, 13, 135–152. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Naja haje Venom | |||
|---|---|---|---|
| Injection Route | µg/Mouse | µg/g | Ratio (IV/IP) |
| Intravenous IV | 18.6 (15.2–20.7) | 0.93 (0.76–1.035) | 1.16 |
| Intraperitoneal IP | 16.05 (13.16–21.49) | 0.80 (0.65–1.074) | |
| Microscopic Observation | Groups | ||||
|---|---|---|---|---|---|
| Control | Naja haje | Monovalent Moroccan Naja haje Antivenom (NhMo_AV) | Commercialzed Antivenom (Inoserp-MENA) | ||
| Complexation | Treatment | Treatment | |||
| Brain Tissue | |||||
| Cerebral edema | − | ++ | + | + | ++ |
| Inflammatory cell infiltrate | − | ++ | − | − | + |
| Vascular congestion | − | +++ | + | + | ++ |
| Cardiac Tissue | |||||
| Loss of striations in the myocardial fibers | − | ++ | − | − | − |
| Interstitial edema | − | +++ | − | + | ++ |
| Myocardial cells Hypertrophy | − | ++ | − | + | + |
| Vascular congestion and dilation | − | +++ | + | + | ++ |
| Lung Tissue | |||||
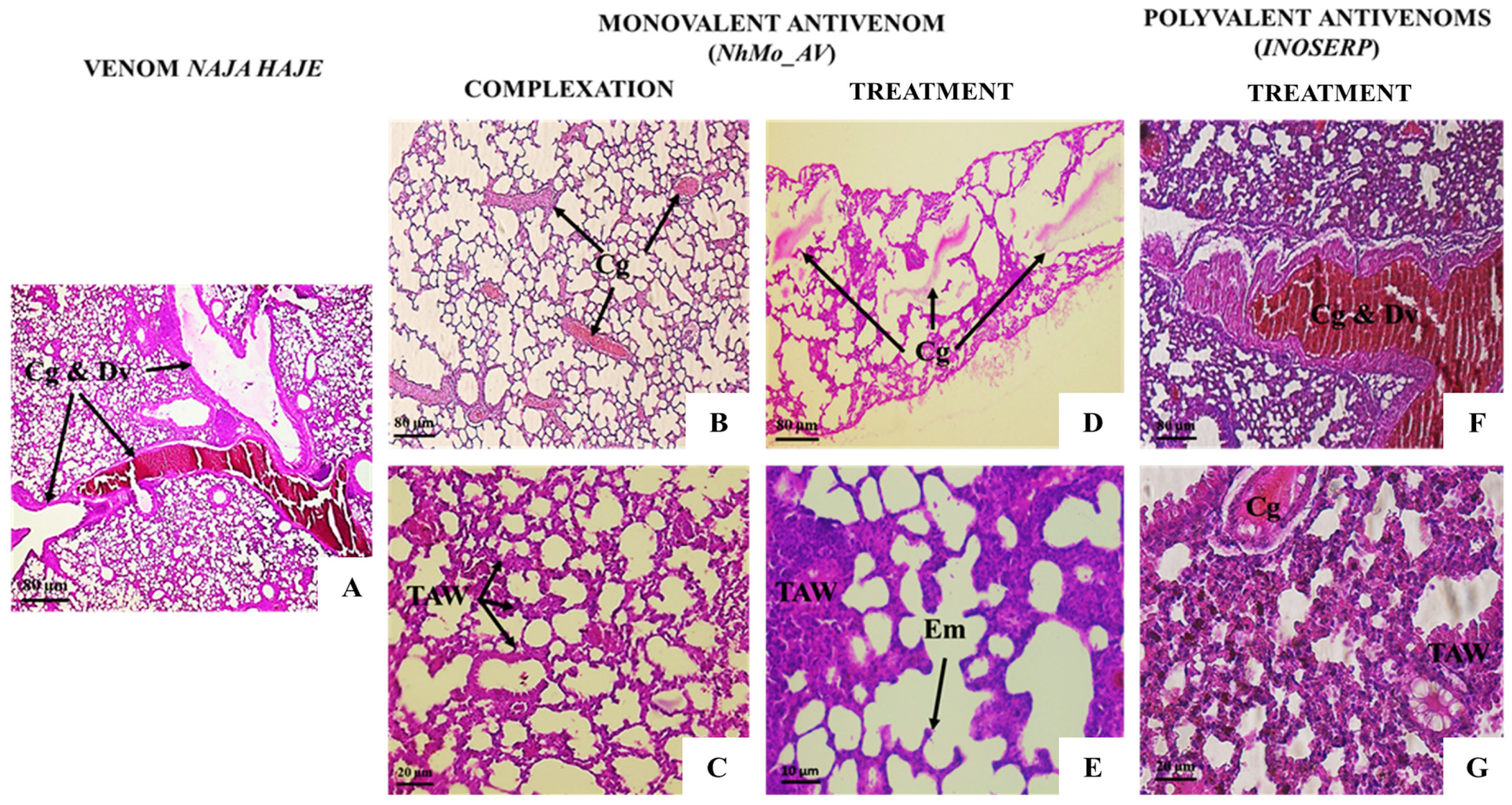
| Thickening of the alveolar walls | − | +++ | + | + | ++ |
| Pulmonary edema | − | +++ | − | + | ++ |
| Emphysema | − | + | + | + | ++ |
| Polymorphic inflammatory cells | − | ++ | − | + | + |
| Vascular congestion and dilation | − | +++ | + | ++ | ++ |
| Hepatic Tissue | |||||
| Ballooning of hepatocytes | − | +++ | + | + | ++ |
| Cytoplasmic vacuolation of hepatocyte | − | ++ | − | + | ++ |
| Dilation of the sinusoidal vessels | − | ++ | + | + | + |
| Congestion and dilation of central venous | − | +++ | + | + | ++ |
| Polynuclear inflammatory cells | − | ++ | − | + | ++ |
| Kidney Tissue | |||||
| Dilation of Bowman’s space | − | ++ | + | + | ++ |
| Cortical and medullary edema | − | +++ | − | − | + |
| Congestion and dilatation of distal and proximal tubules | − | +++ | − | ++ | ++ |
| Cell hypertrophy | − | ++ | + | + | + |
| Infiltration of polymorphic inflammatory cells | − | + | + | + | ++ |
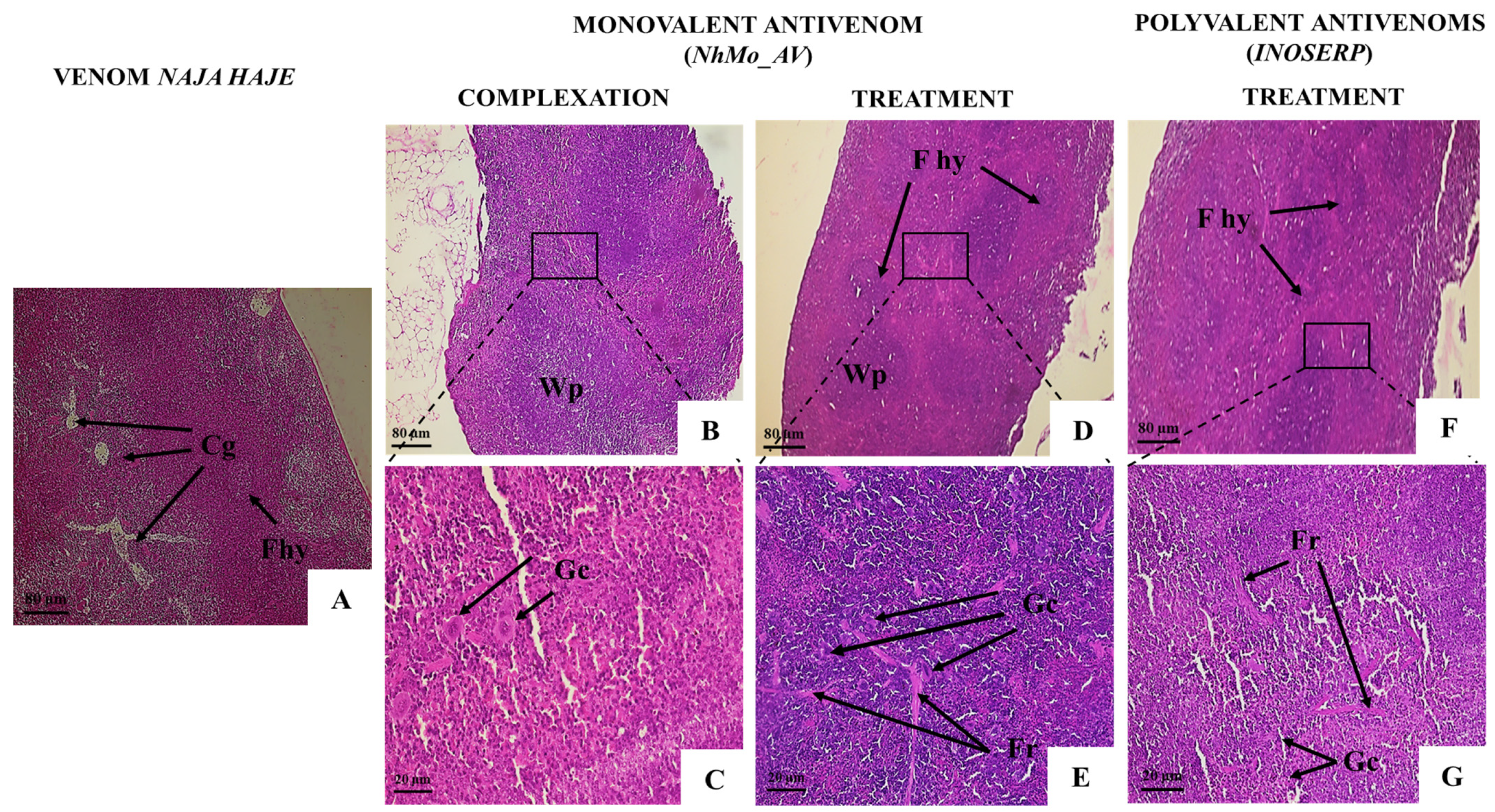
| Splenic Tissue | |||||
| Follicular hyperplasia | − | ++ | + | + | + |
| Presence of giant cells | − | ++ | + | ++ | ++ |
| Congestion | − | ++ | − | − | − |
| Infiltration of inflammatory cells | − | + | − | + | + |
| Naja haje Venom | Antivenom Neutralization | |||||||
|---|---|---|---|---|---|---|---|---|
| i.p LD50 a (μg/g) | Challenge Dose | Antivenom | ED50 b (μL) | ER50 c (mg/mL) | P d (mg/mL) | Antivenom Concentration (mg/mL) | Normalized Potency e (mg/g) | |
| Naja haje | 0.80 | 5 LD50 | NhMo_AV | 67.2 (50.45–70.8) | 1.19 (1.60–1.13) | 0.95 | 74.5 ± 4.70 | 12.84 |
| 3 LD50 | Inoserp-MENA | 100 (112.5–122.5) | 0.48 (0.428–0.39) | 0.32 | 20 ± 3.45 | 16.05 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khourcha, S.; Hilal, I.; Elbejjaj, I.; Karkouri, M.; Safi, A.; Hmyene, A.; Oukkache, N. Assessing the Efficacy of Monovalent and Commercialized Antivenoms for Neutralizing Moroccan Cobra Naja haje Venom: A Comparative Study. Trop. Med. Infect. Dis. 2023, 8, 304. https://doi.org/10.3390/tropicalmed8060304
Khourcha S, Hilal I, Elbejjaj I, Karkouri M, Safi A, Hmyene A, Oukkache N. Assessing the Efficacy of Monovalent and Commercialized Antivenoms for Neutralizing Moroccan Cobra Naja haje Venom: A Comparative Study. Tropical Medicine and Infectious Disease. 2023; 8(6):304. https://doi.org/10.3390/tropicalmed8060304
Chicago/Turabian StyleKhourcha, Soukaina, Ines Hilal, Iatimad Elbejjaj, Mehdi Karkouri, Amal Safi, Abdelaziz Hmyene, and Naoual Oukkache. 2023. "Assessing the Efficacy of Monovalent and Commercialized Antivenoms for Neutralizing Moroccan Cobra Naja haje Venom: A Comparative Study" Tropical Medicine and Infectious Disease 8, no. 6: 304. https://doi.org/10.3390/tropicalmed8060304
APA StyleKhourcha, S., Hilal, I., Elbejjaj, I., Karkouri, M., Safi, A., Hmyene, A., & Oukkache, N. (2023). Assessing the Efficacy of Monovalent and Commercialized Antivenoms for Neutralizing Moroccan Cobra Naja haje Venom: A Comparative Study. Tropical Medicine and Infectious Disease, 8(6), 304. https://doi.org/10.3390/tropicalmed8060304