
Effect of Enzymatic Hydrolysis on Solubility and Emulsifying Properties of Lupin Proteins (Lupinus luteus)


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Lupin Protein Isolate (LPI)
2.3. Chemical Analysis of LPI
2.4. Enzymatic Hydrolysis of LPI
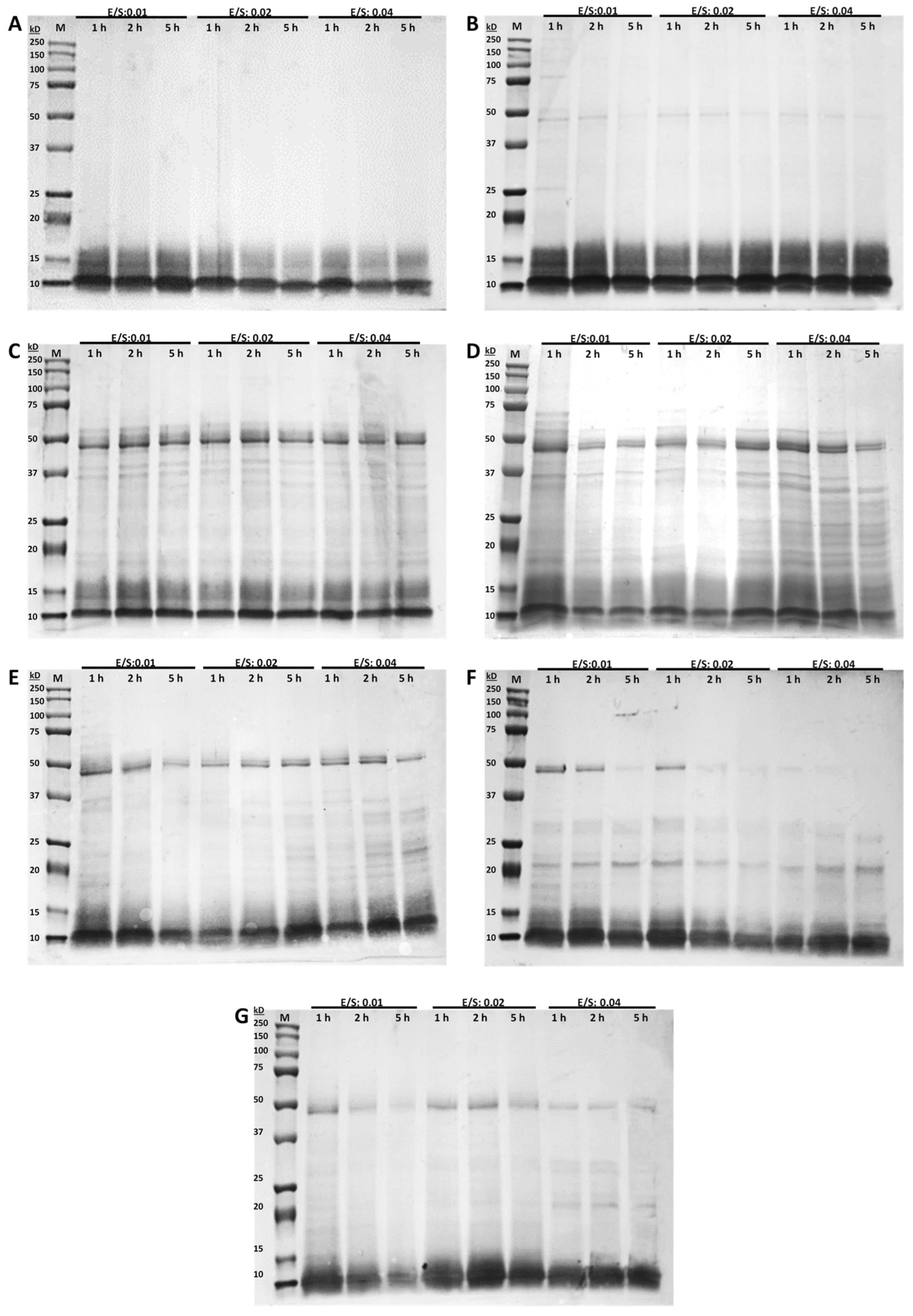
2.5. SDS-PAGE Analysis
2.6. Degree of Hydrolysis (DH)
2.7. Protein Solubility
2.8. Emulsifying Capacity (EC) and Emulsion Stability (ES)
2.9. Statistical Analysis
3. Results and Discussions
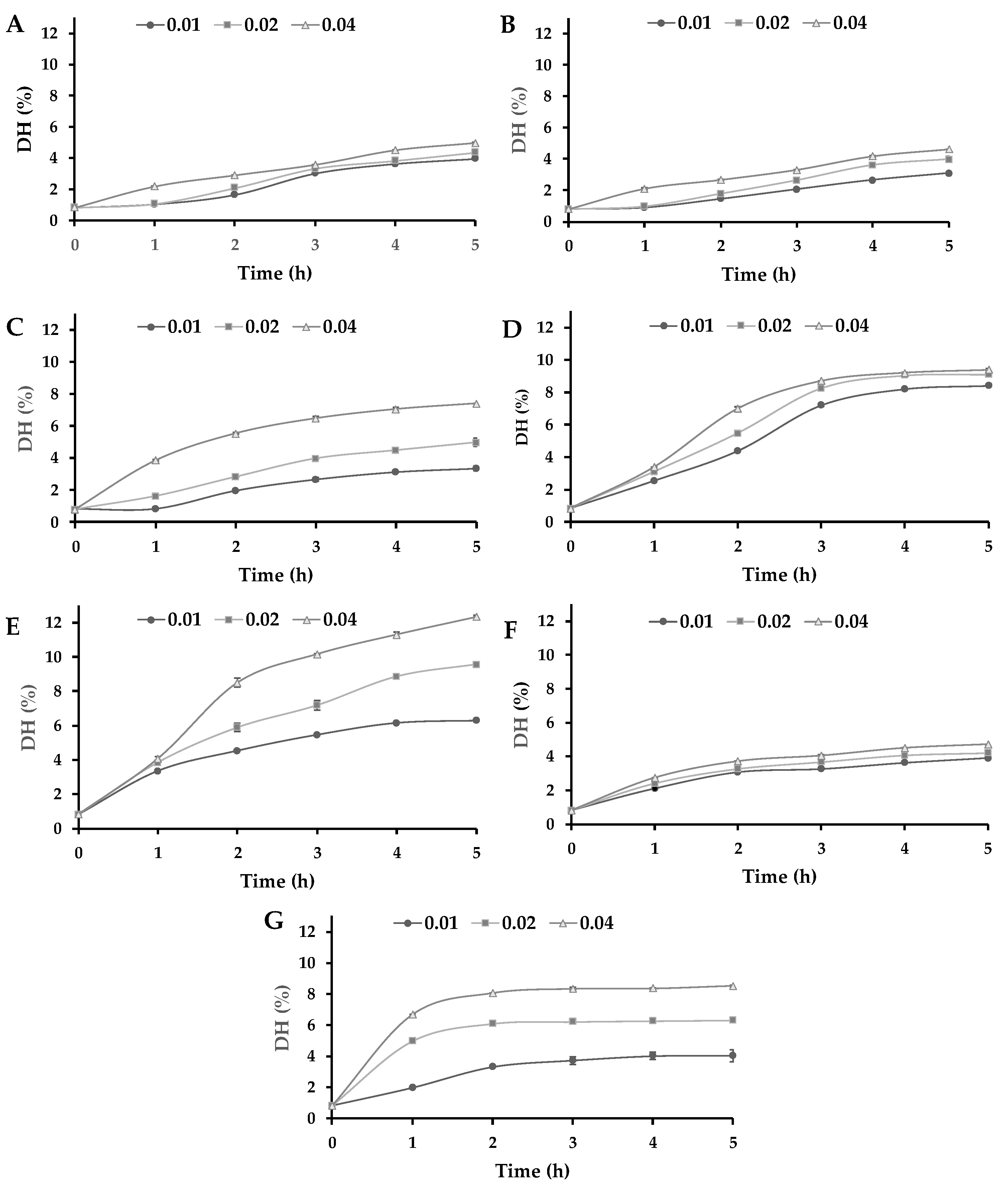
3.1. Enzymatic Hydrolysis of Lupin Proteins
3.2. Molecular Weight Distributions (SDS-PAGE)
3.3. Functional Properties
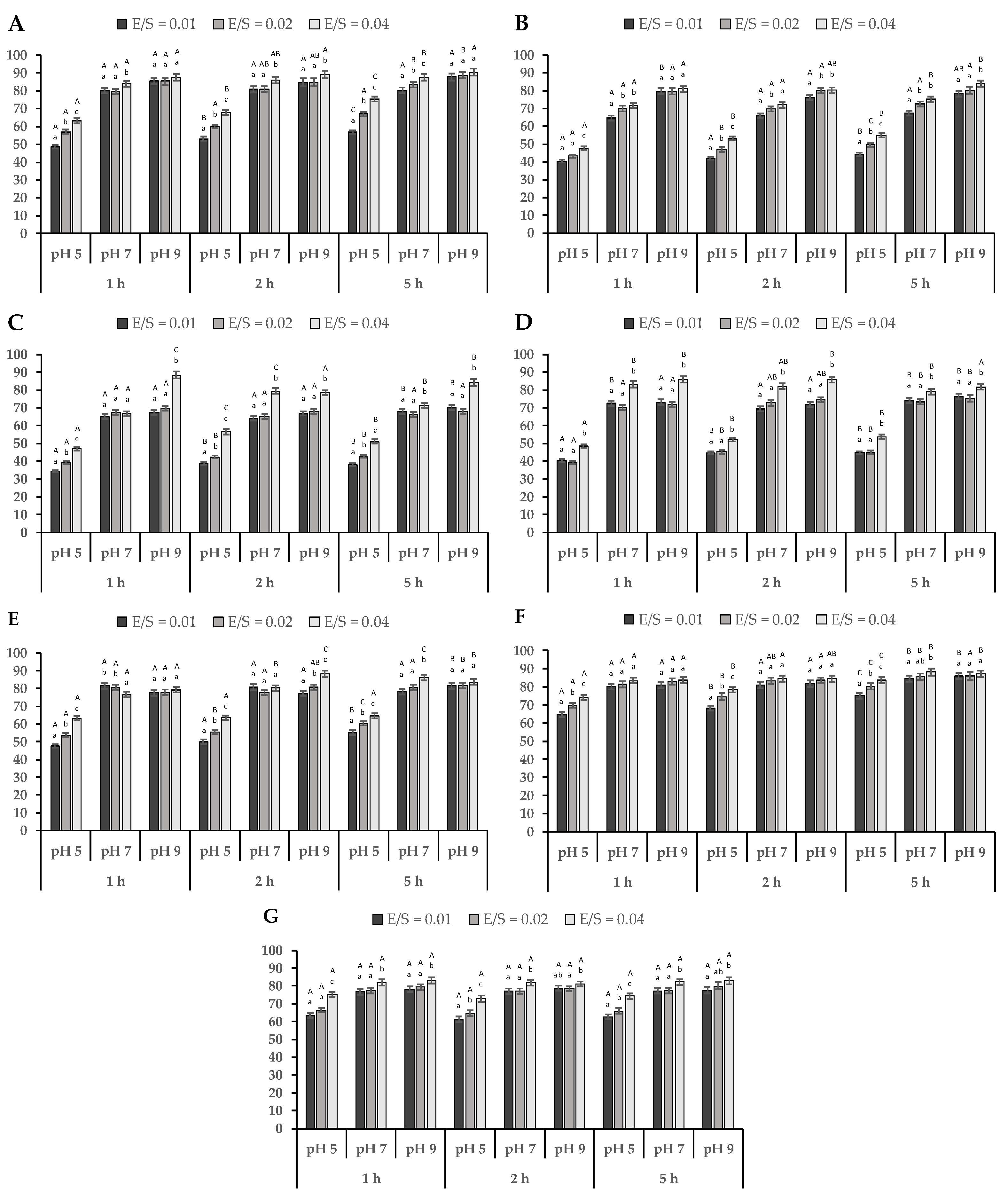
3.3.1. Protein Solubility
3.3.2. Emulsion Capacity (EC) and Emulsion Stability (ES) of Hydrolyzed LPI
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikbakht Nasrabadi, M.; Sedaghat Doost, A.; Mezzenga, R. Modification Approaches of Plant-Based Proteins to Improve Their Techno-Functionality and Use in Food Products. Food Hydrocoll. 2021, 118, 106789. [Google Scholar] [CrossRef]
- McClements, D.J.; Lu, J.; Grossmann, L. Proposed Methods for Testing and Comparing the Emulsifying Properties of Proteins from Animal, Plant, and Alternative Sources. Colloids Interfaces 2022, 6, 19. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Opazo-Navarrete, M.; Wandersleben, T.; Soto-Añual, M.; Barahona, T.; Bustamante, M. Chemical and Nutritional Evaluation of Protein-Rich Ingredients Obtained through a Technological Process from Yellow Lupin Seeds (Lupinus luteus). Plant Foods Hum. Nutr. 2019, 74, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht Nasrabadi, M.; Goli, S.A.H.; Sedaghat Doost, A.; Dewettinck, K.; Van der Meeren, P. Bioparticles of Flaxseed Protein and Mucilage Enhance the Physical and Oxidative Stability of Flaxseed Oil Emulsions as a Potential Natural Alternative for Synthetic Surfactants. Colloids Surf. B Biointerfaces 2019, 184, 110489. [Google Scholar] [CrossRef]
- Hu, G.J.; Zhao, Y.; Gao, Q.; Wang, X.W.; Zhang, J.W.; Peng, X.; Tanokura, M.; Xue, Y.L. Functional Properties of Chinese Yam (Dioscorea opposita Thunb. Cv. Baiyu) Soluble Protein. J. Food Sci. Technol. 2018, 55, 381–388. [Google Scholar] [CrossRef]
- Rivera del Rio, A.; Opazo-Navarrete, M.; Cepero-Betancourt, Y.; Tabilo-Munizaga, G.; Boom, R.M.; Janssen, A.E.M. Heat-Induced Changes in Microstructure of Spray-Dried Plant Protein Isolates and Its Implications on in Vitro Gastric Digestion. LWT Food Sci. Technol. 2020, 118, 108795. [Google Scholar] [CrossRef]
- Avramenko, N.A.; Low, N.H.; Nickerson, M.T. The Effects of Limited Enzymatic Hydrolysis on the Physicochemical and Emulsifying Properties of a Lentil Protein Isolate. Food Res. Int. 2013, 51, 162–169. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Mosi-Roa, Y.; Opazo-Navarrete, M.; Bustamante, M.; Garrido-Miranda, K. Comparative Study of Food-Grade Pickering Stabilizers Obtained from Agri-Food Byproducts: Chemical Characterization and Emulsifying Capacity. Foods 2022, 11, 2514. [Google Scholar] [CrossRef]
- McClements, D.J.; Gumus, C.E. Natural Emulsifiers—Biosurfactants, Phospholipids, Biopolymers, and Colloidal Particles: Molecular and Physicochemical Basis of Functional Performance. Adv. Colloid Interface Sci. 2016, 234, 3–26. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Campanella, O.H. Limited Enzymatic Hydrolysis Induced Pea Protein Gelation at Low Protein Concentration with Less Heat Requirement. Food Hydrocoll. 2022, 128, 107547. [Google Scholar] [CrossRef]
- Liu, N.; Lin, P.; Zhang, K.; Yao, X.; Li, D.; Yang, L.; Zhao, M. Combined Effects of Limited Enzymatic Hydrolysis and High Hydrostatic Pressure on the Structural and Emulsifying Properties of Rice Proteins. Innov. Food Sci. Emerg. Technol. 2022, 77, 102975. [Google Scholar] [CrossRef]
- Gomes, M.H.G.; Kurozawa, L.E. Improvement of the Functional and Antioxidant Properties of Rice Protein by Enzymatic Hydrolysis for the Microencapsulation of Linseed Oil. J. Food Eng. 2020, 267, 109761. [Google Scholar] [CrossRef]
- Al-Ruwaih, N.; Ahmed, J.; Mulla, M.F.; Arfat, Y.A. High-Pressure Assisted Enzymatic Proteolysis of Kidney Beans Protein Isolates and Characterization of Hydrolysates by Functional, Structural, Rheological and Antioxidant Properties. LWT Food Sci. Technol. 2019, 100, 231–236. [Google Scholar] [CrossRef]
- Eckert, E.; Han, J.; Swallow, K.; Tian, Z.; Jarpa-Parra, M.; Chen, L. Effects of Enzymatic Hydrolysis and Ultrafiltration on Physicochemical and Functional Properties of Faba Bean Protein. Cereal Chem. 2019, 96, 725–741. [Google Scholar] [CrossRef]
- Brückner-Gühmann, M.; Heiden-Hecht, T.; Sözer, N.; Drusch, S. Foaming Characteristics of Oat Protein and Modification by Partial Hydrolysis. Eur. Food Res. Technol. 2018, 244, 2095–2106. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Yu, L.; Wu, K.; Zhao, M. Emulsification Performance and Interfacial Properties of Enzymically Hydrolyzed Peanut Protein Isolate Pretreated by Extrusion Cooking. Food Hydrocoll. 2018, 77, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chen, J.; Yu, L.; Wu, K. Improved Emulsifying Capabilities of Hydrolysates of Soy Protein Isolate Pretreated with High Pressure Microfluidization. LWT Food Sci. Technol. 2016, 69, 1–8. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, K.; Sontheimer, K.; Hickisch, A.; Wani, A.A.; Eisner, P.; Schweiggert-Weisz, U. Enzymatic Hydrolysis of Lupin Protein Isolates—Changes in the Molecular Weight Distribution, Technofunctional Characteristics, and Sensory Attributes. Food Sci. Nutr. 2019, 7, 2747–2759. [Google Scholar] [CrossRef] [Green Version]
- Muhoza, B.; Qi, B.; Harindintwali, J.D.; Farag Koko, M.Y.; Zhang, S.; Li, Y. Combined Plant Protein Modification and Complex Coacervation as a Sustainable Strategy to Produce Coacervates Encapsulating Bioactives. Food Hydrocoll. 2022, 124, 107239. [Google Scholar] [CrossRef]
- García Arteaga, V.; Apéstegui Guardia, M.; Muranyi, I.; Eisner, P.; Schweiggert-Weisz, U. Effect of Enzymatic Hydrolysis on Molecular Weight Distribution, Techno-Functional Properties and Sensory Perception of Pea Protein Isolates. Innov. Food Sci. Emerg. Technol. 2020, 65, 102449. [Google Scholar] [CrossRef]
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional Enzymes in Life and Disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Yi, L.; Marek, P.; Iverson, B.L. Commercial Proteases: Present and Future. FEBS Lett. 2013, 587, 1155–1163. [Google Scholar] [CrossRef] [Green Version]
- Padial-Domínguez, M.; Espejo-Carpio, F.J.; Pérez-Gálvez, R.; Guadix, A.; Guadix, E.M. Optimization of the Emulsifying Properties of Food Protein Hydrolysates for the Production of Fish Oil-in-Water Emulsions. Foods 2020, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhong, J.; Chen, J.; Liu, C.; Luo, L.; Luo, S.; Wu, L.; McClements, D.J. Effectiveness of Partially Hydrolyzed Rice Glutelin as a Food Emulsifier: Comparison to Whey Protein. Food Chem. 2016, 213, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickisch, A.; Bindl, K.; Vogel, R.F.; Toelstede, S. Thermal Treatment of Lupin-Based Milk Alternatives–Impact on Lupin Proteins and the Network of Respective Lupin-Based Yogurt Alternatives. Food Res. Int. 2016, 89, 850–859. [Google Scholar] [CrossRef]
- Cabello-Hurtado, F.; Keller, J.; Ley, J.; Sanchez-Lucas, R.; Jorrín-Novo, J.V.; Aïnouche, A. Proteomics for Exploiting Diversity of Lupin Seed Storage Proteins and Their Use as Nutraceuticals for Health and Welfare. J. Proteom. 2016, 143, 57–68. [Google Scholar] [CrossRef]
- Starkute, V.; Bartkiene, E.; Bartkevics, V.; Rusko, J.; Zadeike, D.; Juodeikiene, G. Amino Acids Profile and Antioxidant Activity of Different Lupinus angustifolius Seeds after Solid State and Submerged Fermentations. J. Food Sci. Technol. 2016, 53, 4141–4148. [Google Scholar] [CrossRef] [Green Version]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Yellow Lupin (Lupinus luteus L.) Polysaccharides: Antioxidant, Immunomodulatory and Prebiotic Activities and Their Structural Characterisation. Food Chem. 2018, 267, 319–328. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Piornos, J.A.; Wandersleben, T.; Ogura, T.; Hernández, X.; Rubilar, M. Emulsifying and Foaming Properties of Different Protein Fractions Obtained from a Novel Lupin Variety AluProt-CGNA® (Lupinus luteus). J. Food Sci. 2016, 81, C1699–C1706. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Opazo-Navarrete, M.; Soto-Añual, M.; Leal-Calderón, F.; Bustamante, M. Food-Grade Pickering Emulsion as a Novel Astaxanthin Encapsulation System for Making Powder-Based Products: Evaluation of Astaxanthin Stability during Processing, Storage, and Its Bioaccessibility. Food Res. Int. 2020, 134, 109244. [Google Scholar] [CrossRef] [PubMed]
- Cepero-Betancourt, Y.; Opazo-Navarrete, M.; Janssen, A.E.M.; Tabilo-Munizaga, G.; Pérez-Won, M. Effects of High Hydrostatic Pressure (HHP) on Protein Structure and Digestibility of Red Abalone (Haliotis rufescens) Muscle. Innov. Food Sci. Emerg. Technol. 2020, 60, 102282. [Google Scholar] [CrossRef]
- Opazo-Navarrete, M.; Tagle Freire, D.; Boom, R.M.; Janssen, A.E.M. The Influence of Starch and Fibre on In Vitro Protein Digestibility of Dry Fractionated Quinoa Seed (Riobamba Variety). Food Biophys. 2019, 14, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved Method for Determining Food Protein Degree of Hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Morr, C.V.; German, B.; Kinsella, J.E.; Regenstein, J.M.; Buren, J.P.V.; Kilara, A.; Lewis, B.A.; Mangino, M.E. A Collaborative Study to Develop a Standardized Food Protein Solubility Procedure. J. Food Sci. 1985, 50, 1715–1718. [Google Scholar] [CrossRef]
- Burgos-Díaz, C.; Rubilar, M.; Morales, E.; Medina, C.; Acevedo, F.; Marqués, A.M.; Shene, C. Naturally Occurring Protein–Polysaccharide Complexes from Linseed (Linum usitatissimum) as Bioemulsifiers. Eur. J. Lipid Sci. Technol. 2016, 118, 165–174. [Google Scholar] [CrossRef]
- Yu, Y.; Fan, F.; Wu, D.; Yu, C.; Wang, Z.; Du, M. Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum. Molecules 2018, 23, 1189. [Google Scholar] [CrossRef] [Green Version]
- Shu, G.; Huang, J.; Bao, C.; Meng, J.; Chen, H.; Cao, J. Effect of Different Proteases on the Degree of Hydrolysis and Angiotensin I-Converting Enzyme-Inhibitory Activity in Goat and Cow Milk. Biomolecules 2018, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, M.; Otte, J.; De Gobba, C.; Osman, A.; Hamad, E. Angiotensin I-Converting Enzyme Inhibitory Activity and Antioxidant Capacity of Bioactive Peptides Derived from Enzymatic Hydrolysis of Buffalo Milk Proteins. Int. Dairy J. 2017, 66, 91–98. [Google Scholar] [CrossRef]
- Goggin, D.E.; Mir, G.; Smith, W.B.; Stuckey, M.; Smith, P.M.C. Proteomic Analysis of Lupin Seed Proteins to Identify Conglutin β as an Allergen, Lup an 1. J. Agric. Food Chem. 2008, 56, 6370–6377. [Google Scholar] [CrossRef]
- Lam, A.C.Y.; Can Karaca, A.; Tyler, R.T.; Nickerson, M.T. Pea Protein Isolates: Structure, Extraction, and Functionality. Food Rev. Int. 2016, 34, 126–147. [Google Scholar] [CrossRef]
- Kramer, R.M.; Shende, V.R.; Motl, N.; Pace, C.N.; Scholtz, J.M. Toward a Molecular Understanding of Protein Solubility: Increased Negative Surface Charge Correlates with Increased Solubility. Biophys. J. 2012, 102, 1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsumura, K.; Saito, T.; Tsuge, K.; Ashida, H.; Kugimiya, W.; Inouye, K. Functional Properties of Soy Protein Hydrolysates Obtained by Selective Proteolysis. LWT Food Sci. Technol. 2005, 38, 255–261. [Google Scholar] [CrossRef]
- Liu, Q.; Lu, Y.; Han, J.; Chen, Q.; Kong, B. Structure-Modification by Moderate Oxidation in Hydroxyl Radical-Generating Systems Promote the Emulsifying Properties of Soy Protein Isolate. Food Struct. 2015, 6, 21–28. [Google Scholar] [CrossRef]
- Shen, H.; Stephen Elmore, J.; Zhao, M.; Sun, W. Effect of Oxidation on the Gel Properties of Porcine Myofibrillar Proteins and Their Binding Abilities with Selected Flavour Compounds. Food Chem. 2020, 329, 127032. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, H.; Xia, X.; Sun, F.; Kong, B. Effect of Ultrasound-Assisted Immersion Thawing on Emulsifying and Gelling Properties of Chicken Myofibrillar Protein. LWT Food Sci. Technol. 2021, 142, 111016. [Google Scholar] [CrossRef]
- Pan, N.; Wan, W.; Du, X.; Kong, B.; Liu, Q.; Lv, H.; Xia, X.; Li, F. Mechanisms of Change in Emulsifying Capacity Induced by Protein Denaturation and Aggregation in Quick-Frozen Pork Patties with Different Fat Levels and Freeze–Thaw Cycles. Foods 2021, 11, 44. [Google Scholar] [CrossRef]
- Tamm, F.; Gies, K.; Diekmann, S.; Serfert, Y.; Strunskus, T.; Brodkorb, A.; Drusch, S. Whey Protein Hydrolysates Reduce Autoxidation in Microencapsulated Long Chain Polyunsaturated Fatty Acids. Eur. J. Lipid Sci. Technol. 2015, 117, 1960–1970. [Google Scholar] [CrossRef]
- Chen, C.; Chi, Y.-J.; Zhao, M.-Y.; Xu, Y. Influence of Degree of Hydrolysis on Functional Properties, Antioxidant and ACE Inhibitory Activities of Egg White Protein Hydrolysate. Food Sci. Biotechnol. 2012, 21, 27–34. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | Type | Biological Source | Supplier | Activity (Under Optimal Conditions) |
|---|---|---|---|---|
| Acid Stable Protease | Aspartic endopeptidase | Aspergillus niger | Bio-Cat (Troy, VA, USA) | 4000 SAP/g |
| Fungal Protease A | Aspartic exo- and endopeptidase | Aspergillus oryzae | Bio-Cat (Troy, VA, USA) | 1,000,000 HUT/g |
| Opti-ZiomeTM P3 HydrolyzerTM | Aspartic exo- and endopeptidase | Aspergillus oryzae, Aspergillus melleus | Bio-Cat (Troy, VA, USA) | 130,000 HUT/g |
| Neutral Protease | Metallo endopeptidase | Bacillus subtilis | Bio-Cat (Troy, VA, USA) | 2,000,000 PC/g |
| Protamex® | Serine endopeptidase | Bacillus licheniformis, Bacillus amyloliquefacies | Novozymes A/S (Bagsværd, Denmark) | 1.5 AU-N/g |
| Alcaline Protease L | Serine endopeptidase | Bacillus licheniformis | Bio-Cat (Troy, VA, USA) | 625,000 DU/g |
| Alcalase® 2.4 L FG | Serine endopeptidase | Bacillus licheniformis | Novozymes A/S (Bagsværd, Denmark) | 2.4 AU-A/g |
| Enzyme | E/S (%) | Temperature (°C) | pH Value | Time (h) |
|---|---|---|---|---|
| Acid Stable Protease | 0.01/0.02/0.04 | 55 | 2.5 | 1/2/5 |
| Fungal Protease A | 0.01/0.02/0.04 | 60 | 3.0 | 1/2/5 |
| Opti-ZiomeTM P3 HydrolyzerTM | 0.01/0.02/0.04 | 60 | 6.0 | 1/2/5 |
| Neutral Protease | 0.01/0.02/0.04 | 55 | 7.0 | 1/2/5 |
| Protamex® | 0.01/0.02/0.04 | 55 | 8.0 | 1/2/5 |
| Alcaline Protease L | 0.01/0.02/0.04 | 55 | 8.5 | 1/2/5 |
| Alcalase® 2.4 L FG | 0.01/0.02/0.04 | 70 | 9.0 | 1/2/5 |
| Functional Property | Hydrolysis Time (h) | E/S (%) | Acid Stable Protease | Fungal Protease A | Opti-ZiomeTM P3 HydrolyzerTM | Neutral Protease | Protamex® | Alcaline Protease L | Alcalase® 2.4 L FG |
|---|---|---|---|---|---|---|---|---|---|
| EC (%) | 1 | 0.01 | 85.7 ± 0.6 b,F | 83.3 ± 1.5 a,E | 73.3 ± 1.2 a,B | 80.0 ± 0.0 a,D | 78.0 ± 1.0 b,C | 70.3 ± 0.6 a,A | 78.0 ± 1.0 b,C |
| 0.02 | 82.0 ± 2.0 a,C | 86.7 ± 0.6 a,D | 72.3 ± 0.6 a,A | 77.7 ± 2.5 a,B | 77.7 ± 1.5 b,B | 78.3 ± 0.6 b,B | 74.7 ± 0.6 a,A | ||
| 0.04 | 83.3 ± 1.5 a,b,C | 82.7 ± 3.2 a,C | 71.7 ± 1.5 a,A | 82.7 ± 4.0 a,C | 75.0 ± 0.1 a,A,B | 77.7 ± 2.1 b,B | 75.3 ± 0.6 a,A,B | ||
| 2 | 0.01 | 86.0 ± 1.7 b,E | 84.3 ± 1.2 a,D | 73.3 ± 0.6 a,A | 77.7 ± 0.6 a,B | 79.7 ± 0.6 b,C | 78.3 ± 0.6 a,B,C | 79.7 ± 0.6 b,C | |
| 0.02 | 86.3 ± 1.5 b,D | 82.0 ± 1.0 a,C | 74.3 ± 0.6 b,A | 79.0 ± 1.0 a,B | 79.0 ± 1.0 b,B | 77.0 ± 2.6 a,B | 79.0 ± 1.0 a,b,B | ||
| 0.04 | 75.7 ± 0.6 a,A,B | 83.0 ± 2.0 a,C | 75.0 ± 0.1 b,A | 78.3 ± 2.9 a,B | 75.0 ± 1.0 a,A | 77.0 ± 2.0 a,A,B | 76.3 ± 2.3 a,A,B | ||
| 5 | 0.01 | 86.0 ± 2.0 b,D | 84.7 ± 1.5 b,D | 71.7 ± 1.2 a,b,A | 75.3 ± 1.2 a,B | 84.0 ± 1.7 b,D | 80.3 ± 0.6 c,C | 84.0 ± 1.7 b,D | |
| 0.02 | 83.3 ± 1.2 b,D | 81.7 ± 2.3 a,b,D | 71.3 ± 0.6 a,B | 82.0 ± 2.6 b,B | 81.7 ± 1.5 b,D | 66.3 ± 1.5 b,A | 77.7 ± 1.5 a,C | ||
| 0.04 | 80.3 ± 0.6 a,D | 80.3 ± 1.2 a,D | 73.0 ± 0.0 b,B | 85.3 ± 1.2 b,E | 77.7 ± 1.5 a,C | 61.3 ± 2.3 a,A | 79.3 ± 1.2 a,C,D | ||
| ES (%) | 1 | 0.01 | 81.3 ± 1.1 a,A | 88.0 ± 2.2 b,B | 95.9 ± 1.4 a,C | 97.1 ± 4.0 b,C,D | 98.7 ± 1.3 b,C,D | 90.6 ± 1.9 a,B | 99.5 ± 0.8 c,D |
| 0.02 | 86.6 ± 1.5 b,B | 81.2 ± 0.6 a,A | 96.8 ± 0.8 a,E | 90.4 ± 0.7 a,b,C | 94.0 ± 0.8 a,D | 93.4 ± 1.6 a,D | 93.6 ± 1.3 b,D | ||
| 0.04 | 84.8 ± 2.2 b,A | 84.7 ± 2.3 a,b,A | 97.7 ± 1.6 a,C | 84.8 ± 4.4 a,A | 91.0 ± 3.3 a,B | 96.6 ± 0.8 b,C | 88.9 ± 0.8 a,A,B | ||
| 2 | 0.01 | 79.9 ± 1.4 a,A | 87.0 ± 1.0 a,B | 95.9 ± 0.0 a,E | 94.4 ± 2.0 b,D,E | 93.7 ± 1.2 a,D,E | 89.8 ± 1.2 a,C | 93.3 ± 1.4 a,D | |
| 0.02 | 82.3 ± 2.4 a,A | 88.2 ± 1.5 a,B | 95.5 ± 3.3 a,D | 94.9 ± 0.1 b,C,D | 92.4 ± 2.1 a,C,D | 91.8 ± 1.7 a,B,C | 92.4 ± 2.1 a,C,D | ||
| 0.04 | 94.3 ± 1.5 b,B,C | 88.4 ± 3.4 a,A | 93.8 ± 0.8 a,B,C | 89.0 ± 2.6 a,A | 96.9 ± 0.7 b,C | 96.1 ± 1.2 b,C | 90.9 ± 2.4 a,A,B | ||
| 5 | 0.01 | 81.4 ± 1.0 a,A | 85.5 ± 2.5 a,B | 97.7 ± 0.8 b,D | 93.1 ± 0.9 b,C | 97.9 ± 1.9 b,D | 91.3 ± 1.3 a,C | 92.1 ± 1.7 a,C | |
| 0.02 | 85.2 ± 0.6 b,A | 87.4 ± 2.6 a,A,B | 98.1 ± 0.8 b,E | 94.7 ± 1.4 b,D | 92.1 ± 1.7 a,C,D | 90.0 ± 1.5 a,B,C | 97.9 ± 1.9 b,E | ||
| 0.04 | 87.6 ± 0.1 c,A | 88.0 ± 1.6 a,A,B | 93.2 ± 2.4 a,C | 87.5 ± 1.3 a,A | 89.8 ± 1.7 a,A,B,C | 91.3 ± 4.0 a,B,C | 93.3 ± 0.7 a,C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opazo-Navarrete, M.; Burgos-Díaz, C.; Garrido-Miranda, K.A.; Acuña-Nelson, S. Effect of Enzymatic Hydrolysis on Solubility and Emulsifying Properties of Lupin Proteins (Lupinus luteus). Colloids Interfaces 2022, 6, 82. https://doi.org/10.3390/colloids6040082
Opazo-Navarrete M, Burgos-Díaz C, Garrido-Miranda KA, Acuña-Nelson S. Effect of Enzymatic Hydrolysis on Solubility and Emulsifying Properties of Lupin Proteins (Lupinus luteus). Colloids and Interfaces. 2022; 6(4):82. https://doi.org/10.3390/colloids6040082
Chicago/Turabian StyleOpazo-Navarrete, Mauricio, César Burgos-Díaz, Karla A. Garrido-Miranda, and Sergio Acuña-Nelson. 2022. "Effect of Enzymatic Hydrolysis on Solubility and Emulsifying Properties of Lupin Proteins (Lupinus luteus)" Colloids and Interfaces 6, no. 4: 82. https://doi.org/10.3390/colloids6040082
APA StyleOpazo-Navarrete, M., Burgos-Díaz, C., Garrido-Miranda, K. A., & Acuña-Nelson, S. (2022). Effect of Enzymatic Hydrolysis on Solubility and Emulsifying Properties of Lupin Proteins (Lupinus luteus). Colloids and Interfaces, 6(4), 82. https://doi.org/10.3390/colloids6040082