4.1. Plio-Pleistocene Rucervus from Europe
Cervus ardei Croized & Jobert, 1828 from the Late Pliocene of Perrier-Etouaires (France) is the earliest cervid from the European paleontological record that displays a bauplan close to the
Rucervus antler. Heintz [
25] included
Cervus ardei in the new monotypic genus
Arvernoceros, which is regarded here as the subgenus of
Rucervus. The total length of the type antler of
R. (
Arvernoceros)
ardei (no number, MNHN) attains 610 mm [
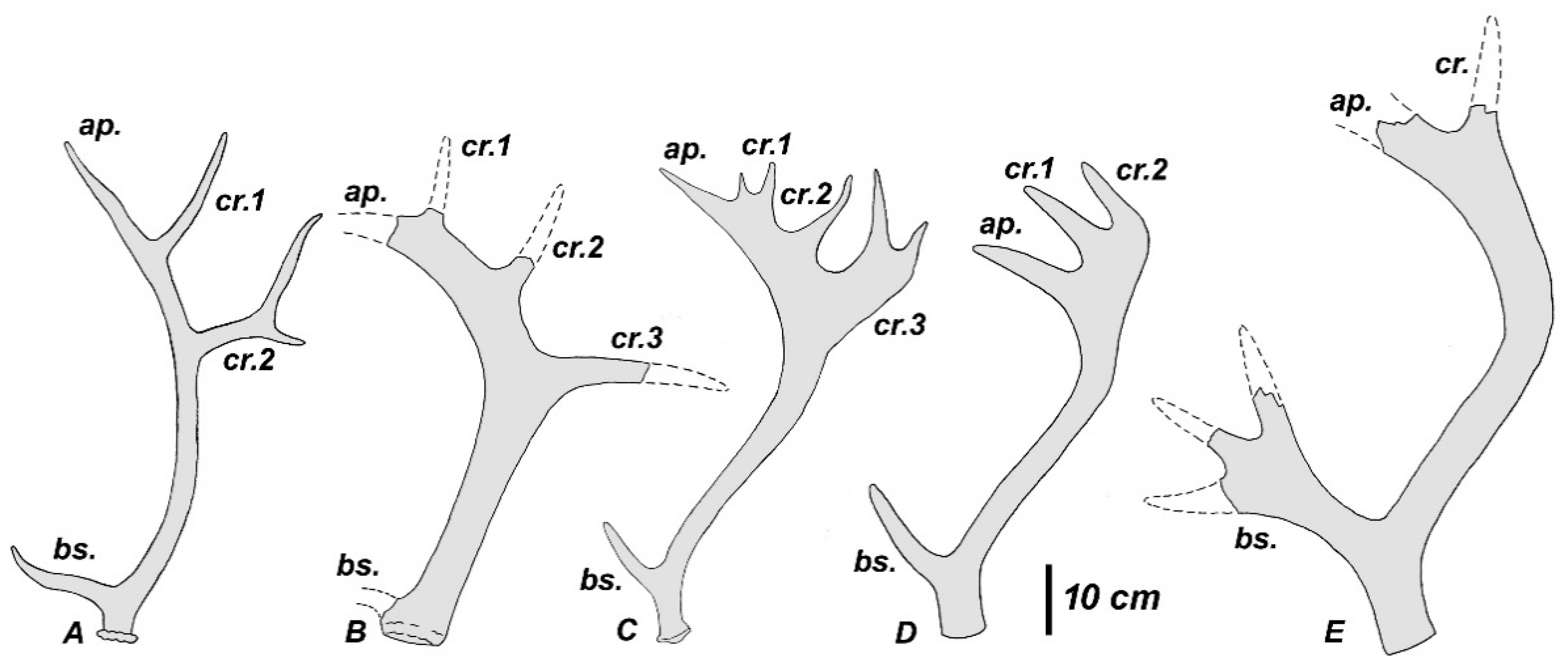
25]. The first tine in the type specimen is situated rather high above the burr: the first ramification height measures more than twice as much as the DAP of the antler base. The first tine is supplemented with an accessory prong, as in modern
R. (
Rucervus)
duvaucelii (not visible on the
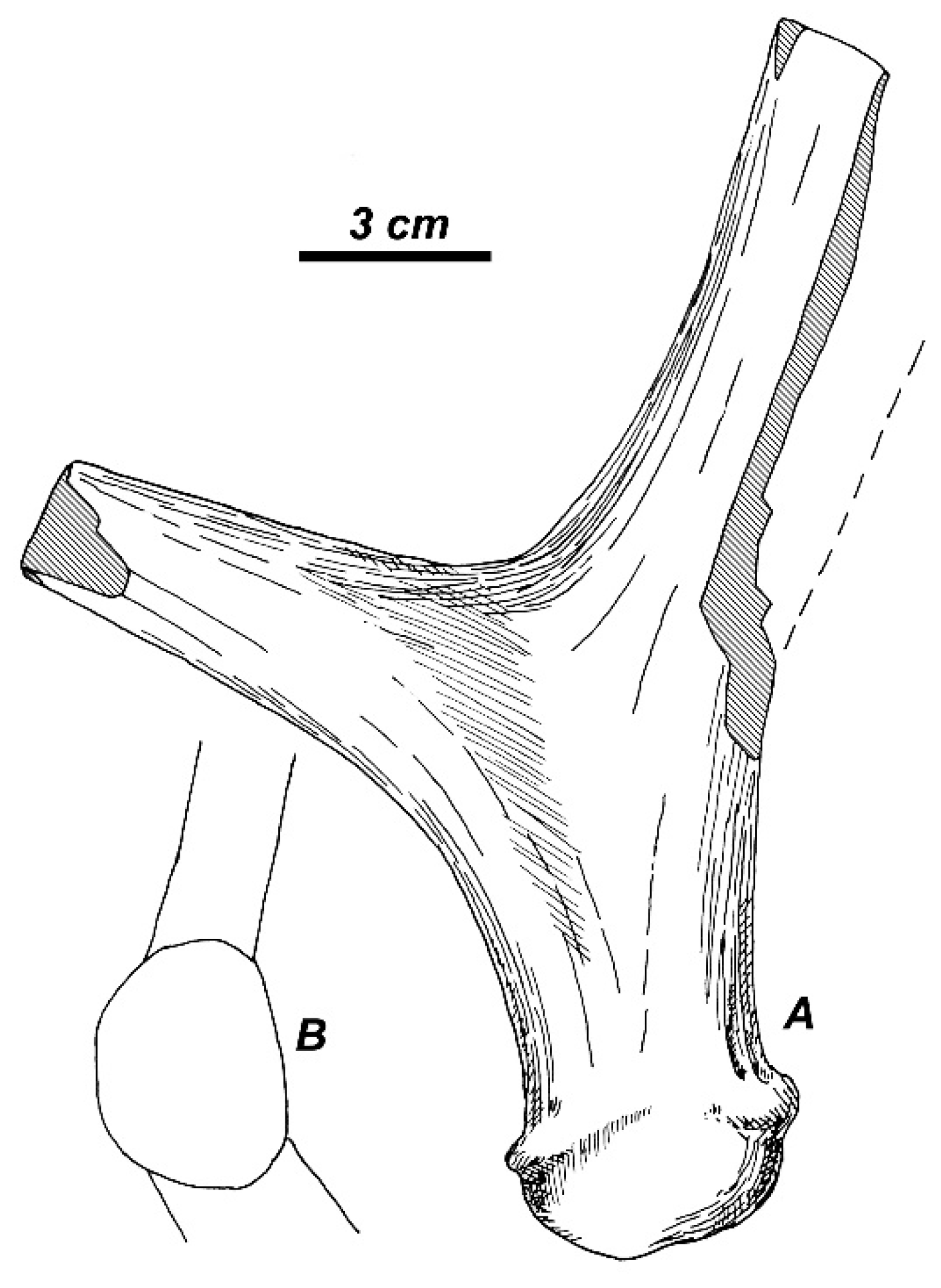
Figure 15). This accessory prong may be variable in shape and position and in some cases is situated in the area of ramification. The middle tine is never present. The antler beam is cylinder-shaped and curved backward from the first ramification and then somewhat bowed anteriorly in its distal part. The antler is terminated by a small palmation with three tines (the maximum number of tines recorded for this species), which are pointed upward and forward (
Figure 15D) and do not show the typical for
Rucervus crown construction with distal tines being inserted on the posterior side of the beam. Nonetheless, the direction of two posterior tine may not be exact, since the antler is reconstructed at the base of each tine. Even if the shape of palmation is reconstructed correctly, it may represent an individual variation, which deviated from the “initial” described for
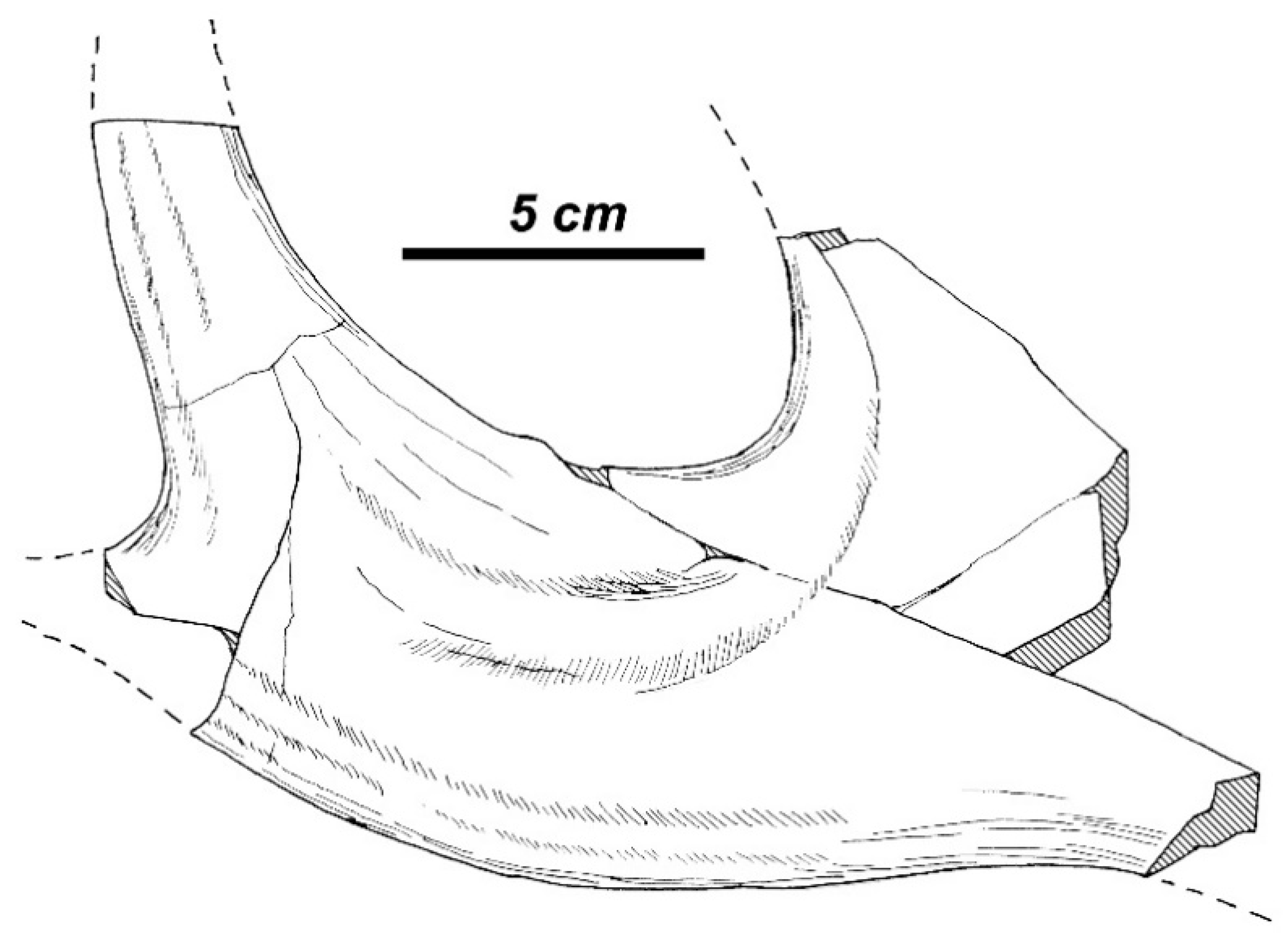
Rucervus antler construction pattern. The distal fragment of antler PET1024 (MNHN) is of interest for this discussion, since it represents a part of palmation resembling the posterior palmation lobe with two terminal prongs in
R. radulescui (
Figure 16). Therefore, the distal tines of the type specimen of
Cervus ardei could be interpreted as the apical tine and the two posterior tines that form a posterior lobe of palmation. Heintz [
25] ascribed to
R. (
Arvernoceros)
ardei upper molars with a strongly developed cingulum. Mandibles and postcranial bones of large cervids from Perrier-Etouaires represent an unresolved problem, since the type locality has yielded the remains of another deer species of similar size
Praeelaphus perrieri [
25,
44]. The pedicles of
R. (
Arvernoceros)
ardei are quite long, cylinder-shaped, and little divergent, and therefore represent the most primitive morphological condition within
Rucervus. Other cranial characters of
R. ardei remain unknown. The specific morphological traits of
R. ardei that could be regarded as diagnostic for the subgenus
Arvernoceros are the very high position of the first ramification and the relatively short crown part of the antler that attains less than one-third of the total antler length.
Heintz [
25] assumed the phylogenetic relationship between
Rucervus ardei and
Megaloceros giganteus, however, he acknowledged that this hypothesized phylogenetic link is not demonstrated. Vislobokova [
45] supported the direct phylogenetic relationship between
Rucervus (
Arvernoceros)
ardei and
M. giganteus. This link, according to the cited authors, is based on such shared characters as the development of small distal palmation with distal tines directed towards the anterior, and the presence of cingulum in the upper molars. Actually, cingulum is a quite variable morphological feature in giant deer and it is well-developed mostly in the better known Irish sample, while the continental forms of
M. giganteus are characterized by a very weak lingual cingulum in the upper molars [
44], therefore, this character, as well as the apomorphic development of distal palmation serve as weak arguments for the evolutionary link between
Arvernoceros and
Megaloceros.
R. verestchagini (David, 1992) is a much larger species coming from the Early Pleistocene of Salcia (Moldova). The general antler bauplan is very similar to that of
R. ardei, however, besides the extremely large size—the diameter of its antler base is ca. 10 cm—
R. verestchagini is characterized by the poor development of crown (
Figure 15E), which is represented in the type specimen by a simple fork formed by the apical part of the beam and the thinner posterior tine, and the particularly large and flattened first tine terminated by three prongs [
46].
Another comparatively large cervid has been reported from Liventsovka (Russia) as
Arvernoceros sp. [
34,
47]. As
R. ardei and
R. verestchagini, the deer from Liventsovka is characterized by the high position of the first tine (
Figure 14B). The pedicles are cylindrical, robust, and divergent. The shape of basal tine and the distal part of the antler are unknown. Titov [
34] reported from Liventsovka some fragments of palmated antlers as Cervidae gen. indet., which, possibly, might belong to this cervid form. The size of the deer from Liventsovka is quite large but smaller than that of
R. verestchagini. Since the main diagnostic characters of this deer (the shape of the first tine and the shape of distal portion) remain unknown, this cervid is regarded here as
Rucervus (
Arvernoceros) sp. The high position of the first tine may suggest its phylogenetic relationship with European forms. The geological age of the large
Rucervus from Liventsovka remains unclear, since the Livenzovka collection seems to include material from two different stratigraphic levels at least: one of early Villafranchian age (MN16b) and another one quite younger (MN17) [
48,
49].
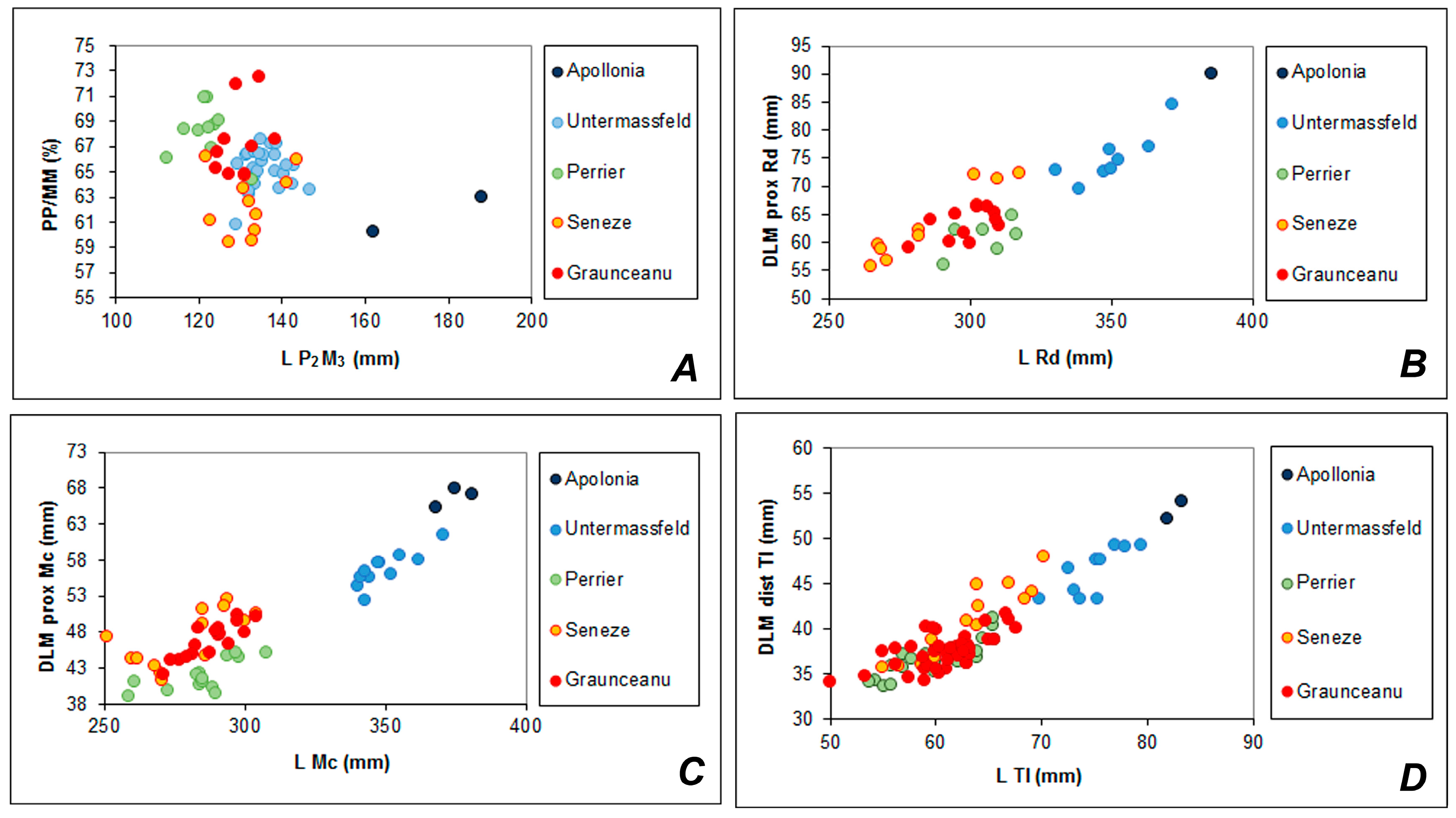
The large-sized deer from the final Villafranchian from Saint-Prest (France), Rosières (France), and Untermassfeld (Germany) is another cervid form that should be included in the group of European
Rucervus. This is a conspicuously long-limbed and large-sized deer (larger than European
Eucladoceros) with a primitive lower fourth premolar and a practically absent cingulum in the upper molars. This deer was described under different names:
Cervus (
Megaceros)
dupuisi Shehlin, 1912 based on the sample from Rosières [
50];
Eucladoceros giulii Kahlke, 1997 based on the sample from Untermassfeld [
37]; and, reported by Guerin et al. [
51] as
Praemegaceros verticornis (Dawkins) from Saint-Prest [
19]. According to Azzaroli [
52], some mandibles from the Forest Bed Formation (England) of the intermediary between
P. verticornis and
Praemegaceros dawkinsi size are very similar to
Megaceros dupuisi from Rosieres. The mandibles in question from the Forest Bed Formation were ascribed by Azzaroli [
52] to
Praedama savini (=
Megaceros savini according to Azzaroli, 1953). The complete mature antlers of this cervid form are unknown. Kahlke [
37] proposed a hypothetical reconstruction of antlers that shows the comb-like bauplan of European
Eucladoceros ctenoides. Actually, a “typical”
E. ctenoides is present in the roughly coeval with Untermassfeld French site of Ceyssaguet where this species is represented by fine complete antlers and it is characterized by “normal” for European
Eucladoceros body size [
53]. Unlike
E. ctenoides from Ceyssaguet, the cervid from Untermassfeld is characterized by the relatively longer lower premolar series and the unsplit lingual walls in the upper molars [
38]. The hypothesized antler reconstruction proposed by Kahlke [
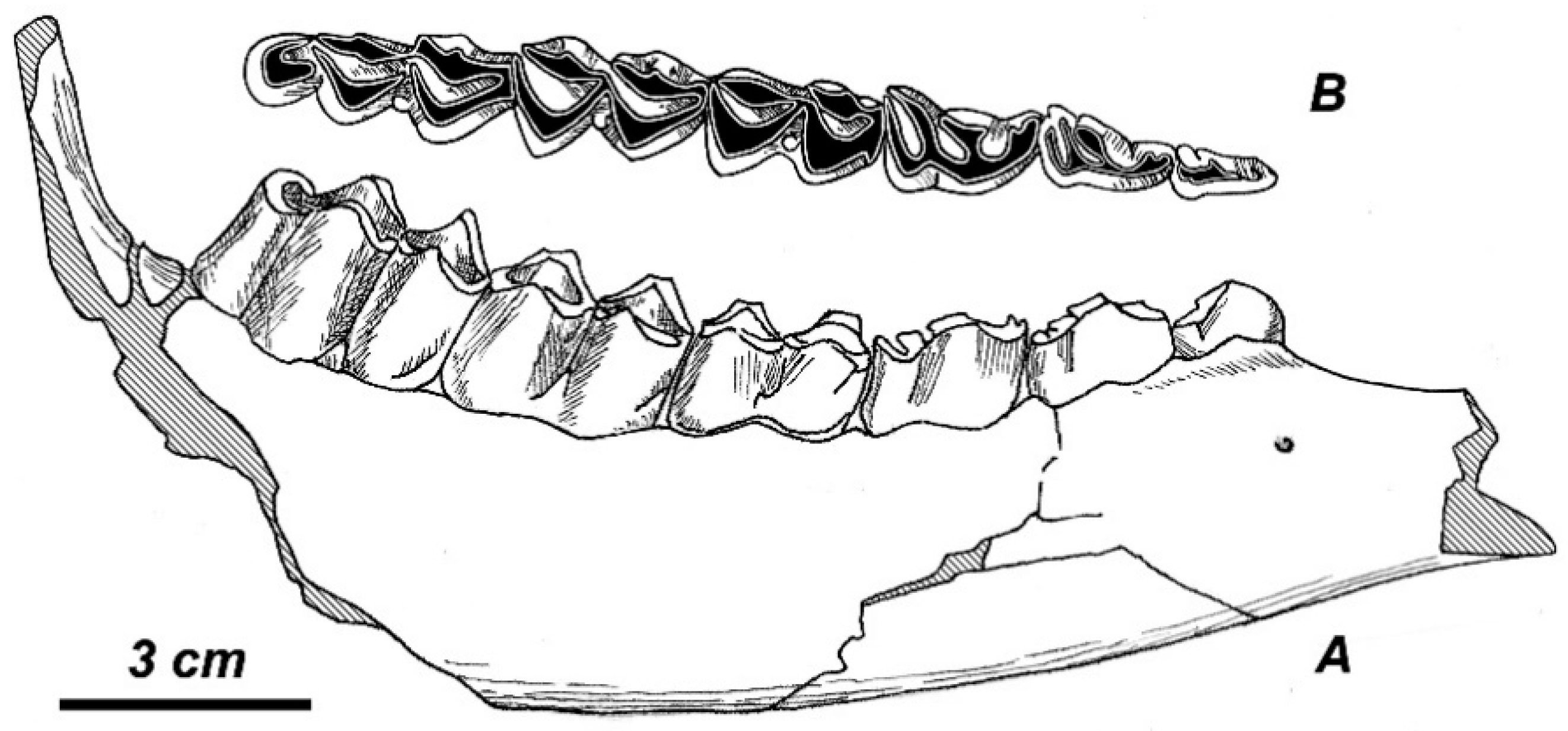
37] has been questioned by Croitor and Kostopoulos [
38]. The only complete antler from Untermassfeld is juvenile (p. 229, Figure 26, [
37]), but it shows some features characteristic of
Rucervus (
Arvernoceros): the first tine is situated high above the burr, flattened from the sides, and terminated with a bifurcation. The angle of the first ramification is 55°. The main beam is somewhat flattened in the area of distal bifurcation. The general morphological specialization of the antler under discussion recalls
R. verestchagini [
38].
4.2. Plio-Pleistocene Rucervus from the Siwaliks and Caucasus
The cervid fossil record from the Siwaliks and Caucasus is significant for the understanding of the origin and systematic position of Early Pleistocene deer from southeastern Europe, however, this important source on the geological history of cervids is undeservedly ignored.
Up to seven deer species and forms are reported from the Siwalik Hills. The Siwalik cervid species were established mostly on the morphology of upper molars, the fact that limits the understanding of their taxonomic status and systematic position. As a matter of fact, only three types of antlers have so far been reported from the Siwaliks, which attest the presence of the genera
Metacervocerus (the sister genus of modern
Axis),
Panolia (reported by Azzaroli [
54] as
Rucervus sp. I and
Euctenoceros sp.), and
Rucervus [
36,
54,
55]. Another smaller
Axis-like deer
Cervus rewati is described by Arif et al. [
56] from the Upper Siwaliks of Pakistan.
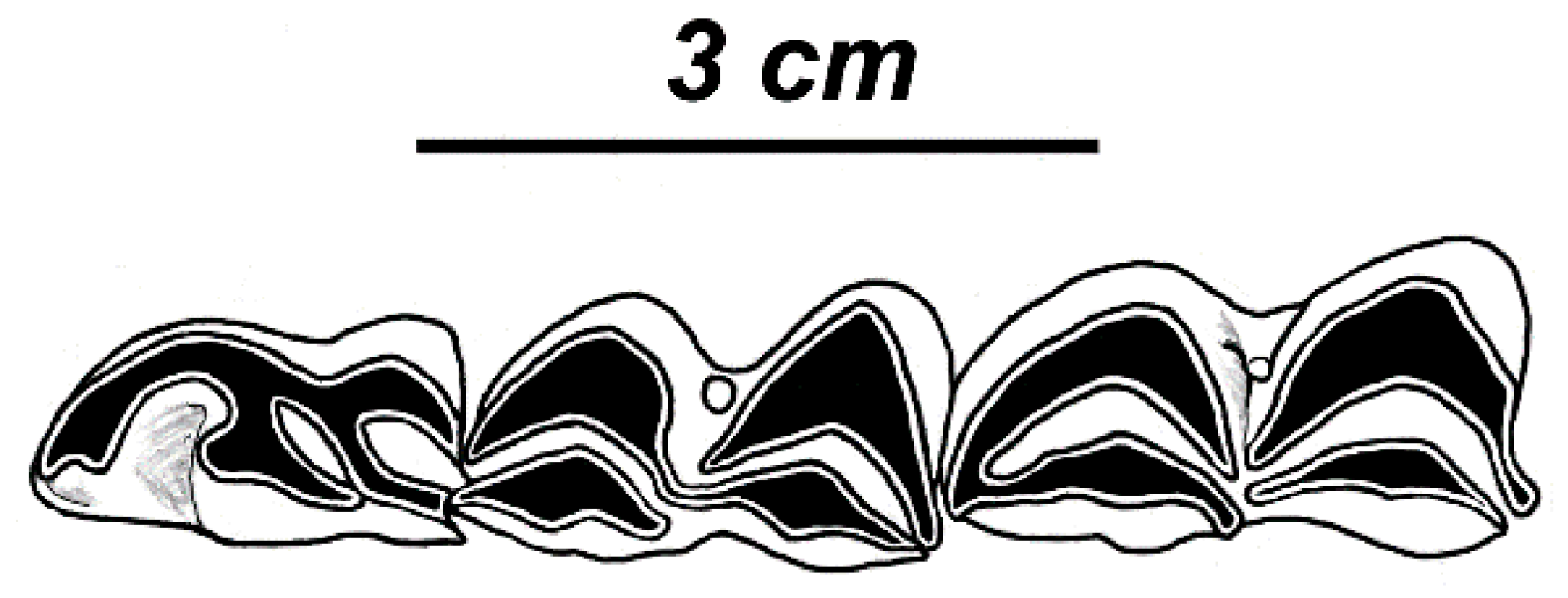
Cervus latidens Lydekker, 1876 was described on the basis of few very large isolated molars [
57]. The dental remains of this poorly known ruminant form are as large as those of the deer from Apollonia measurements of M
3 (L = 31.2 mm [1.23 in.], D = 25. 4 mm [1.0 in.], the measurements are adapted from Lydekker [
57]), but the tooth crown of the Siwalik form is relatively narrower in the linguolabial dimension. This is a dental characteristic correlated with the comparatively more advanced degree of hypsodonty in the Siwalik teeth. Besides that, the figures of upper molars (Pl. VIII, Figures 4, 6, 7, 10, [
58]) show the typical bovid characteristics: the anterior and posterior walls of molars are parallel, the entostyle is high and column-shaped, and the hypoconal enamel spur is missing (the hypoconal spur is present in
R. gigans). According to Lydekker [
58], the initial inclusion of this species to Cervidae was arbitrary and later he included “
Cervus latidens” in the bovid genus
Oreas [
59]. This taxonomic revision was never contested afterwards.
Cervus triplidens Lydekker, 1876 is another questionable ruminant species from the Siwaliks included in the family Cervidae. The species is based on the maxilla with high-crowned M
2-M
3 ([
58]: Pl. VIII, Figures 1 and 2), is characterized by high and strong column-shaped entostyles, and therefore looks rather like a bovid. Koken [
60] expressed his doubts about the attribution of the specimen in question to a cervid. It is important to mention that Lydekker [
59] himself indicated in the footnote on page 116 the “superficial” similarity of dental morphology of
C. triplidens with
Boselaphus sp. According to Colbert [
36], this species is based on a very meagre material and therefore is of “little value”. Colbert [
36] provisionally ascribed to
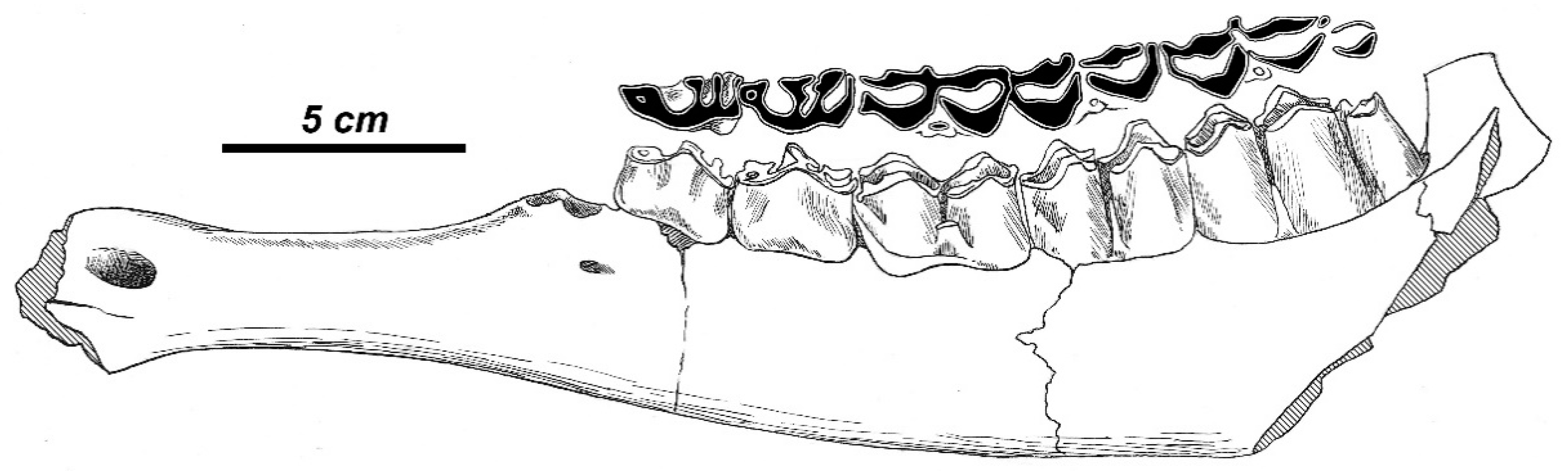
C. triplidens the left maxilla No. 19792 (AMNH) with P
3-M
3. This specimen shows a typical cervid morphology. The molars are low crowned and are supplemented with a quite strong and flat entostyle (not column-shaped as in the type specimen), but are devoid of a hypoconal spur.
Cervus simplicidens is the only Lydekker’s species from the Siwaliks (Pl. VIII, Figure 3, [
57]) that is certainly based on true cervid remains: the upper molars of this species are supplemented by a small lingual cingulum and a small entostyle [
54]. The stratigraphic provenance of this cervid form is uncertain (Middle or Upper Siwaliks [
36,
54]). According to Lydekker [
58], the general shape of the molars is very similar to those of the living
Rucervus duvaucelii. However, another maxilla with P
3-M
3 (Pl. VIII, Figure 6, [
59]) described as
C. simplicidens, again, shows the typical bovid morphological characters: massive column-like entostyles, high crowns with parallel anterior and posterior walls, and no cingulum. Colbert [
36] described as
C. simplicidens a juvenile cervid right maxilla with deciduous teeth and two molars supplemented with a small entostyle and a hypoconal enamel spur. Azzaroli [
54] ascribed to
C. simplicidens a fine male skull B.M. 39570 (NHML), noticing the great similarity of its molar morphology with the type specimen. The skull also displays a great morphological affinity—except for the relatively shorter muzzle—with modern
R. duvaucelii, the fact that permitted to Azzaroli [
54] to include the species under discussion in the genus
Rucervus. The available for observation morphological characters of the cranium BM39570 (the small upper canines; the large ethmoidal openings with a long border formed by nasal bones at more than ½ of ethmoidal opening length; the position of the anterior edges of the orbits above the posterior edge of M
3) confirm the opinion of Azzaroli [
54].
“Cervus” sivalensis Lydekker, 1876 is based on the lower molars that are figured by Lydekker [
57] in the Tab. XIII, Figure 5. Originally, Lydekker (
ibidem) supposed that this specimen could belong to
Cervus triplidens, but in the introduction chapter, which was prepared somewhat later, Lydekker proposes for this specimen a new species name
Cervus sivalensis. Later on, Lydekker [
59] ascribed to this species the quite worn M
2 and M
3, which, however, provide the typical cervid morphology and they are supplemented with a small lingual cingulum. According to Lydekker (
ibidem), the size and morphology of this specimen agree with those of modern
R. duvaucelii. The morphology of the upper third molar figured by Lydekker (p. 122, Figure 8, [
59]) strongly recalls the type specimen of
Cervus simplicidens and most probably should be ascribed to this species. Azzaroli [
54] regarded this specimen (BM48440, NHML) as the holotype of “
Cervus”
sivalensis, but this is not the case, since the species originally was based on the lower molars figured in Lydekker [
57]. Actually, the type specimen of “
Cervus”
sivalensis provides poor diagnostic characters and its taxonomic status for the moment is unclear since the Siwalik fauna contains another cervid of the similar size,
Metacervocerus punjabiensis (Brown, 1926) [
61,
62]. Colbert [
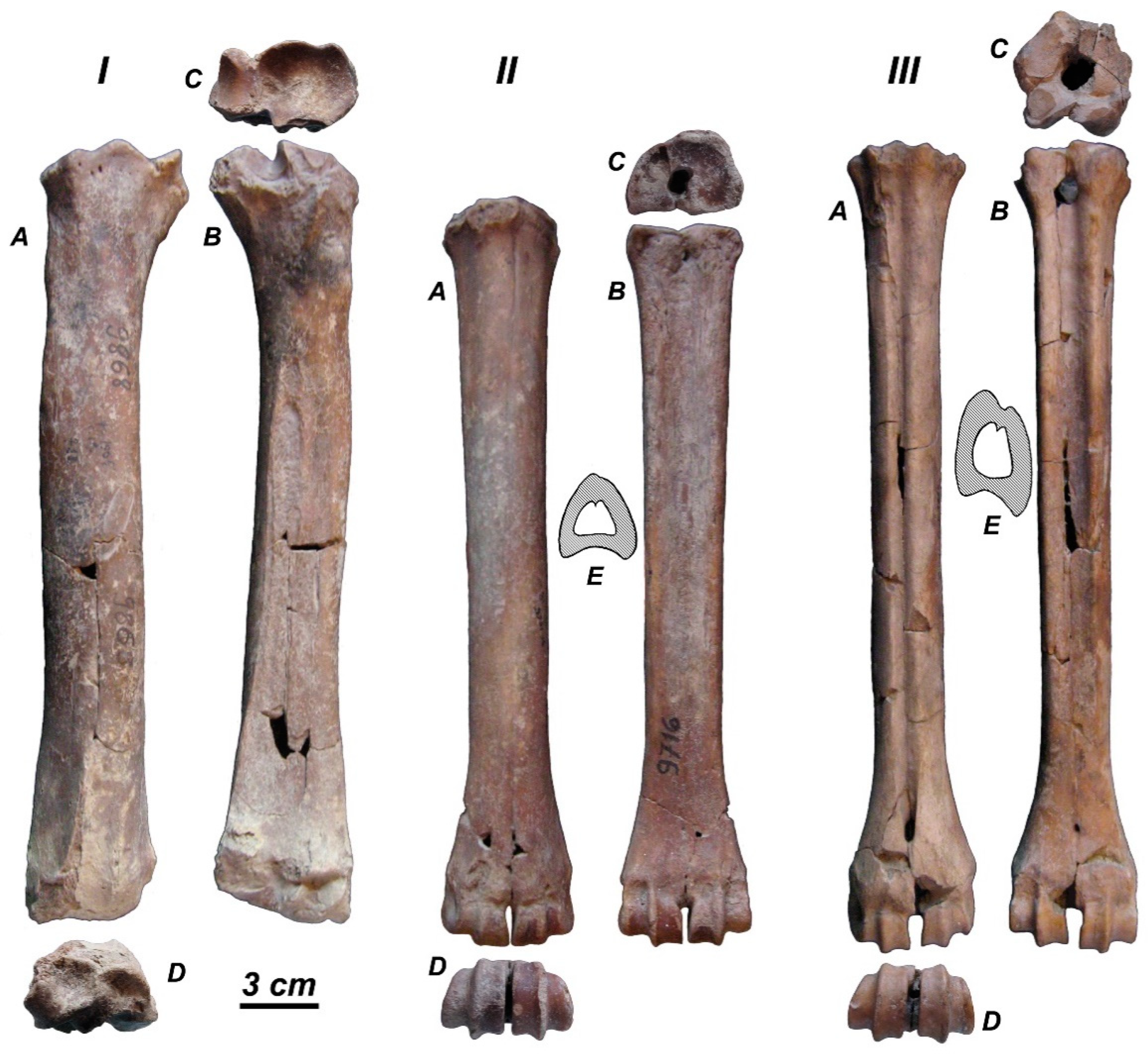
36] ascribed to “
Cervus”
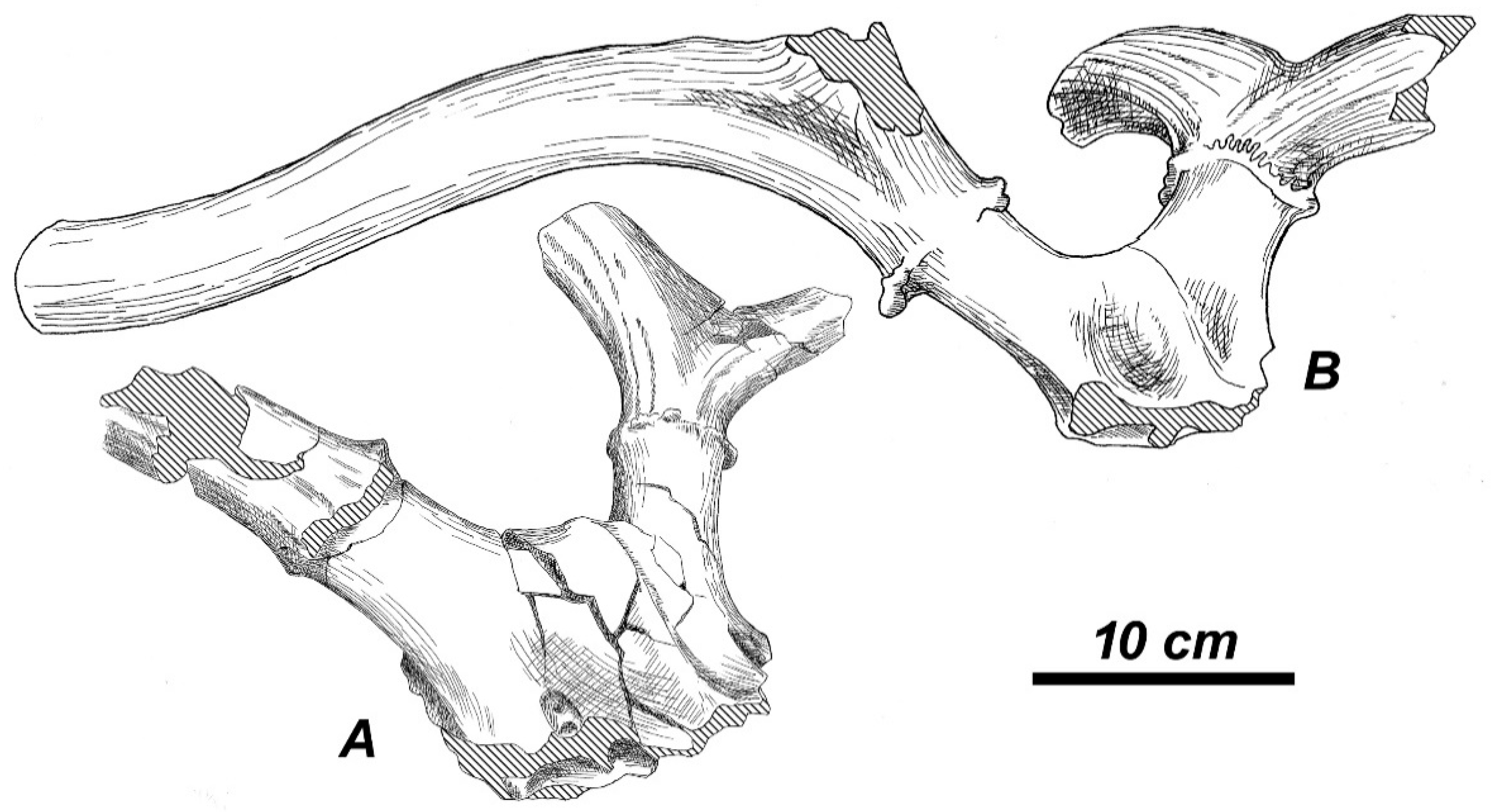
sivalensis a well-preserved skull No. 19829 (AMNH) and the almost complete antler No. 19807 (AMNH), which strongly resemble
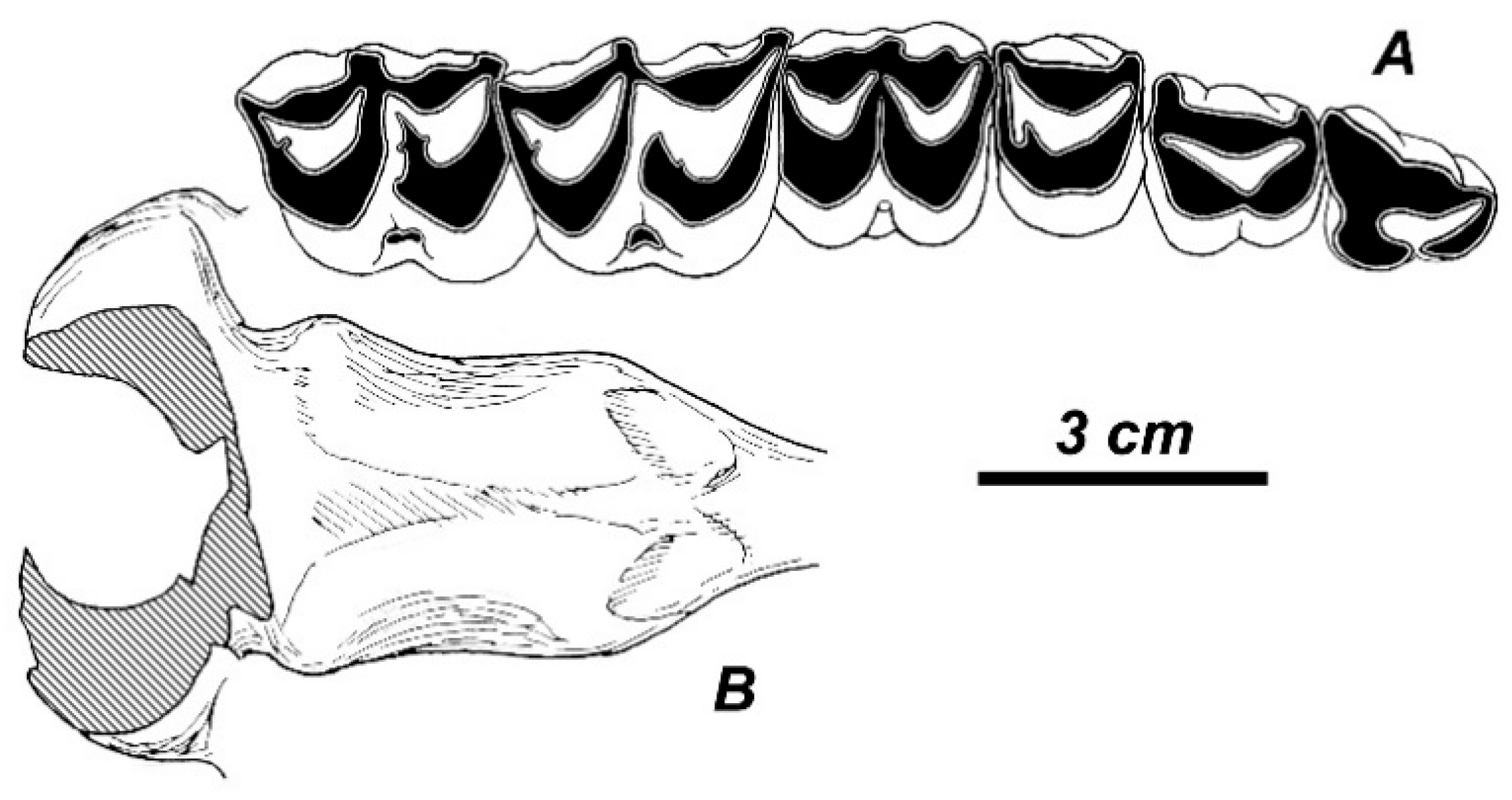
R. duvaucelii (
Figure 15A,B). The antler is heavily built, with round cross-section; the basal tine is situated very close to the burr; the antler crown is formed by at list three crown tines that were inserted on the posterior side of the distal part of the beam [
36], as in modern
R. duvaucelii. The upper molars of the skull are deeply worn, however, as one can see on the photographic image (Figure 145, [
36]), there is a weak but clear lingual cingulum [
54].
Azzaroli [
54] proposed a new species name
Cervus colberti for the skull No. 19829 (AMNH), since, according to the cited author, it is distinguished from
R. simplicidens by a somewhat larger size. Azzaroli (
ibidem) distinguishes his new species from “
Cervus”
sivalensis by smaller cingulum and more rounded protocone and hypocone. Actually, the degree of cingulum expression may be an individually variable character, while the lingual outlines of protocone and hypocone depend on the stage of molar wear, which seems to be very advanced in the type specimen of
Rucervus colberti (Figure 145, [
36]). The proposed in my previous work [
55] body mass estimation based on the figure of skull 19829 (AMNH), as published by Colbert [
36], is exaggerated. In fact, the body size of
R. colberti is not very different from that of
R. simplicidens and it generally corresponds to the size of
Eucladoceros. It is difficult to say whether
R. colberti (Azzaroli, 1954) is indeed a larger advanced form closely related to
R. simplicidens (Lydekker, 1876) since the measurements of the discussed cervid remains from the Siwaliks stored in NHML [
54] are still missing. There is a probability that we are dealing with only one species
R. simplicidens and
R. colberti is its junior synonym.
The paleontological record of the Caucasian region brings several poorly known and poorly understood ruminant forms reported as cervids. Some of them seem to be related to the Siwalik species, however, a direct comparative study was never done.
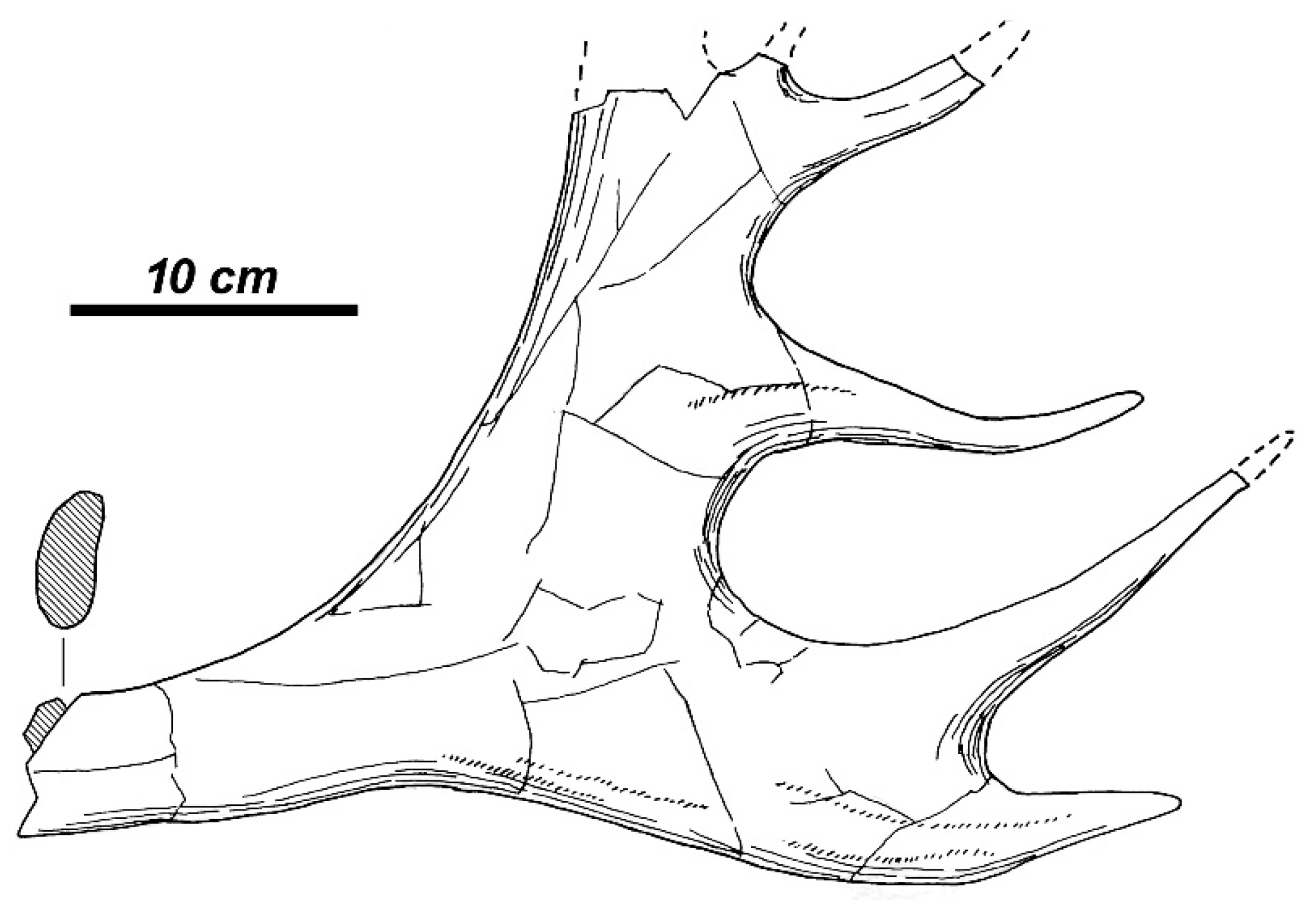
Pseudalces mirandus Flerov, 1962 is an imperfectly known very large ruminant from southeastern Europe based on very poor cranial material from the mixed Pliocene fauna of Kosiakino Quarry (South Russia, Stavropol). The new genus and the new species were proposed by Flerov [
63] on the basis of the orbitofrontal part of the skull with upper left tooth row P
4-M
3. Flerov and Shevyreva [
64] suggested the morphological affinity of
Pseudalces with
Eucladoceros but excluded the possibility of the direct phylogenetic relationship between those two genera. Vislobokova [
65] regards
P. mirandus as a side phylogenetic branch of true elks (Alceini). The similar large ruminant form was described by Vekua [
66] from the late Early Pliocene of Kvabebi (Georgia) as
Pseudalces sp. The dental remains from Kosiakino Quarry and Kvabebi belong to a rather large ruminant (the estimated body mass exceeds 400 kg) and they show some morphological peculiarities that cast doubts on their attribution to a cervid: the protocone in the upper molars has no additional enamel fold as it could be expected in primitive elks, while the premolars remain relatively large, distinguishing the specimens under question from Cervinae. The enamel “pockets” from the labial side of the upper molars from Kvabebi remind the morphological condition seen in
Giraffa punjabiensis (Figure 195, [
36]). The systematical position of
P. mirandus from Kosiakino Quarry and
Pseudalces sp. from Kvabebi requires a revision and possibly those remains actually belong to Giraffidae.
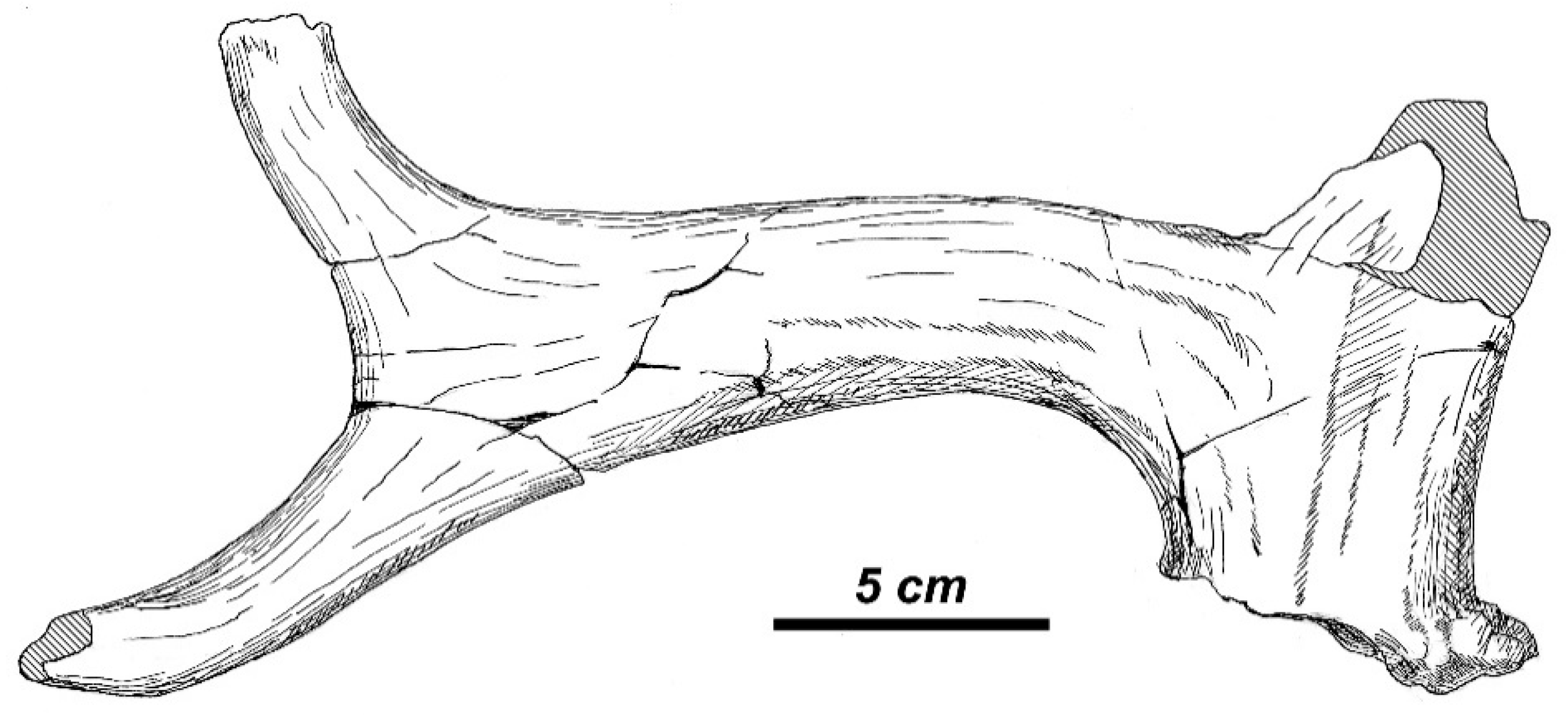
The somewhat smaller ruminant from Kvabebi reported by Vekua [
66] as
Eucladoceros sp. is a cervid similar in body size to
Eucladoceros from Sénèze. This deer form is characterized by the certain distance between the antler burr and the first tine, and the laterally compressed antler portion in the area of the first ramification (p. 202, Figure 50, [
66]). The basal part of the first tine is flattened and forms a right angle with the main beam, which, in its turn, is sharply bent backwards from the first ramification, as in
Rucervus from the Siwaliks figured by Azzaroli [
54]. Pedicles are quite short (the pedicle diameter exceeds the pedicle length), cylindrical, and divergent, with an angle of divergence of 95°. The metatarsal bone from Kvabebi (p. 202, Figure 51, [
66]) fits the size and proportions of
R. radulescui. The mandible K-1022 described as
Pseudalces sp. (p. 207, Figure 53, [
66]) certainly belongs to this cervid form. This specimen is characterized by the low mandibular body, the primitive shape of P
4, and the relatively long premolar series. The mandible from Kvabebi is distinguished from the sample of
Eucladoceros from Sénèze by its relatively longer premolar series, which is regarded here as a primitive morphological condition. The size and dental row proportion of the specimen K-1022 is very similar to those of
R. radulescui from Valea Grăunceanului.
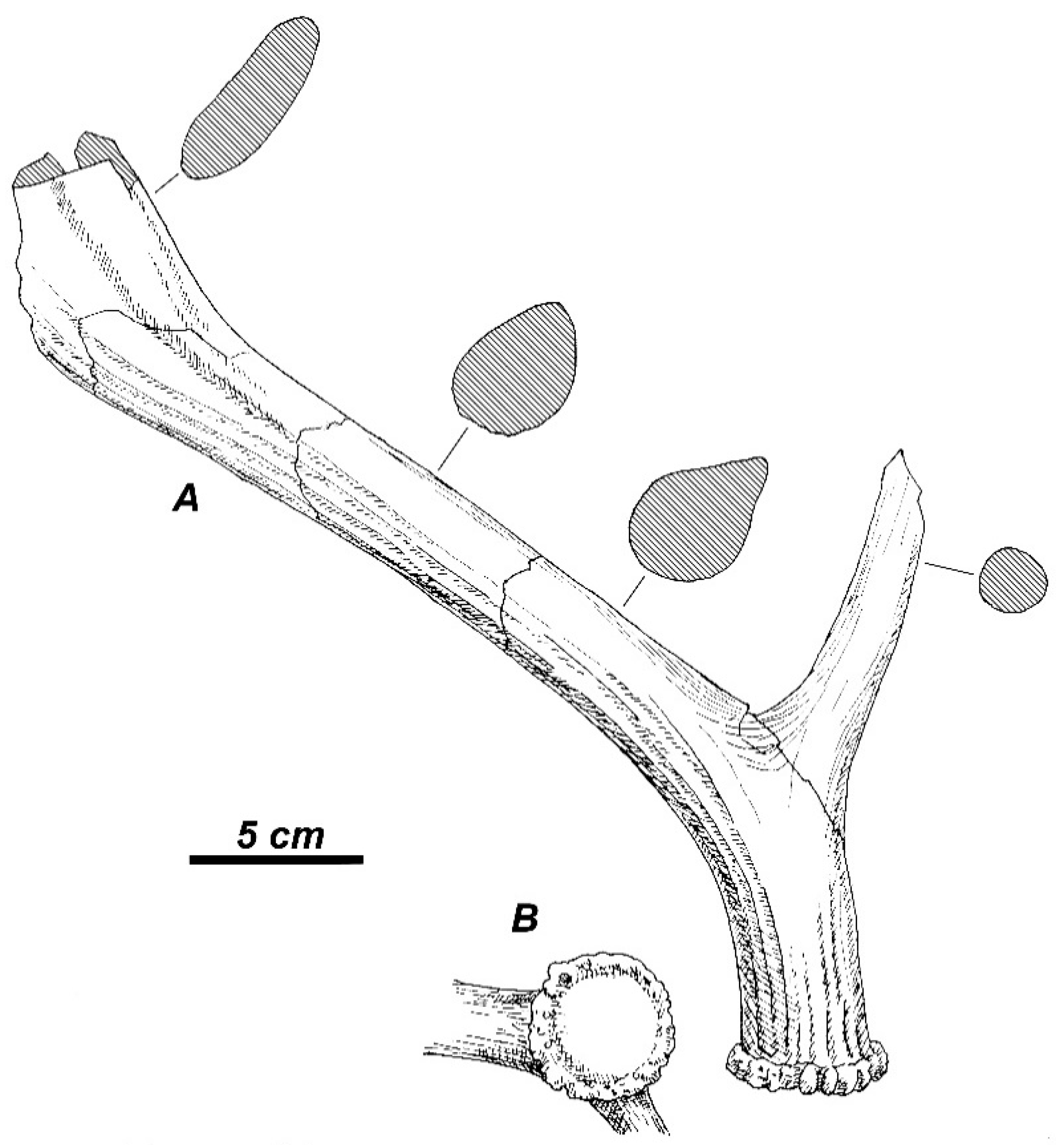
Arvernoceros insolitus Vekua et al., 2010 from Early Pleistocene of Dmanisi (Georgia) is a large-sized deer that is characterized by the very low position of the first tine and the distal palmation, which is developed on the posterior side of the beam and bears small posterior digitations [
67]. The distalmost part of the tine is terminated with a very long anterior tine and another broader and more flattened tine separated from the anterior tine and the posterior palmation by broad and deep notches. The first tine is situated very close to the burr; the middle tine is not present. The very low position of the first tine approaches this deer to
Rucervus simplicidens/
colberti and
R. gigans. Therefore, the South Asian origin of this species is quite probable. Nonetheless, the lack of sufficient data does not allow for the moment to demonstrate the relationships of
A. insolitus with
Rucervus from the Siwaliks and Apollonia-1.
Titov and Shvyreva [
68] described another large-sized deer species
Megaloceros stavropolensis from the Late Villafranchian of Ciscaucasia. This species is based on a large antler with the large fan-shaped distal palmation terminated by strong and long tines on its posterior portion. The first tine is situated at a certain distance from the burr. There are no middle and posterior tines. The cross-section of the beam is irregularly triangular. The antler beam bends backwards from the first ramification and is gently bowed sidewards [
68]. The antler shape generally is very different from that of
A. insolitus and it cannot be regarded as an individual morphological variant of the same species. Titov and Shvyreva [
68] regard this cervid as a transitional form between
R. (
Arvernoceros)
ardei and
M. giganteus, thus following Heintz’s [
25] hypothesized relationship between
Arvernoceros and
Megaloceros. Apparently, this cervid form is of East Asian origin. The general shape of palmation recalls the small-sized
Dama sericus Teilhard de Chardin & Trassaert, 1937 from the Plio-Pleistocene formations of South-Eastern Shansi, China [
44]. Grubb [
69] included
D. sericus in the genus
Arvernoceros, and, in accordance with this viewpoint,
M. stavropolensis was also included in
Arvernoceros [
44]. Nonetheless, it is possible that the Ciscaucasian deer is closely related to the Asian
Sinomegaceros lineage, which, in its turn, could be phylogenetically related to
Rucervus [
44].
4.3. Morphological Similarities and Geographic Distribution of Rucervus radulescui and Rucervus gigans
The available cranial characteristics of
R. radulescui generally correspond to the cranial morphology of modern
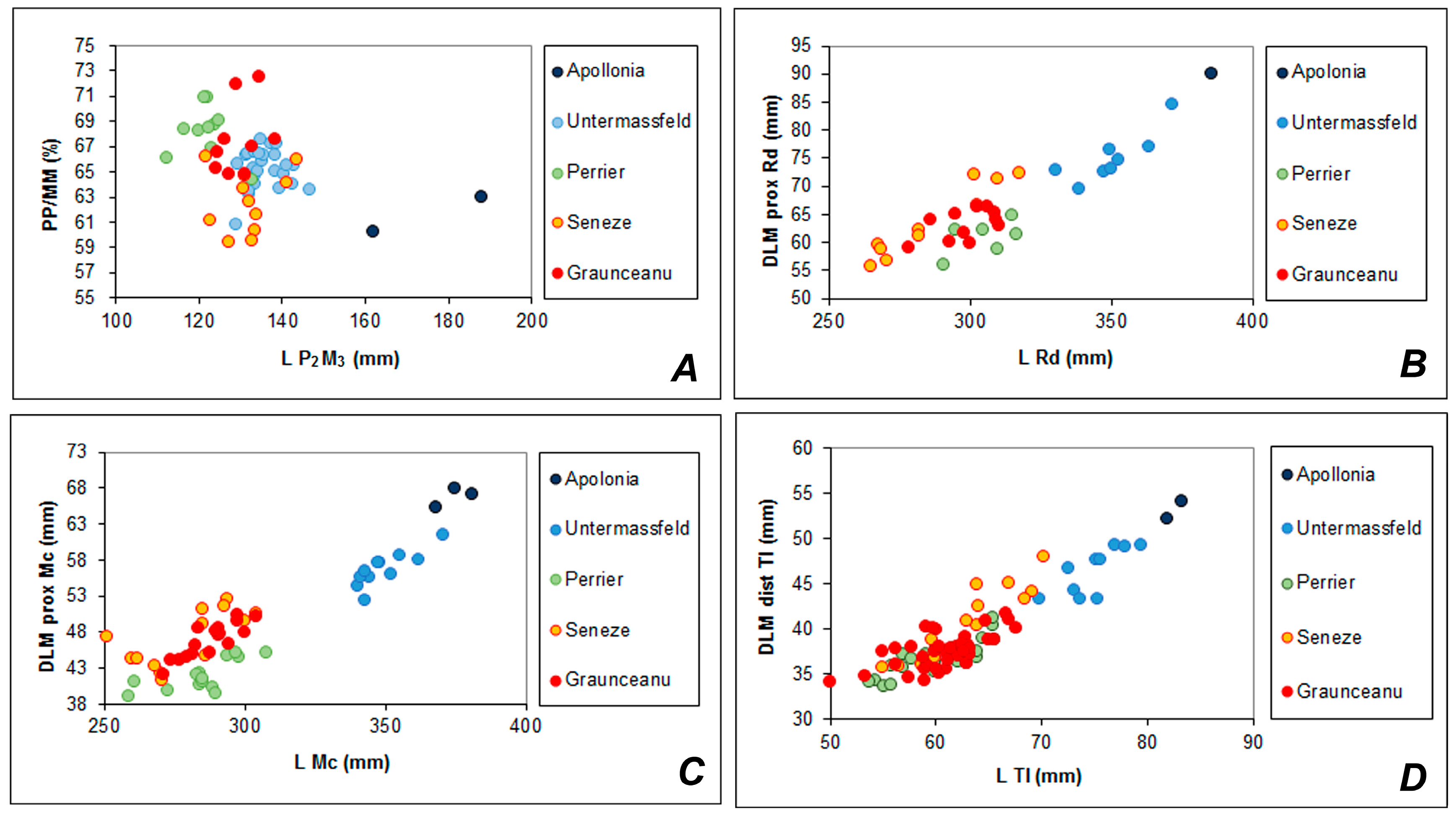
R. duvaucelii: the nasal bones of the Romanian species are rather short and they do not reach the anterior orbital line, the preorbital fossae are large, the ethmoidal openings are equally large with long (ca. 1/2 of the orifice length) contact with the nasal bones, the basioccipital bone is broadened in the pharyngeal tuberosities. The difference with modern species concerns mostly the relatively shorter diastema in
R. radulescui, which apparently is correlated with the comparatively shortened muzzle. The palatal width between the third molars of
R. radulescui (70.7 mm; the female skull from ISER) seems to be somewhat broader than that in
R. simplicidens (the skull No. 19829, AMNH), which amounts to 55 mm (measured from the Figure 145 in Colbert [
36]).
R. radulescui shows the specific antler bauplan characteristic of the genus
Rucervus (the missing middle tine, the posterior insertion of the crown tines that may occasionally be bifurcated in their distal parts). The distal portion of the antler is transformed into the relatively large bilobed palmation, however, the initial
Rucervus-type construction of the antler crown is easily recognisable. The antler specialization is seen in the loss of the accessory prong on the first tine and the cylindrical shape of the first tine. This morphological variant is found in the antler 34590 (NHML) from Perrier-Etouaires, described as
Cervus cylindroceros Dawkins, 1878. The distal portion of
C. cylindroceros is terminated with a simple fork formed by the longer anterior tine, which may be interpreted as the continuation of the main beam, and the shorter posterior tine [
70]. According to Heintz [
25],
C. cylindroceros represents an early ontogenetic stage of antler development of
R. (
Arvernoceros)
ardei.
The antler proportions of R. radulescui permit to group this cervid form with European species R. ardei and R. verestchagini. These species share the relatively high position of the first (basal) tine and the very compact or even poor (as in R. verestchagini) development of the crown part. The crown part (palmation) attains less than 30% of the antler length in the type specimen of R. ardei and apparently the antler from Valea Grăunceanului is characterized by similar proportions. These shared antler characteristics permit to keep Arvernoceros Heintz, 1970 as a subgenus of Rucervus.
The dentition of
R. radulescui remains quite primitive in its relatively long lower premolar series and the low degree of molarization of P
4. The lost cingulum in the upper molars and the more complicated morphology of P
3 and P
4 represent the advanced dental features of
R. radulescui. The dental diagnostic differences between
R. radulescui and
E. ctenoides are quite weak and are clearly visible when a large serial material is analyzed. Generally,
R. radulescui is distinguished from
E. ctenoides by the relatively longer lower premolar series and the stronger development of entoconid and posterior wing of metaconid in P
3 and P
4 with a trend towards the fusion of the posterior premolar elements. It should be noted that
Eucladoceros generally shows a trend towards the stronger development of the anterior wing of the metaconid in P
4 and its occasional fusion with the paraconid [
25,
53].
Unlike the species of the subgenus
Arvernoceros, the South Asian
Rucervus species, which correspondingly are included here in the nominotypical subgenus
Rucervus, are characterized by the very low position of the first tine and the proportionally larger crown part of the antler. The crown part of the antler attains almost ½ of the antler length in
R. duvaucelii [
29,
30,
69], and apparently this is the case for
R. simplicidens/colberti. The crown part of antler in
R. schomburgki is even relatively longer (Figure 3, [
69]).
The complete antlers of
R. gigans are unknown, however, the low position of its first ramification resembles the antler from the Siwaliks figured by Colbert [
36]. The dentition of
R. gigans shares with
R. simplicidens the weak development of cingulum in the upper third molar and the presence of a well-developed flat entostyle. These details of antler and dental morphology suggest that
R. gigans should be included in the subgenus
Rucervus being a peculiar South Asian element of the fauna from Apollonia-1.
4.4. Early Pleistocene Dispersals of Ruminants in Southeastern Europe
The earliest occurrence of
Rucervus in Western Eurasia has been recorded in the Early Pliocene of Weze-1 (Poland) [
62] and Kvabebi (Georgia). This roughly simultaneous occurrence of
Rucervus in Eastern Europe and Transcaucasia suggests the general dispersal event of this genus in Western Eurasia and probably is the starting point of divergence between the subgenera
Rucervus and
Arvernoceros. Apparently, the origin of
R. radulescui is related to the evolutionary branch of
Rucervus evolved in southeastern Europe, since this species shares some details of its antler morphology with other species from the European continent (
R. ardei,
R. verestchagini), and its area of distribution includes the Balkan Peninsula and the North Pontic area. The western border of the distribution area, probably, is shaped by the ecological incompatibility with
E. ctenoides. The West European cervid is characterized by a similar body size and a similar degree of evolutionary specialization. One can assume that those two deer generally depended on similar ecological resources, as it was demonstrated by the study of forage habits of large Early Pleistocene cervids from Western Europe and the Balkans [
71]. The analysis of the dental wear of
E. ctenoides from Chilhac, Sénèze, and Saint-Vallier and that of the cervid from Gerakarou—regarded in the present study as
R. radulescui—shows the similar range of forage resource exploitation extending from browsing to mixed feeding [
71]. However, according to the morphofunctional study of the postcranial skeleton that was carried out by Curran [
5,
6],
R. radulescui and
E. ctenoides were adapted to different types of landscape. The geometric morphometric analysis of limb bones demonstrated that
E. ctenoides from Saint-Vallier and Sénèze inhabited the intermediate open environment, while
R. radulescui was adapted to open and probably mountainous habitats [
5,
6]. Therefore,
R. radulescui may be regarded as an ecological vicar species of
E. ctenoides.
The associated with
R. radulescui fauna is quite archaic with a certain oriental paleobiogeographic influence. The associated with
R. radulescui fauna from Valea Grăunceanului is similar to those from Varshets and Slivnitsa, and it rather represents an archaic Villafranchian association of quite specialized and stenobiont species that existed in Europe before the “wolf event” (sensu Azzaroli [
15]) or
Pachycrocuta faunal turnover [
16]. This is the case of sabertooth cats [
4],
Acinonyx pardinensis [
72], the Villafranchian bovids
Pliotragus,
Gazellospira, and
Procamptoceras, and the small
Axis-like cervid
Metacervocerus rhenanus [
3]. According to Spassov [
7], the wave of dispersals of new bovid forms from the East characterize the early Late Villafranchian Balkan faunas and indicates the progressive aridification of the region. The presence of
Mitilanotherium,
Gazellospira, Ovis, and
Manis gives to the “pre-
Pachycrocuta” Balkan faunas a rather Central and East Asian affinity [
3,
36,
73].
The “post-
Pachycrocuta” Balkan faunas show a greater similarity with Transcaucasian coeval faunas. The fauna from Fântâna lui Mitilan (MN18) and the middle faunal horizon (T-2) of the Irimești Area (Romania) are interesting by the earliest well-dated record of the large-sized deer
Praemegaceros obscurus in Europe. The faunal assemblages from Fântâna lui Mitilan and T-2 of Irimești are correlated with Tasso Faunal Unit and Psekups [
10,
74] and share with the fauna from Dmanisi several cervid species with a quite restricted biogeographic distribution.
P. obscurus is known by its abundant remains from Salcia (Moldova), Pietrafitta (Italy), and Forest Bed Formation (England). Some rare and very fragmentary remains of
P. obscurus are also known from the Tamanian fauna of Semibalki-1 (Russia) [
75]. This is the only species of the genus reported from the Early Pleistocene of the Near East [
75]. The age of
P. obscurus from Ubeidiya (Israel) generally corresponds to that of the findings of this deer from Italy. The remains of
P. obscurus from Dmanisi (Georgia) [
67] are older and they suggest the South Asian origin of this species. It is quite probable that
P. obscurus is phylogenetically close to
Panolia sp. from the Siwaliks [
44].
Cervus nestii and
Alces gallicus are two other cervid species shared by the faunas from Fântâna lui Mitilan and Dmanisi [
73,
76,
77].
C. nestii is closely related to modern
Cervus elaphus but shows much smaller body size and simple antlers lacking the bez tine and terminated with the simple fork [
44]. The fine complete antler of this species from Dmanisi was described by Kahlke [
78] as
Cervus abesalomi [
44,
76]. The distribution of
C. nestii in Europe was rather restricted and included only the Balkans and the Italian Peninsula. This species is regarded here as a warm-loving South Asian faunal element.
Alces gallicus is a side branch of
Alces lineage with quite specialized large antlers, but primitive dentition with unmolarized P
3 and
Palaeomeryx fold in lower molars [
44]. The remains of this species are known from Liventsovka (Russia) and Kuruksai (Tadjikistan) [
34,
79], therefore, the Central Asian origin for this species is suggested. The occurrence of
A. gallicus in Dmanisi is interesting, since it indicates the two-way faunal exchange between Eastern Europe and the Transcaucasian Land. Some poor remains of
Alces ex gr.
minor-gallicus have been also reported from the Late Villafranchian fauna of the Denizli basin (Turkey) [
80]. The faunal exchange between the Balkans and the Anatolian-Caucasian Land probably occurred due to the temporary closing of Bosporus that supposedly took place during this geological period [
7] and it could be triggered by the 41 kyr glacial cycles. Spassov [
7] indicated a presence of a Late Villafranchian paleobiogeographic unity of Southeastern Europe and Southwestern Asia analogous to the Greco-Iranian Late Miocene paleobiogeographic province. Apparently, the Balkan and Caucasian-Anatolian glacial refugia acted as communicating vessels during the cold phases. This is also the case of the Italian glacial refugium, which was biogeographically more open for faunal exchanges with the eastern refugia during the cold phases due to the sea level drop and the deforestation of the Dinaric Alps [
39,
73].
Possibly, the temporary closure of Bosporus during one of the cold phases of 41 kyr glacial cycles also enabled early hominins to disperse into the Balkan Peninsula.
The northern Peri-Pontic path of southern warm loving species dispersals (including
Homo ex gr.
erectus), as hypothesized by Ivanova et al. [
81] and Strait et al. [
9], seems to be less probable. First of all, the Late Villafranchian climate cooling that was triggered by the higher amplitude 41 kyr cycles caused the increase of seasonality, and, for the first time in Europe, the drop of the mean temperature of cold months below freezing point, as it was shown for Central Europe [
82]. It is necessary to note that water obstacles should be practically insurmountable for warm-loving species in the conditions of cold climate. Therefore, the unfavourable climate conditions could impede warm-loving terrestrial mammals to disperse northward from the Caucasian mountain range. The mountain range of the Greater Caucasus protected the Caucasian land from the North and apparently created specific mild climate conditions, as in the case of large European southern peninsulas protected from the north by the Alpine-Himalayan mountain belt [
73]. On the other hand, the Caucasian land at that time was a rather large peninsula that was separated from the West and from the East by Kujalnik (Black) and Akchagyl (Caspian) seas, and by the Manych Straight from the North [
83]. This also explains the refugial character of the area of distribution of the endemic Pliocene holdover deer
Praeelaphus sp. that survived in the fauna from Dmanisi (=
Cervus ex gr.
Arvernoceros ardei fide Vekua [
84]) [
44].
The dispersals of south Asian ruminants toward the Balkan Peninsula continued during the final stage of Late Villafranchian. This is the case of
R. gigans from Greece, which coexists with
P. pliotarandoides, a large-sized deer of supposedly Central Asian origin [
44].
Bison (Eobison) tamanensis/Bison (Eobison) sp. with the larger area of distribution (Greece, Italy, Moldova, Ukraine, and South Russia) is another large Late Villafranchian ruminant that entered southeastern Europe from South Asia. Apparently, this primitive bison is related to B. (Eobison) georgicus from Dmanisi and B. (Eobison) sivalensis from Siwaliks.
The paleobiogeographic origin of the small-sized deer from the Early Pleistocene of the Near East and Anatolia is not clear due to the insufficient data. The antler remains from Ubeidya [
85] and the Denizli basin [
80] remind the European
Metacervocerus rhenanus in their size and the high position of the first ramification. This cervid genus is known from the Siwalik fauna as the rather large species
Metacervocerus punjabiensis, which is characterized by its pronounced mesodonty [
61]. However, it is difficult to estimate the difference in the relative cheek tooth crowns height from the figures that are provided by Geraads [
85] and Brown [
61] first of all because the figured specimens are characterized by a different stage of wear, but if the difference in crown height of those two species will be demonstrated, the direct phyletic relationship between
M. rhenanus and
M. punjabiensis should be ruled out. Both
M. rhenanus and
M. punjabiensis are very close to
Axis shansius from Shansi (China) [
86]. The Chinese species is as large as
M. punjabiensis but is distinguished by brachyodont dentition. The molarized P
4 from Ubeidya figured by Geraads (Pl. II, [
85]) may attest to the presence of the genus
Dama in this fauna.
The problem of small-sized deer from the Near East is complicated by the presence of the
Axis-like
Cervus rewati from the Upper Siwaliks described on the basis of a mandible [
56]. The antler morphology of this species remains unknown.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}