
Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
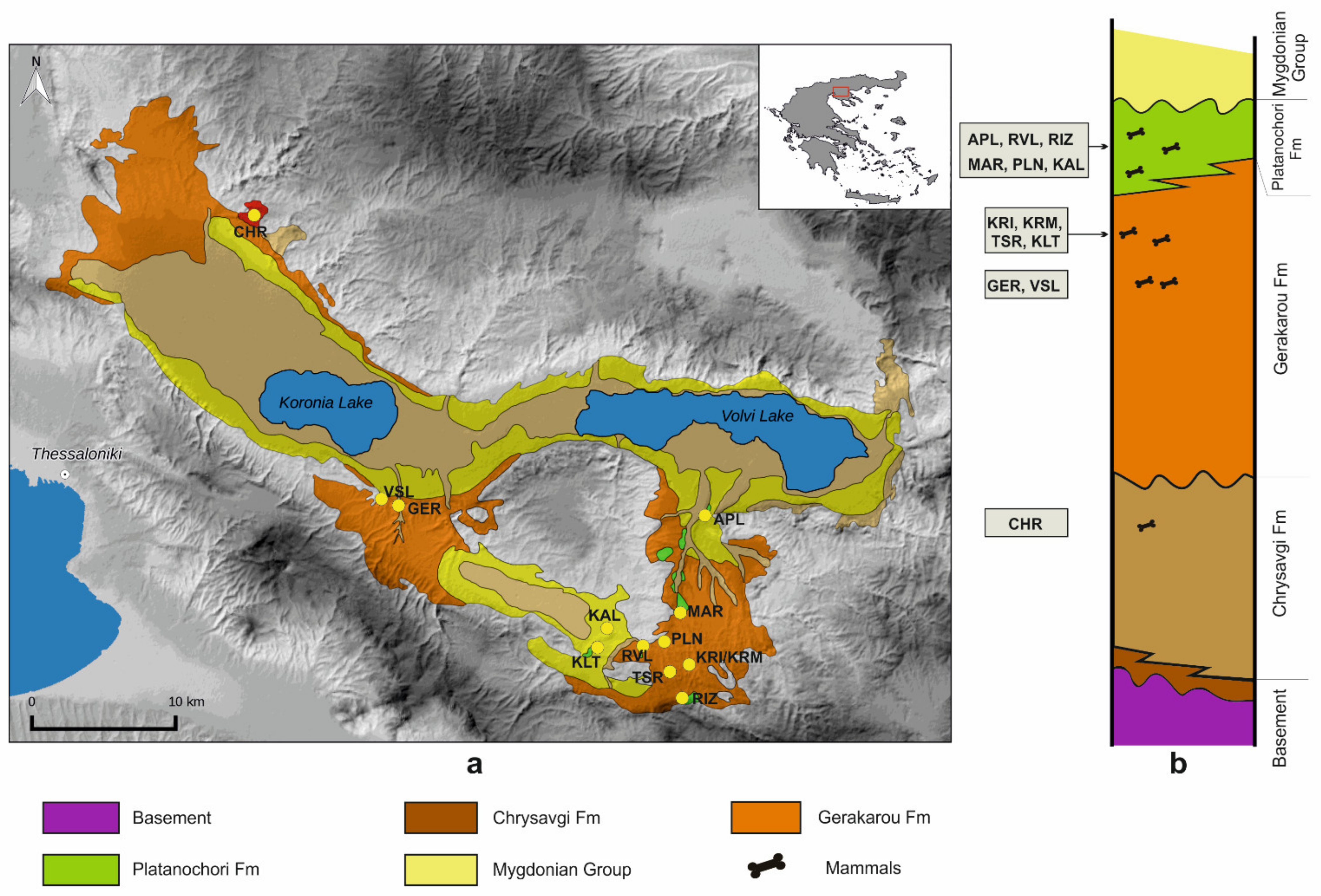
2. Geological Setting
3. Materials and Methods
4. Systematic Palaeontology
4.1. Description
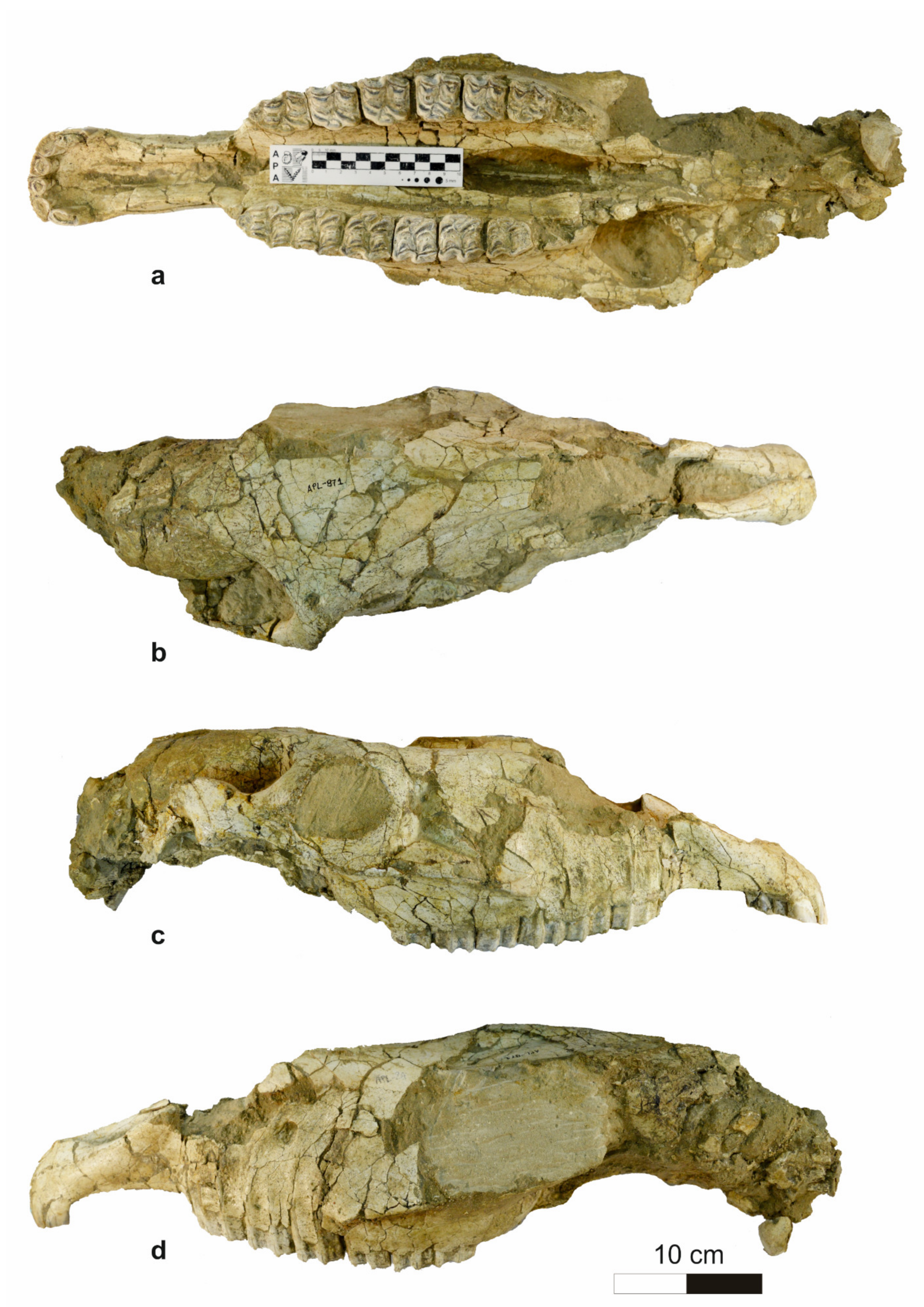
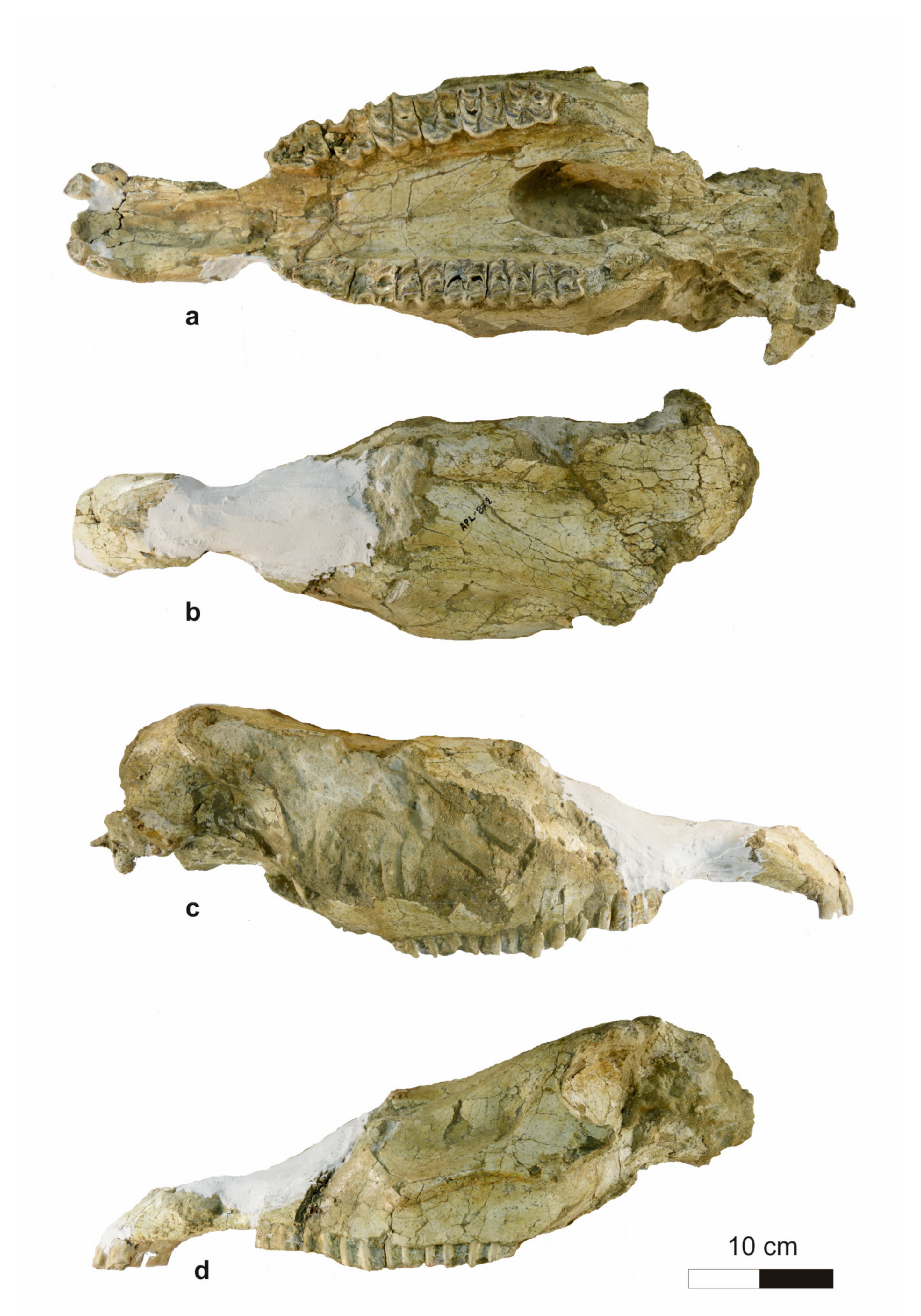
4.1.1. Cranium
4.1.2. Mandible
4.1.3. Upper dentition
4.1.4. Lower Dentition
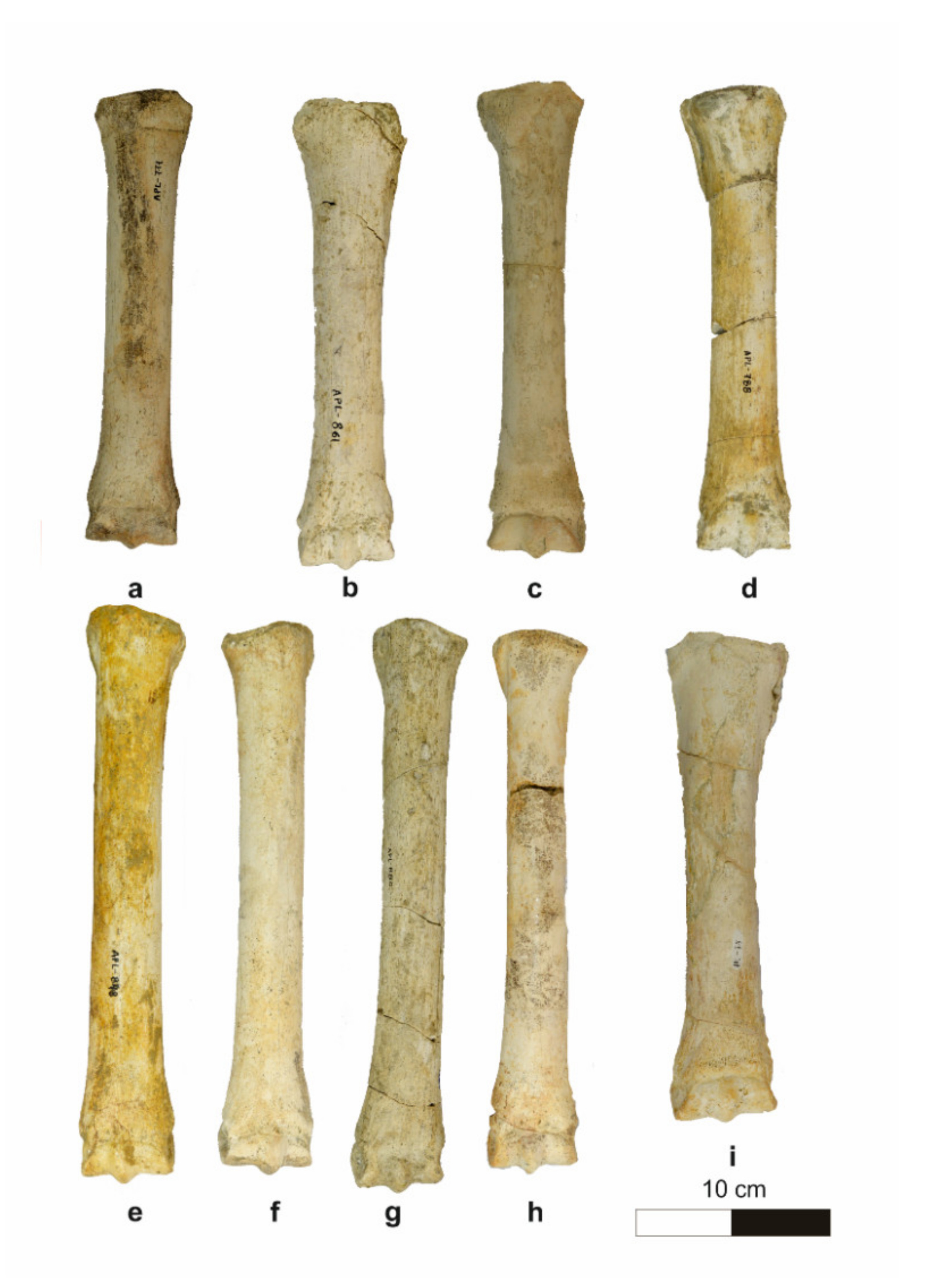
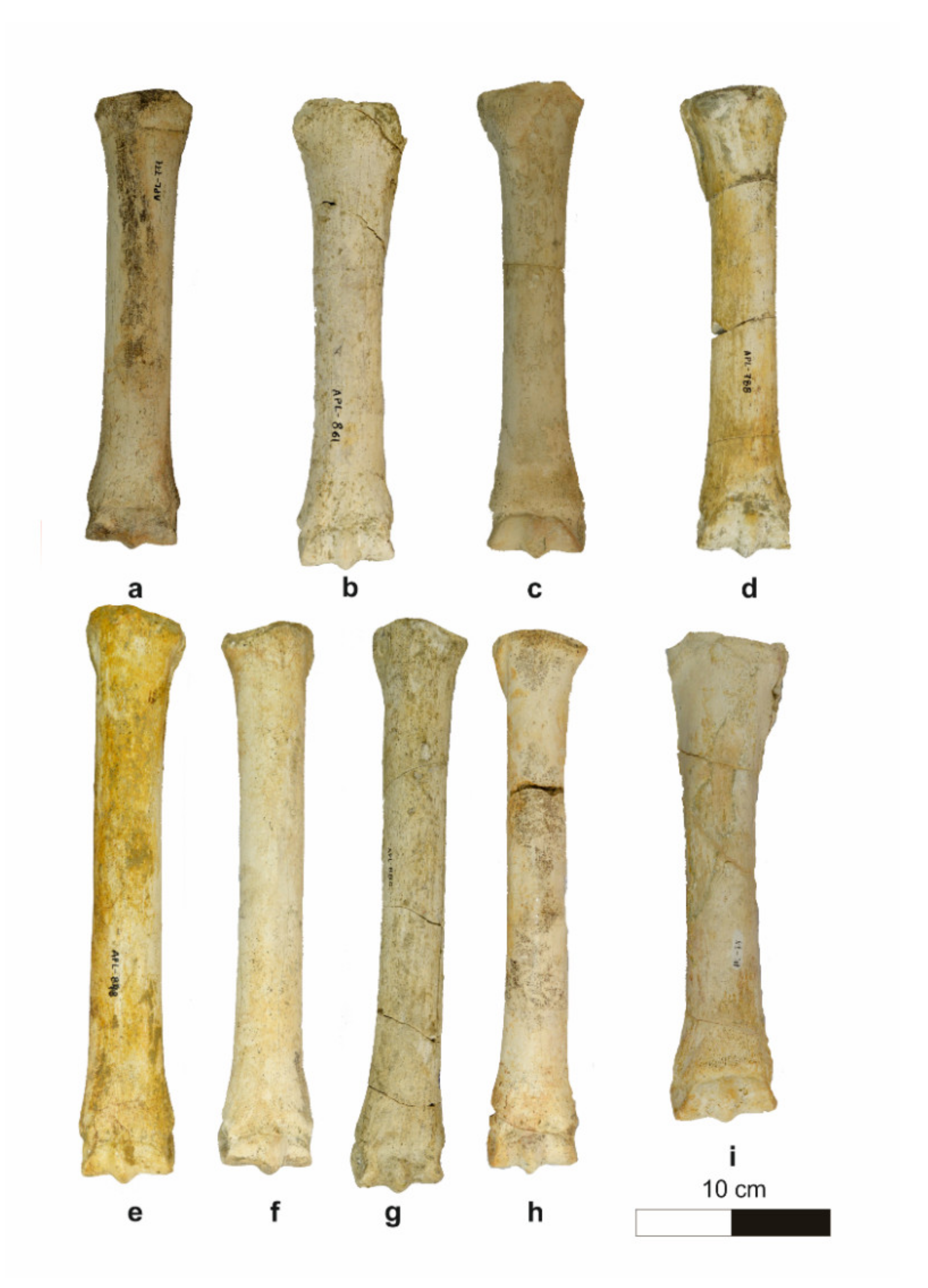
4.1.5. Metapodials
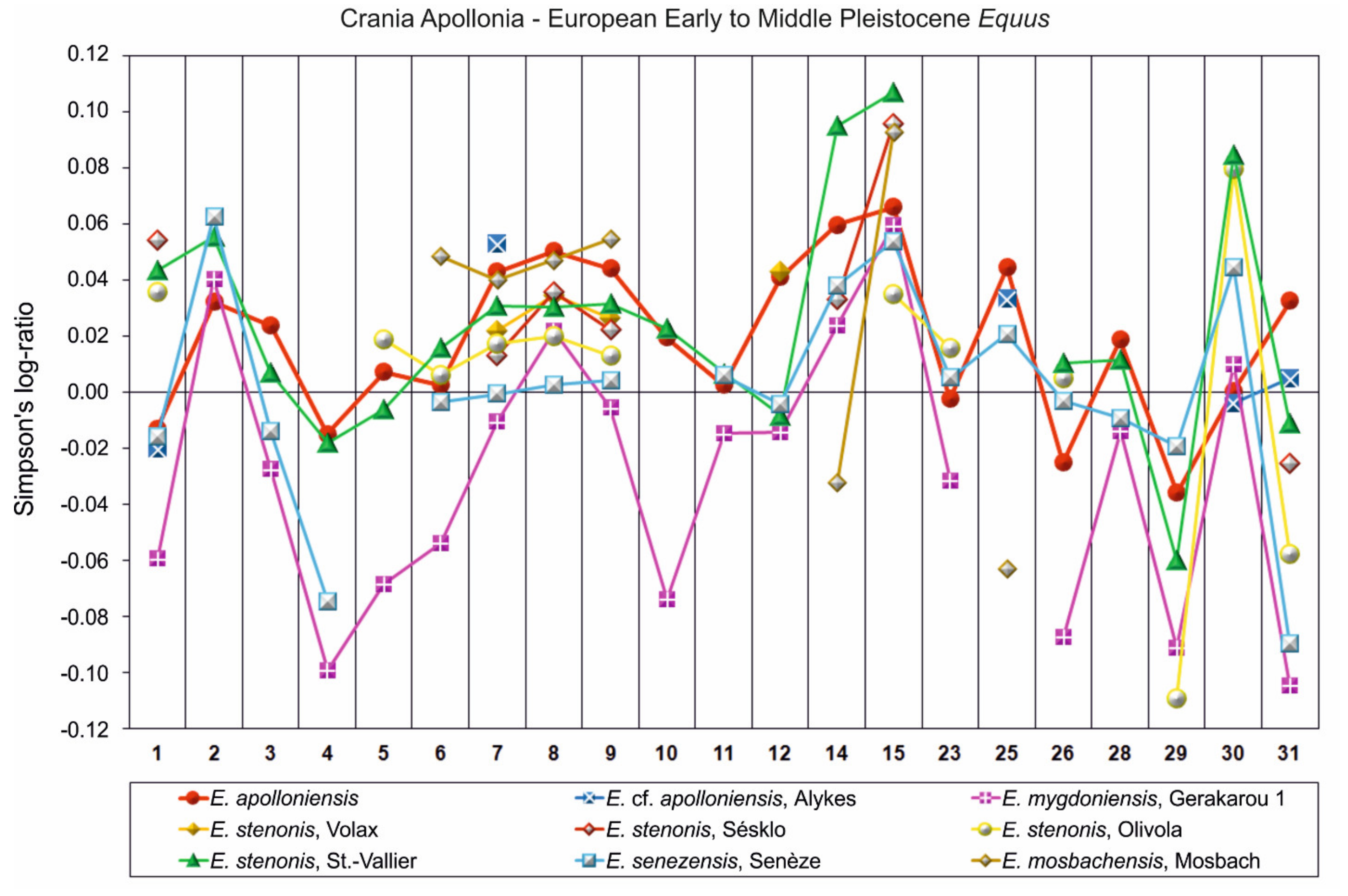
5. Comparisons
6. Discussion
- (a)
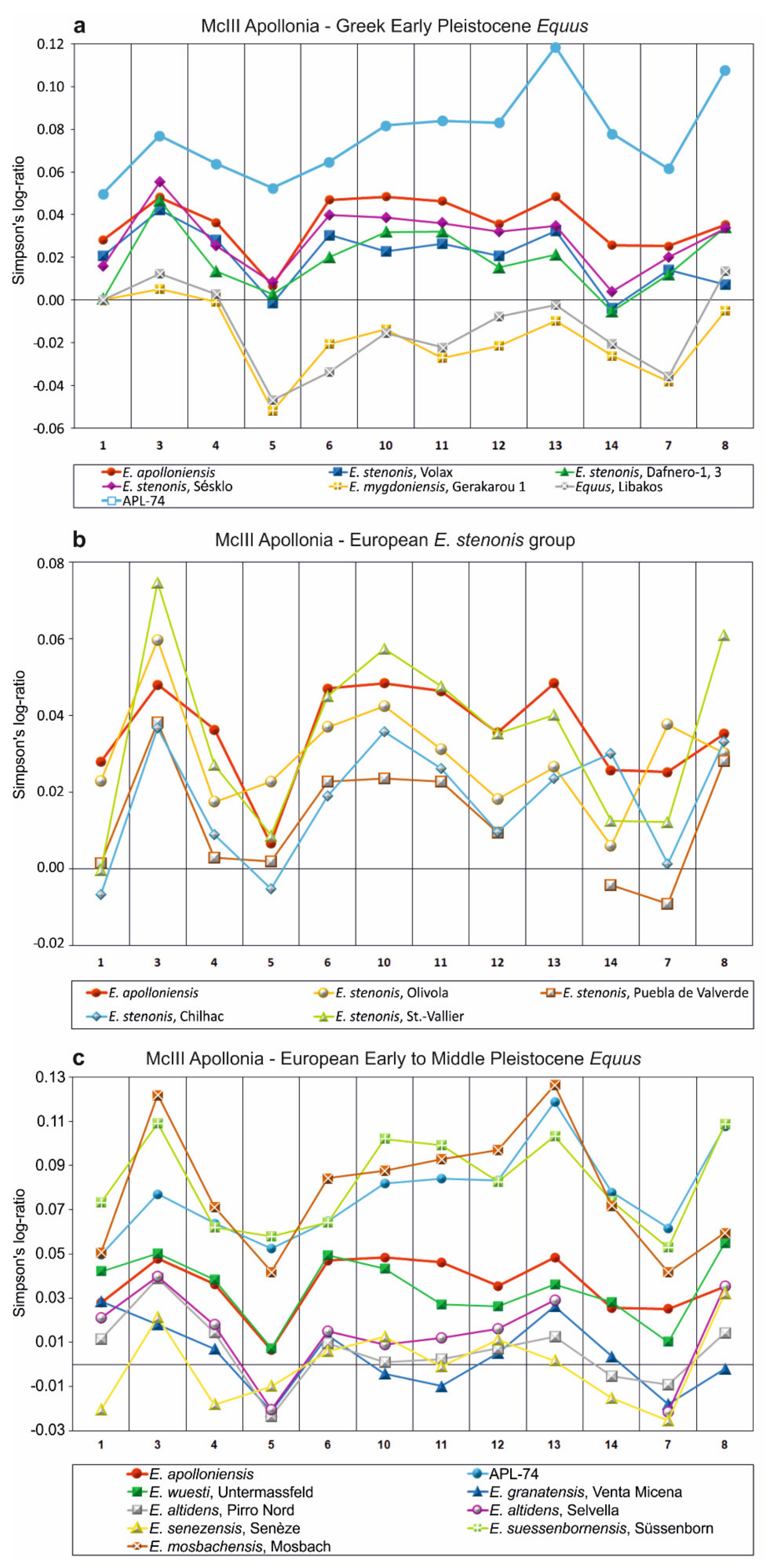
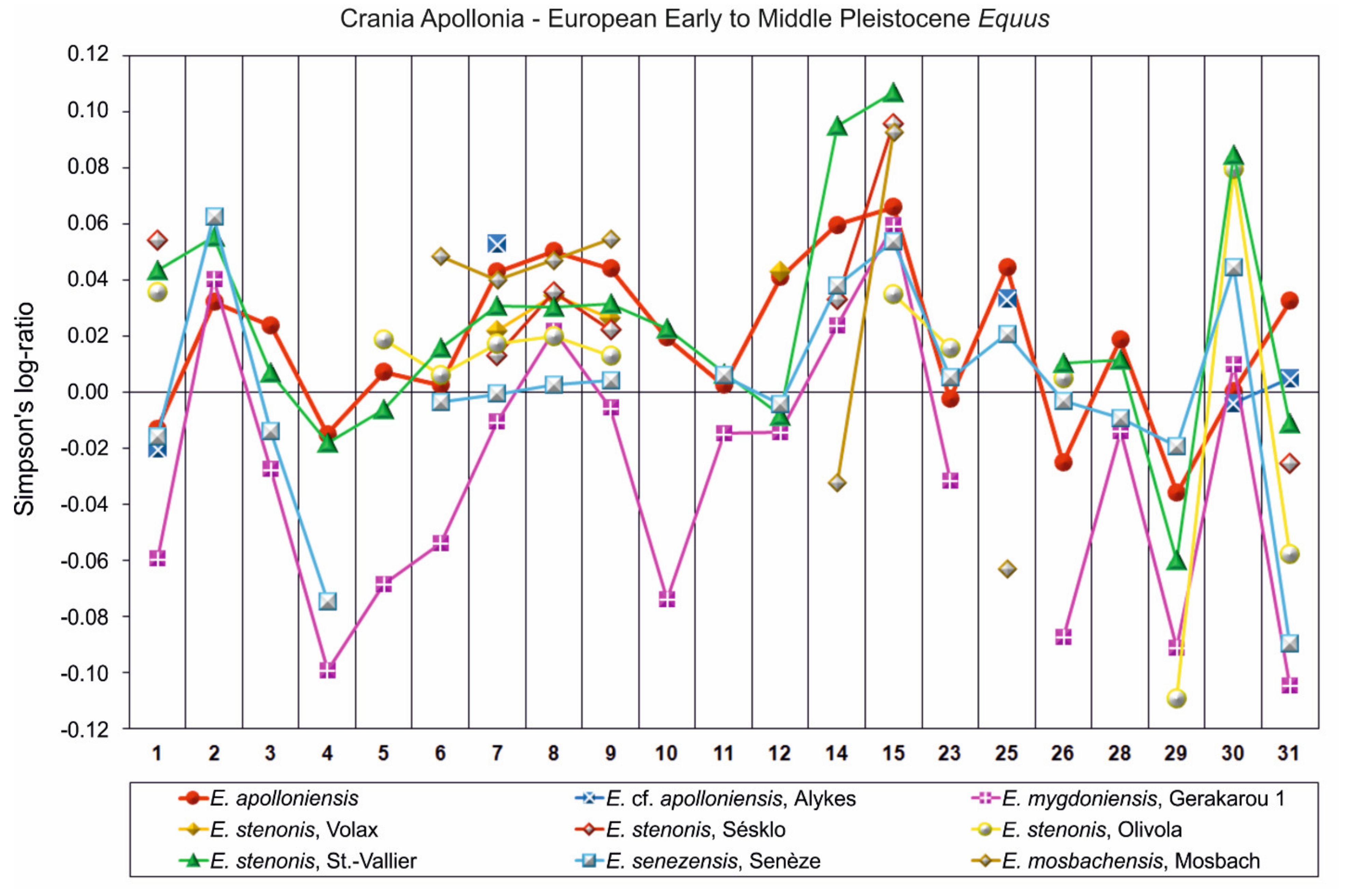
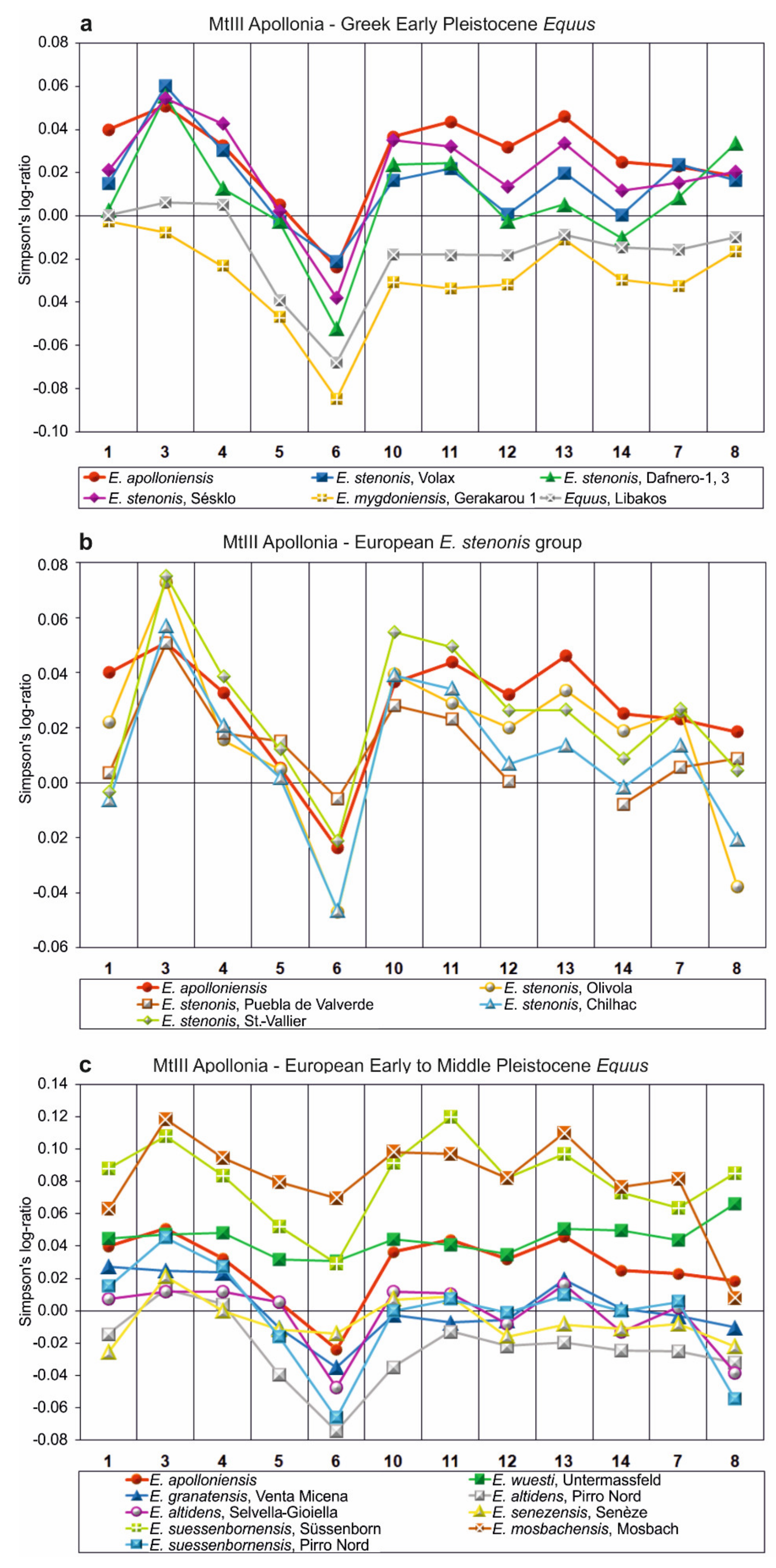
- E. apolloniensis could be an intermediate form between E. stenonis and the middle Pleistocene equid, E. suessenbornensis. Although the general size of the metapodials of E. apolloniensis is similar to those of E. stenonis, the proportions fit better with the slender but smaller E. altidens from Pirro Nord and E. mygdoniensis. In other words, there is an obvious proportional analogy associated with a size increase through time from the metapodials of E. mygdoniensis and E. altidens from Pirro Nord to those of E. apolloniensis (Figure 9) (see also [41]). E. apolloniensis may originate from an E. mygdoniensis–E. altidens stock-keeping similar proportions on the metapodials but increasing in size. Ref. [103] suggested that E. mygdoniensis, E. altidens, and perhaps E. wuesti and E. apolloniensis could represent different local species or ecomorphotypes of the same lineage. Although this scenario cannot be ruled out with the available data, differences on the crania (at least with the better-known E. mygdoniensis) contradict the hypothesis of local ecomorphotypes and rather support the idea of different species from the same lineage. The affinities with E. suessenbornensis at the distal articulations of the metapodials (analogies between meas. 10–11) could also support a link between the two species. This hypothesis rather supports the idea that both species, E. suessenbornensis and possibly E. apolloniensis, evolved in Europe and belonged to stenonoid lineage.
- (b)
- Based on similarities of both upper and especially lower teeth and metapodials, Ref. [61] noted affinities between E. apolloniensis and the modern wild asses (E. africanus). If indeed, as these authors state, E. apolloniensis represents an early step in the lineage of asses, it cannot be linked to stenonoid s. s. and E. suessenbornensis [100]. Although the lower teeth of the mandible APL-570 are ass-like, the revised description of both old and new material suggests that E. apolloniensis resembles asses as much as hemiones.
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Localities: | |
| ALK | Alykes (Thessaly, Greece); |
| APL | Apollonia (Macedonia, Greece); |
| CH | Chilhac (France); |
| CHR | Chrysavgi, (Macedonia, Greece); |
| DFN, DFN3 | Dafnero 1, 3 (Macedonia, Greece); |
| GER | Gerakarou 1 (Macedonia, Greece); |
| KAL, KLT | Kalamoto 1, 2 (Macedonia, Greece); |
| KRI, KRM | Krimni 1, 2 (Macedonia, Greece); |
| LIB | Libakos (Macedonia, Greece); |
| MAR | Marathousa 1 (Macedonia, Greece); |
| PLN | Platanochori (Macedonia, Greece); |
| PN | Pirro Nord; |
| RIZ | Riza 1 (Macedonia, Greece); |
| RVL | Ravin Voulgarakis (Macedonia, Greece); |
| SES | Sésklo (Thessaly, Greece); |
| SU | Süssenborn (Germany); |
| SE | Senèze (France); |
| SG | Selvella-Gioiella; |
| StV | Saint-Vallier (France); |
| TSR | Tsiotra Vryssi (Macedonia, Greece); |
| VOL | Volax (Macedonia, Greece); |
| VSL | Vassiloudi (Macedonia, Greece) |
| Other: | |
| McIII | third metacarpal; |
| MtIII | third metatarsal; |
| DAP | antero-posterior diameter; |
| meas. X | measurement X; |
| PCA | principal component analysis; |
| Fm | Formation |
References
- Melentis, J.K. Die dentition der Pleistozänen proboscidier des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques des Pays Helléniques 1961, 12, 153–262. [Google Scholar]
- Melentis, J.K. Über Equus abeli aus dem Mittelpleistozän des Beckens von Megalopolis im Peloponnes (Griechenland). Proc. Acad. Athens 1963, 38, 507–519. [Google Scholar]
- Melentis, J.K. Die Pleistozänen Nashörner der Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Des. Pays Helléniques 1965, 16, 363–402. [Google Scholar]
- Melentis, J.K. Über Hippopotamus antiquus DESMAREST aus dem Mittelpleistozän des Beckens von Megalopolis in Peloponnes (Griechenland). Ann. Géologiques des Pays Helléniques 1965, 16, 403–435. [Google Scholar]
- Sickenberg, O. Eine säugetierfauna des tieferen Biharius aus dem Becken von Megalopolis (Peloponnes, Griechenland). Ann. Géologiques des Pays Helléniques 1975, 27, 25–73. [Google Scholar]
- Forsyth-Mayor, C.I. Faune mammalogische delle isole di Kos e di Samos (let al prof. B. Meneghini). Atti della Soc. Ital. Sci. Nat. 1887, 5, 272–275. [Google Scholar]
- Brunn, J.H. Contribution a l’etude geologique du Pinde Septenrional et d’une partie de la Macedoine occidentale. Ann. Géologiques des Pays Helléniques 1956, 7, 1–358. [Google Scholar]
- Sickenberg, O. Revision der wirbeltierfauna der höhle Petralona (Griech. Macedonien). Ann. Géologiques des Pays Helléniques 1971, 23, 230–264. [Google Scholar]
- Sickenberg, O. Die unterpleistozäne fauna von Wolkas (Griech.-Mazedonien): I. Eine neue giraffe (Macedonitherium Martinii nov. gen. nov. spec.) aus dem unteren Pleistozän von Griechenland. Ann. Géologiques des Pays Helléniques 1967, 18, 314–330. [Google Scholar]
- Sickenberg, O. Die unterpleistozäne fauna von Wolkas (Griech.-Mazedonien): II. die carnivoren. Ann. Géologiques des Pays Helléniques 1968, 19, 621–646. [Google Scholar]
- Marinos, G. Contribution to the study of the Pleistocene of Mecedonia. Sci. Ann. Fac. Phys. Math. Aristotle Univercity Thessalon. 1964, 9, 95–111. [Google Scholar]
- Paraskevaidis, I. Canis und Equus aus Attika. Neues Jahrb. für Geol. Paläontologie Abh. 1953, 96, 453–496. [Google Scholar]
- Paraskevaidis, I. Saugetierreste aus Griechenland. In Proceedings of the VI Colloquium on the Geology of the Aegean Region, Athens; Kommission bei Springer: Berlin/Heidelberg, Germany, 1977; Volume 3, pp. 1143–1154. [Google Scholar]
- Sakellariou-Mane, H.; Psilovikos, A.; Koufos, G.D. Contribution to the study of Villafranchian in N. Chalkidiki. Sci. Ann. Fac. Phys. Math. Aristotle Univercity Thessalon. 1979, 19, 279–296. [Google Scholar]
- Koufos, G.D. A new late Pleistocene (Würmian) mammal locality from the basin of Drama (northern Greece). Sci. Ann. Fac. Phys. Math. Aristotle Univercity Thessalon. 1981, 21, 129–148. [Google Scholar]
- Symeonidis, N.K.; Tataris, A. The first results of the geological and palaeontological study of the Sesklo basin and its broader environment (eastern Thessaly, Greece). Ann. Géol. Pays Hellén 1983, 31, 146–190. [Google Scholar]
- Koufos, G.D.; Melentis, J.K. New data from the Villafranchian mammal locality of Gerakarou (Macedonia, Greece). Proc. Acad. Athens 1983, 58, 181–185. [Google Scholar]
- Koufos, G.D.; Syrides, G.E.; Koliadimou, K.K. A new Pleistocene mammal locality from Mygdonia basin (Macedonia, Greece). Contribution to the study of Villafranchian (Villanyian) in c. Macedonia. Bull. Geol. Soc. Greece 1988, 23, 113–124. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Koliadimou, K.K. Un nouveau gisement de mammifères dans le Villafranchien de Macédoine occidentale (Grèce). Comptes Rendus Acad. des Sci. Ser. II Sci. la Terre des Planetes 1991, 313, 831–836. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Koliadimou, K.K.; Syridis, G. Apollonia, a new vertebrate site in the Pleistocene of the Mygdonia Basin (Macedonia, Greece)-the first fossil fresh-water mollusks in the area. Comptes Rendus Acad. des Sci. Ser. II Sci. la Terre des Planetes 1992, 315, 1041–1046. [Google Scholar]
- Koufos, G.D. The Pleistocene carnivores of the Mygdonia basin (Macedonia, Greece). Ann. Paléontologie 1992, 78, 205–257. [Google Scholar]
- Koufos, G.D. Early Pleistocene equids from Mygdonia basin (Macedonia, Greece). Palaeontogr. Ital. 1992, 79, 168–199. [Google Scholar]
- Kostopoulos, D.S. The Plio–Pleistocene Artiodactyls of Macedonia (N. Greece). Systematics, Palaeoecology, Biochronology, Biostratigraphy. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1996. [Google Scholar]
- Kostopoulos, D.S. The Plio-Pleistocene artiodactyls (Vertebrata, Mammalia) of Macedonia 1. The fossiliferous site “Apollonia-1”, Mygdonia basin of Greece. Geodiversitas 1997, 19, 845–875. [Google Scholar]
- Kostopoulos, D.S. The Plio-Pleistocene artiodactyls from Macedonia, Greece: 2. The fossilirerous locality of Volakas, VOL (Volakas Basin, Drama, NE Greece). Paleontol. I Evol. 1997, 30–31, 83–92. [Google Scholar]
- Kostopoulos, D.S.; Koufos, G.D. The Plio-Pleistocene artiodactyls of Macedonia (northern Greece) and their biostratigraphic significance; preliminary report. Comptes Rendus-Acad. des Sci. Ser. II Sci. la Terre des Planetes 1994, 318, 1267–1272. [Google Scholar]
- Athanassiou, A. Contribution to the Study of the Fossil Mammals of Thessaly. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 1998. [Google Scholar]
- Athanassiou, A. New data on the Equus stenonis Cocchi, 1867 from the late Pliocene locality of Sésklo (Thessaly, Greece). Geodiversitas 2001, 23, 439–469. [Google Scholar]
- Athanassiou, A. A Villafranchian Hipparion-bearing mammal fauna from Sésklo (E. Thessaly, Greece): Implications for the question of Hipparion–Equus sympatry in Europe. Quaternary 2018, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Maniakas, I. Contribution to the Study of Chrono-Spatial Distribution of Palaeocological Adaptations of European Pleistocene Bovini Based on Ecomorphological Analyses and Geometric Morphometrics. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2019. [Google Scholar]
- Konidaris, G.E.; Tourloukis, V.; Kostopoulos, D.S.; Thompson, N.; Giusti, D.; Michailidis, D.; Koufos, G.D.; Harvati, K. Two new vertebrate localities from the Early Pleistocene of Mygdonia Basin (Macedonia, Greece): Preliminary results. Compt. Rendus Palevol. 2015, 14, 353–362. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Kostopoulos, D.S.; Koufos, G.D. Mammuthus meridionalis (Nesti, 1825) from Apollonia-1 (Mygdonia basin, northern Greece) and its importance within the Early Pleistocene mammoth evolution in Europe. Geodiversitas 2020, 42, 69–91. [Google Scholar] [CrossRef]
- Koufos, G.D. Late Pliocene carnivores from western Macedonia (Greece). Paläontologische Z. 1993, 67, 357–376. [Google Scholar] [CrossRef]
- Steensma, K.J. Plio-Pleistozäne Großsäugetiere (Mammalia) Aus Dem Becken von Kastoria/Grevena, Südlich von Neapolis-NW Griechenland. Ph.D. Thesis, Technische Universität Clausthal, Clausthal, Germany, 1988. [Google Scholar]
- Tsoukala, E. Contribution to the Study of the Pleistocene Fauna of large Mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona Cave, Chalkidiki (N. Greece). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1989. [Google Scholar]
- Tsoukala, E.; Chatzopoulou, K. A new Early Pleistocene (Latest Villafranchian) site with mammals in Kalamoto (Mygdonia basin, Macedonia, Greece)-preliminary report. Mitt. Komm. Quartärforsch. Österr. Akad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Koufos, G.D.; Vlachou, T. Equus stenonis from the middle Villafranchian locality of Volax (Madeconia, Greece). Geodiversitas 1997, 19, 641–657. [Google Scholar]
- Koliadimou, K.K. Palaeontological and Biostratigraphical Study of the Neogene/Quaternary Micromammals from Mygdonia Basin. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1996. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S. A stenonoid horse (Equidae, Mammalia) from the Villafrnachian of western Macedonia (Greece). Bull. Geol. Soc. Greece 1993, 28, 131–143. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S. New carnivore material from the Plio-Pleistocene of Macedonia (Greece) with a description of a new canid. Münchner Geowiss. Abh. 1997, 34, 33–63. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, Macedonia, Greece. Paleontol. i Evol. 1997, 30–31, 49–76. [Google Scholar]
- Koufos, G.D.; Konidaris, G.E.; Harvati, K. Revisiting Ursus etruscus (Carnivora, Mammalia) from the Early Pleistocene of Greece with description of new material. Quat. Int. 2018, 497, 222–239. [Google Scholar] [CrossRef]
- Symeonidis, N.K. Fossil mammals of Lower Pleistocene (Villafranchian) age from the Sesklo basin (Volos). Ann. Géologiques Des. Pays Helléniques 1992, 35, 1–21. [Google Scholar]
- Azzaroli, A. Quaternary mammals and the “end-Villafranchian” dispersal event-a turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 44, 117–139. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Cerdeño, E.; López Martínez, N.; Morales, J.; Soria, M.D. La fauna Villafranquiense de El Rincón-1 (Albacete, Castilla-La Mancha). Estud. Geológicos 1997, 53, 69–93. [Google Scholar] [CrossRef]
- Bernor, R.L.; Cirilli, O.; Jukar, A.M.; Potts, R.; Buskianidze, M.; Rook, L. Evolution of early Equus in Italy, Georgia, the Indian subcontinent, east Africa, and the origins of African zebras. Front. Ecol. Evol. 2019, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Rook, L.; Bernor, R.L.; Avilla, L.S.; Cirilli, O.; Flynn, L.; Jukar, A.; Sanders, W.; Scott, E.; Wang, X. Mammal biochronology (land mammal ages) around the world from late Miocene to middle Pleistocene and major events in horse evolutionary history. Front. Ecol. Evol. 2019, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Cirilli, O.; Bernor, R.L.; Rook, L. New insights on the Early Pleistocene equids from Roca-Neyra (France, central Europe): Implications for the Hipparion LAD and the Equus FAD in Europe. J. Paleontol. 2020, 1–20. [Google Scholar] [CrossRef]
- Eisenmann, V.; Brunet, M. Présence simultanée de cheval et d’hipparion dans le Villafranchien moyen de France à Roccaneyra (Puy-de-Dôme); etude critique de cas semblables; Europe et Proche-Orient). 1973. Available online: https://vera-eisenmann.com/IMG/pdf/4.Roccaneyra_OK.pdf (accessed on 1 April 2021).
- de Bruijn, H.; Dawson, M.R.; Mein, P. Upper Pliocene rodentia, lagomorpha and insectivora (Mammalia) from the isle of Rhodes (Greece). Proc. Kon. Nederl. Akad. Wetten 1970, 73, 535–584. [Google Scholar]
- Benda, L.; Meulenkamp, J.E.; Van de Weerd, A. Biostratigraphic correlations in the eastern Mediterranean Neogene. Newsl. Stratigr. 1977, 6, 117–130. [Google Scholar] [CrossRef]
- van der Meulen, A.J.; Van Kolfschoten, T. Review of the Late Turolian to Early Biharian mammal faunas from Greece and Turkey. Mem Della Soc. Geol. Ital. 1986, 31, 201–211. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S. The Plio-Pleistocene large mammal record of Greece: Implications for early human dispersals into Europe. In Paleoanthropology of the Balkans and Anatolia: Human Evolution and Its Context; Harvati, K., Roksandic, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 269–280. [Google Scholar] [CrossRef]
- Athanassiou, A. Neogene and Quaternary mammal faunas of Thessaly. Ann. Géologiques des Pays Helléniques 2002, 39, 279–293. [Google Scholar]
- Koufos, G.D.; Syrides, G.E.; Kostopoulos, D.S.; Koliadimou, K.K. Preliminary results about thestratigraphy and the palaeoenvironment of Mygdonia Basin, Macedonia, Greece. Geobios 1995, 28, 243–249. [Google Scholar] [CrossRef]
- Psilovikos, A. Paleogeographic Development of the Basin and Lake of Mygdonia (Lagada-Volvi Area, Greece). Ph.D Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1977. [Google Scholar]
- Koufos, G.D. New material and revision of the carnivora, mammalia from the lower Pleistocene locality Apollonia 1, Greece. Quaternary 2018, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in south-eastern Europe and the megafaunal migratory waves from the east to Europe. Rev. Paleobiol. 2003, 22, 197–229. [Google Scholar]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Musil, R. Die equiden-reste aus dem Unterpleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld Bei Meiningen (Thüringen). Teil 2, Monographien Des. RGZM; Kahlke, R.D., Ed.; Verl. d. Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2001; pp. 557–587. [Google Scholar]
- Eisenmann, V.; Boulbes, N. New results on equids from the Early Pleistocene site of Untermassfeld. In The Pleictocene of Untermassfeld Near Meiningen (Thüringen, Germany), Part. 4, Monographien Des. RGZM; Kahlke, R.-D., Ed.; Monographi; Römisch-Germanischen Zentralmuseums: Weimar, Germany, 2020; Volume 40, pp. 1295–1321. [Google Scholar]
- Eisenmann, V. Mosbach et E. mosbachensis. 2008. Available online: https://vera-eisenmann.com/-mosbach-et-e-mosbachensis- (accessed on 15 December 2020).
- Eisenmann, V. Les chevaux Equus (sensu lato) fossiles et actuels: Crânes et dents jugales supérieures. Cah Paléontologie 1980, 19, 1–186. [Google Scholar]
- Eisenmann, V. Venta Micena et Equus granatensis. 2011. Available online: https://vera-eisenmann.com/-venta-micena-et-equus-granatensis- (accessed on 10 December 2020).
- Alberdi, M.T.; Palombo, M.R. The Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 2013, 288, 25–44. [Google Scholar] [CrossRef]
- Gkeme, A.G. Study of the Pleiocene Equids of the Eltgen Collection (Localities Livakos, Kapetanios, Polylakkos, W. Macedonia). Master’s Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2016. [Google Scholar]
- Gkeme, A.G.; Koufos, G.D.; Kostopoulos, D.S. The Early Pleistocene stenonoid horse from Libakos and Polylakkos (Western Macedonia, Greece): Biochronological and palaeoecological implications and dispersal events. In Proceedings of the Abstracts of 15th Congress of the Regional Committee on Mediterranean Neogene Startigraphy: Exploring a ‘Physical Laboratory: The Mediterranean Basin’, Athens, Greece, 3–6 September 2017; p. 67. [Google Scholar]
- Simpson, G.G.; Roe, A. Quantitative Zoology: Numerical Concepts and Methods in the Study of Recent and Fossil Animals; McGraw-Hill Book Company: New York, NY, USA, 1939. [Google Scholar] [CrossRef]
- Eisenmann, V. Equus Grevyi. 2008. Available online: https://vera-eisenmann.com/ (accessed on 10 December 2020).
- Hammer, O. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Eisenmann, V. Double Knots of Lower Cheek Teeth. 2017. Available online: https://vera-eisenmann.com/double-knots-of-lower-cheek-teeth (accessed on 15 January 2021).
- Eisenmann, V. Family Equidae. In Koobi Fora Research Project: Vol. 2. The Fossil Ungulates: Proboscidea, Perissodactyla, and Suidae; Harris, J.M., Ed.; Clarendon Press: Oxford, UK, 1983; pp. 156–214. [Google Scholar]
- Eisenmann, V.; Alberdi, M.T.; De Giuli, C.; Staesche, U. Methodology. In Studying Fossil Horses, Collected Papers after the New York International Hipparion Conference, 1981; Leiden: New York, NY, USA, 1988; pp. 1–71. [Google Scholar]
- Smuts, M.M.S.; Penzhorn, B.L. Descriptions of anatomical differences between skulls and mandibles of Equus zebra and E. burchelli from southern Africa. Afr. Zool. 1988, 23, 328–336. [Google Scholar]
- Azzaroli, A. The two Villafranchian horses of the Upper Valdarno. Palaeontogr. Ital. 1965, 59, 1–12. [Google Scholar]
- Eisenmann, V. The Senèze equids. 2017. Available online: https://veraeisenmann.com/the-seneze-equids-text (accessed on 20 December 2020).
- Azzaroli, A. The genus Equus in Europe. In European Neogene Mammal. Chronology, NATO ASI Series, Series A: Life Sciences, Vol. 180. NATO ASI S.; Lindsay, E.H., Fahlbusch, V., Mein, P., Eds.; Springer: New York, NY, USA, 1990; pp. 339–356. [Google Scholar] [CrossRef]
- Eisenmann, V.; Baylac, M. Extant and fossil Equus (Mammalia, Perissodactyla) skulls: A morphometric definition of the subgenus Equus. Zool. Scr. 2000, 29, 89–100. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Ortiz-Jaureguizar, E.; Prado, J.L. A quantitative review of European stenonoid horses. J. Paleontol. 1998, 72, 371–387. [Google Scholar] [CrossRef]
- Eisenmann, V. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). Compt. Rendus Biol. 2010, 333, 235–240. [Google Scholar] [CrossRef]
- Eisenmann, V. Equus granatensis of Venta Micena and evidence for primitive non-stenonid horses in the Lower Pleistocene. In The Hominids and Their Environment during the Lower and Middle Pleistocene of Eurasia, Proceedings of the International Conference of Human Palaeontology, Orce, Spain; Gilbert, J., Sanchez, F., Gilbert, L., Ribot, F., Eds.; Museo de Prehistoria y Paleontología: Calle Tiendas, Spain, 1999; pp. 175–189. [Google Scholar]
- Lindsay, E.H.; Opdyke, N.D.; Johnson, N.M. Pliocene dispersal of the horse Equus and Late Cenozoic mammalian dispersal events. Nature 1980, 287, 135–138. [Google Scholar] [CrossRef]
- Azzaroli, A. Ascent and decline of monodactyl equids a case for prehistoric overkill. Ann. Zool. Fennici. 1992, 28, 151–153. [Google Scholar]
- Azzaroli, A. Notes on some Middle and Late Pleistocene equids of Italy. Boll. Soc. Paleontol. Ital. 1999, 38, 97–108. [Google Scholar]
- Alberdi, M.T.; Caloi, L.; Palombo, M.R. The Quaternary fauna of Venosa: Equidae. Bull. Mus. d’Anthropology Préhistoric Monaco 1988, 31, 5–39. [Google Scholar]
- Alberdi, M.T.; Ortiz Jaureguizar, E.; Prado, J.L. Comments on “Equus species as stratigraphic markers. Reality or wishful thinking?” and “A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms” by A. Forsten. Quat. Sci. Rev. 2001, 20, 1149–1151. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Bernor, R.L.; Scott, R.S.; Franzen, J.L.; Solounias, N. New interpretations of the systematics and palaeoecology of the Dorn-Dόrkheim 1 hipparions (Late Miocene, Turolian age [MN11]), Rheinhessen, Germany. Senckenbergiana Lethaea 2003, 83, 103–133. [Google Scholar] [CrossRef]
- Musil, R. Die Pferde aus der oberpliozänen Spaltenfüllung Schernfeld bei Eichstätt. Mitt. der Bayer Staatssamml. Palδontologie Hist. Geol. 1992, 32, 115–162. [Google Scholar]
- Eisenmann, V. Origins, dispersals, and migrations of Equus (Mammalia, Perissodactyla). Cour Forsch. Senckenb 1992, 153, 161–170. [Google Scholar]
- Eisenmann, V. Equus: An evolution without lineages? In Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations (Weimar: Terra Nostra), 18th International Senckenberg Conference 2004, VI International Palaeontological Colloquium in Weimar, Weimar, Germany, 25–30 April 2004; Alfred-Wegener-Stiftung: Berlin, Germany, 2004; pp. 101–104. [Google Scholar]
- Caloi, L. New forms of equids in western Europe and palaeoenvironmental changes. Geobios 1997, 30, 267–284. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, M.R. Osservazioni sugli equidi Italiani del Pleistocene medio inferiore. Geol. Rom. 1987, 26, 187–221. [Google Scholar]
- Barrena, P.P.; Guerrero-Alba, S. Estudio morfométrico del caballo de Venta Micena (Orce, Granada) y su comparación con los équidos modernos y del Plio-Pleistoceno en el viejo y nuevo mundo. Paleontol. I Evol. 1997, 30, 93–148. [Google Scholar]
- Forstén, A. A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms. Quat. Sci. Rev. 1999, 18, 1373–1408. [Google Scholar] [CrossRef]
- Palombo, M.R.; Alberdi, M.T.; Bellucci, L.; Sardella, R. An intriguing middle-sized horse from Coste San Giacomo (Anagni Basin, central Italy). Quat. Res. 2017, 87, 347–362. [Google Scholar] [CrossRef]
- Palombo, M.R.; Alberdi, M.T. Light and shadows in the evolution of South European stenonoid horses. Foss Impr. 2017, 73, 115–140. [Google Scholar] [CrossRef]
- van der Made, J.; Rosell, J.; Blasco, R. Faunas from Atapuerca at the Early–Middle Pleistocene limit: The ungulates from level TD8 in the context of climatic change. Quat. Int. 2017, 433, 296–346. [Google Scholar] [CrossRef] [Green Version]
- Azzaroli, A. On a Late Pleistocene ass from Tuscany, with notes on the history of asses. Palaeontogr. Ital. Pisa. 1979, 71, 27–47. [Google Scholar]
- Eisenmann, V.; Kuznetsova, T. Early Pleistocene equids (Mammalia, Perissodactyla) of Nalaikha, Mongolia, and the emergence of modern Equus Linnaeus, 1758. Geodiversitas 2004, 26, 535–561. [Google Scholar]
- Boulbes, N.; Van Asperen, E.N. Biostratigraphy and palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Eisenmann, V. Discriminating Equus skulls: The Franck’s index and the new palatal index. In Equids in Time and Space: Papers in Honour of Véra Eisenmann, Proceedings of the Conference of the International Council of Archaeozoology, Durham, UK, 23–28 August 2002; Albarella, U., Dobney, K., Rowley-Conwy, P., Eds.; Oxbow Books: Oxford, UK, 2006; pp. 172–182. [Google Scholar]
- Maul, L.C.; Rekovets, L.; Heinrich, W.-D.; Keller, T.; Storch, G. Arvicola mosbachensis (Schmidtgen 1911) of Mosbach 2, A basic sample for the early evolution of the genus and a reference for further biostratigraphical studies. Senckenbergiana Lethaea 2000, 80, 129–147. [Google Scholar] [CrossRef]
- Palombo, M.R. Large mammals faunal dynamics in southwestern Europe during the late Early Pleistocene: Implications for the biochronological assessment and correlation of mammalian faunas. Alp. Mediterr. Quat. 2016, 29, 143–168. [Google Scholar]
- de Menocal, P.B. African climate change and faunal evolution during the Pliocene–Pleistocene. Earth Planet. Sci. Lett. 2004, 220, 3–24. [Google Scholar] [CrossRef]
- Alberdi, M.T. Estudio de los caballos de los yacimientos de Fuente Nueva-3 y Barranco León-5 (Granada). In Ocupaciones Humanas En El Pleistoceno Inferior y Medio de La Cuenca de Guadix-Baza; Toro, I., Martínez-Navarro, B., Agustí, J., Monografías, A., de Andalucía, J., de Cultura, C., Eds.; Editorial Junta de Andalucia: Sevilla, Spain, 2010; pp. 221–306. [Google Scholar]
- Eisenmann, V. The primitive horses of the Vatera formation (Lesvos). Ann. Géologiques des Pays Helléniques 2002, 39, 131–153. [Google Scholar]
- Konidaris, G.E.; Kostopoulos, D.S.; Maron, M.; Schaller, M.; Ehlers, T.A.; Aidona, E.; Marini, M.; Tourloukis, V.; Muttoni, G.; Koufos, G.D. Dating of the Lower Pleistocene vertebrate site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, magnetostratigraphy, and cosmogenic radionuclides. Quaternary 2021, 4, 1. [Google Scholar] [CrossRef]
- Boulbes, N.; Mayda, S.; Titov, V.V.; Alcicek, M.C. The Late Villafranchian large mammals from the Denizli Basin travertines (southwest Anatolia, Turkey). LAnthropologie 2014, 118, 44–73. [Google Scholar] [CrossRef]
- Lebatard, A.-E.; Alçiçek, M.C.; Rochette, P.; Khatib, S.; Vialet, A.; Boulbes, N.; Bourlès, D.L.; Demory, F.; Guipert, G.; Mayda, S. Dating the Homo erectus bearing travertine from Kocabaş (Denizli, Turkey) at at least 1.1 Ma. Earth Planet. Sci. Lett. 2014, 390, 8–18. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gkeme, A.G.; Koufos, G.D.; Kostopoulos, D.S. Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece). Quaternary 2021, 4, 12. https://doi.org/10.3390/quat4020012
Gkeme AG, Koufos GD, Kostopoulos DS. Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece). Quaternary. 2021; 4(2):12. https://doi.org/10.3390/quat4020012
Chicago/Turabian StyleGkeme, Anastasia G., George D. Koufos, and Dimitris S. Kostopoulos. 2021. "Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece)" Quaternary 4, no. 2: 12. https://doi.org/10.3390/quat4020012
APA StyleGkeme, A. G., Koufos, G. D., & Kostopoulos, D. S. (2021). Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece). Quaternary, 4(2), 12. https://doi.org/10.3390/quat4020012