
Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments


Abstract
1. Introduction
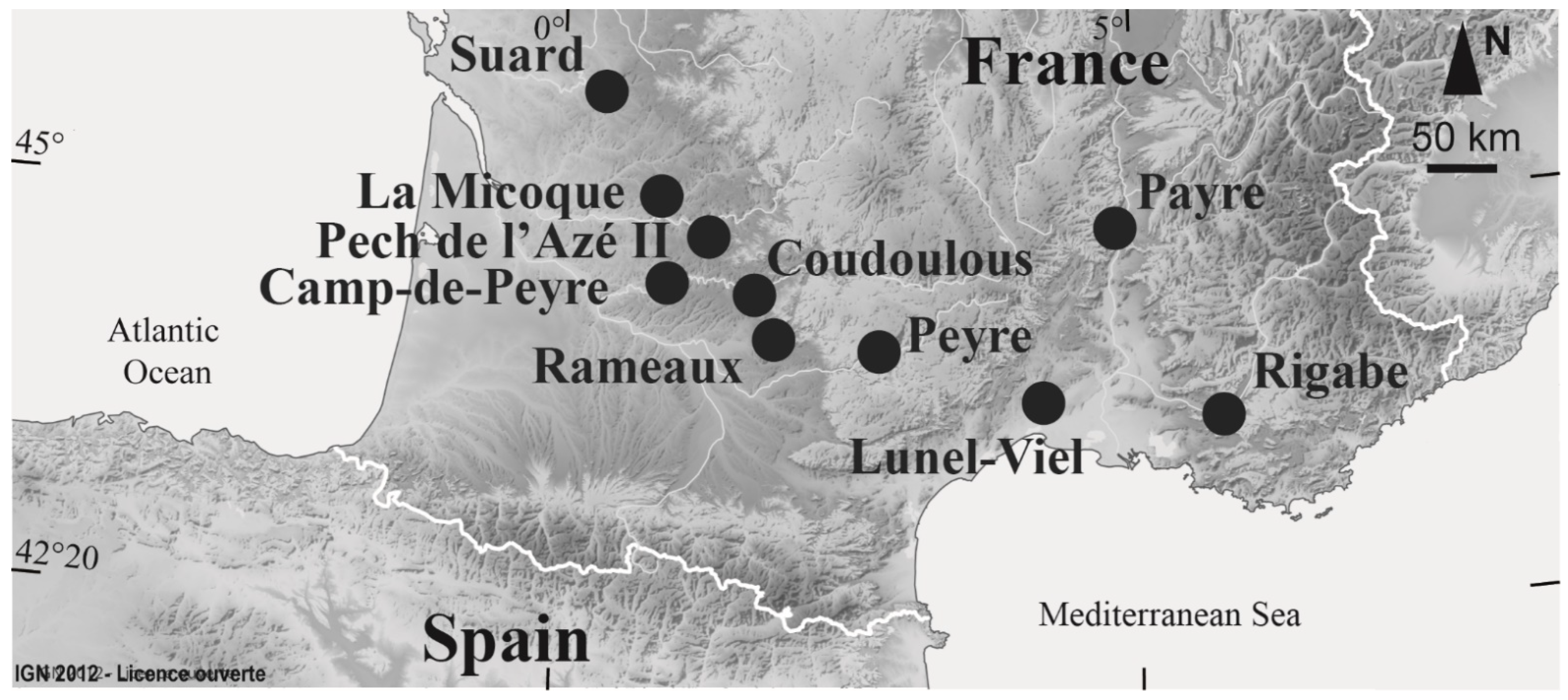
2. Paleoenvironmental Parameters of the Study Sites
3. Biometric and Dental Wear Methods
3.1. Osteologic Measurement Protocols and Taphonomic Limitations
3.2. Body Mass Estimation and Limb Proportions
3.3. Mesowear and Microwear Methods
3.3.1. Dental Mesowear
3.3.2. Dental Microwear
3.4. Statistical Analysis
4. Results
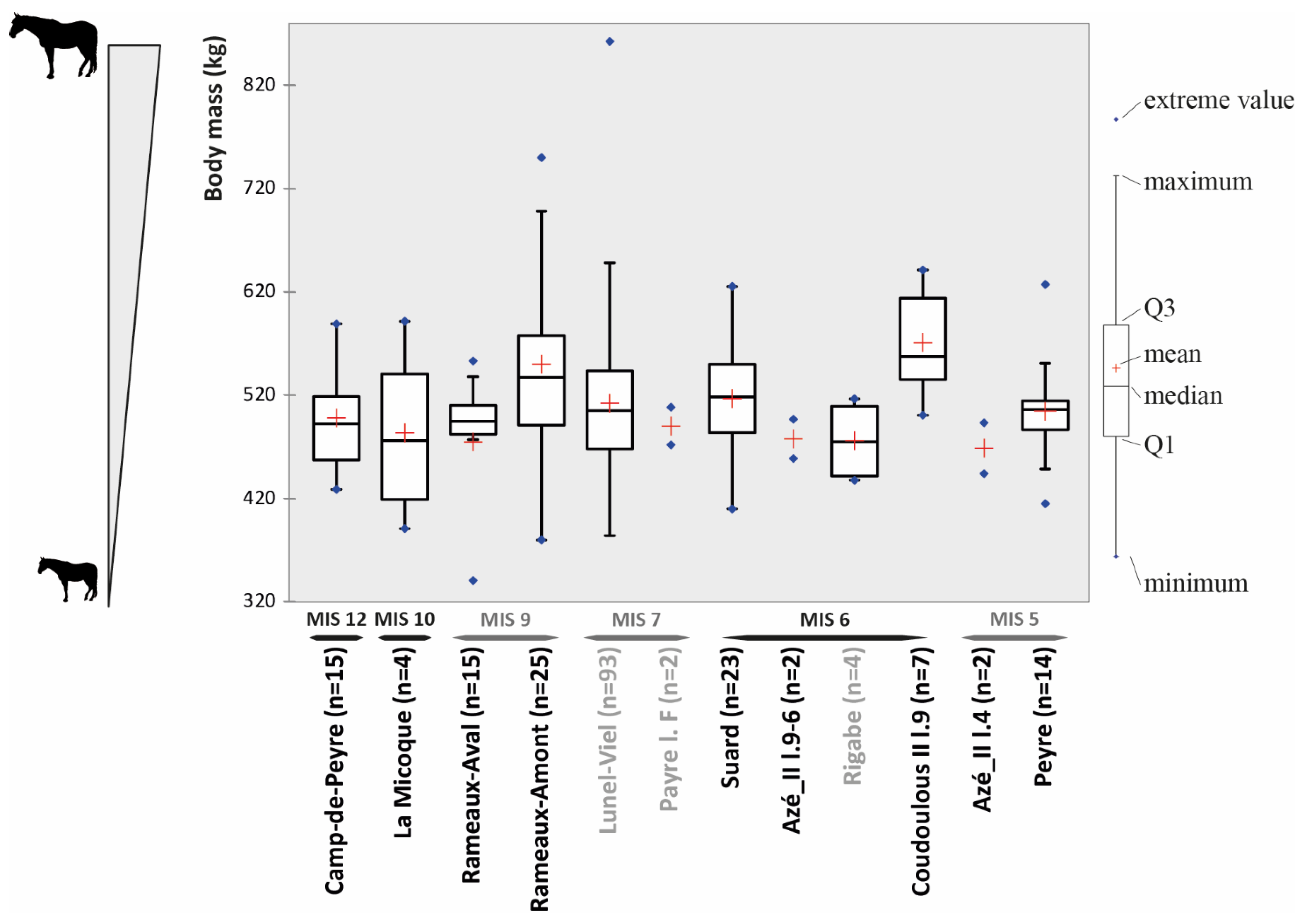
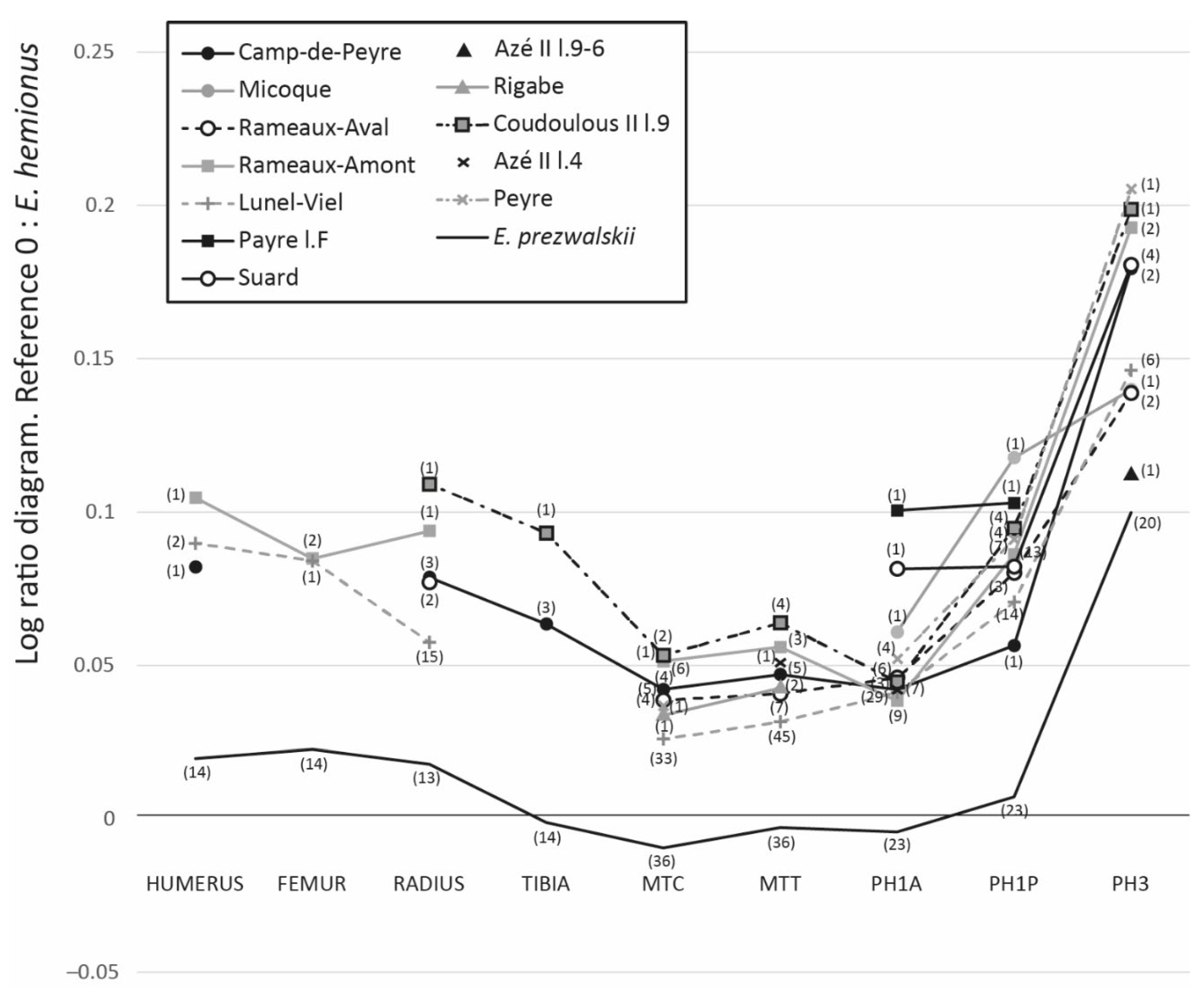
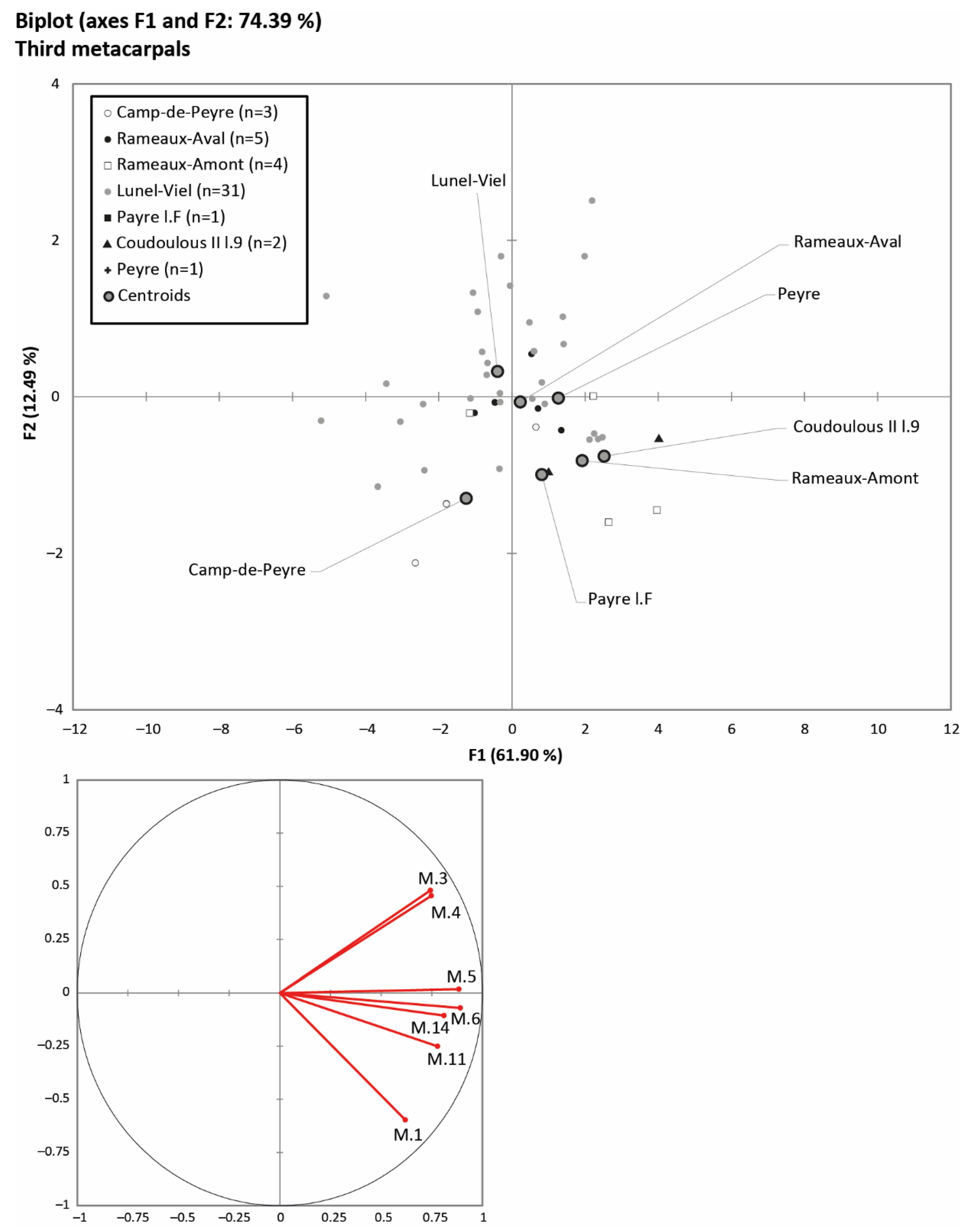
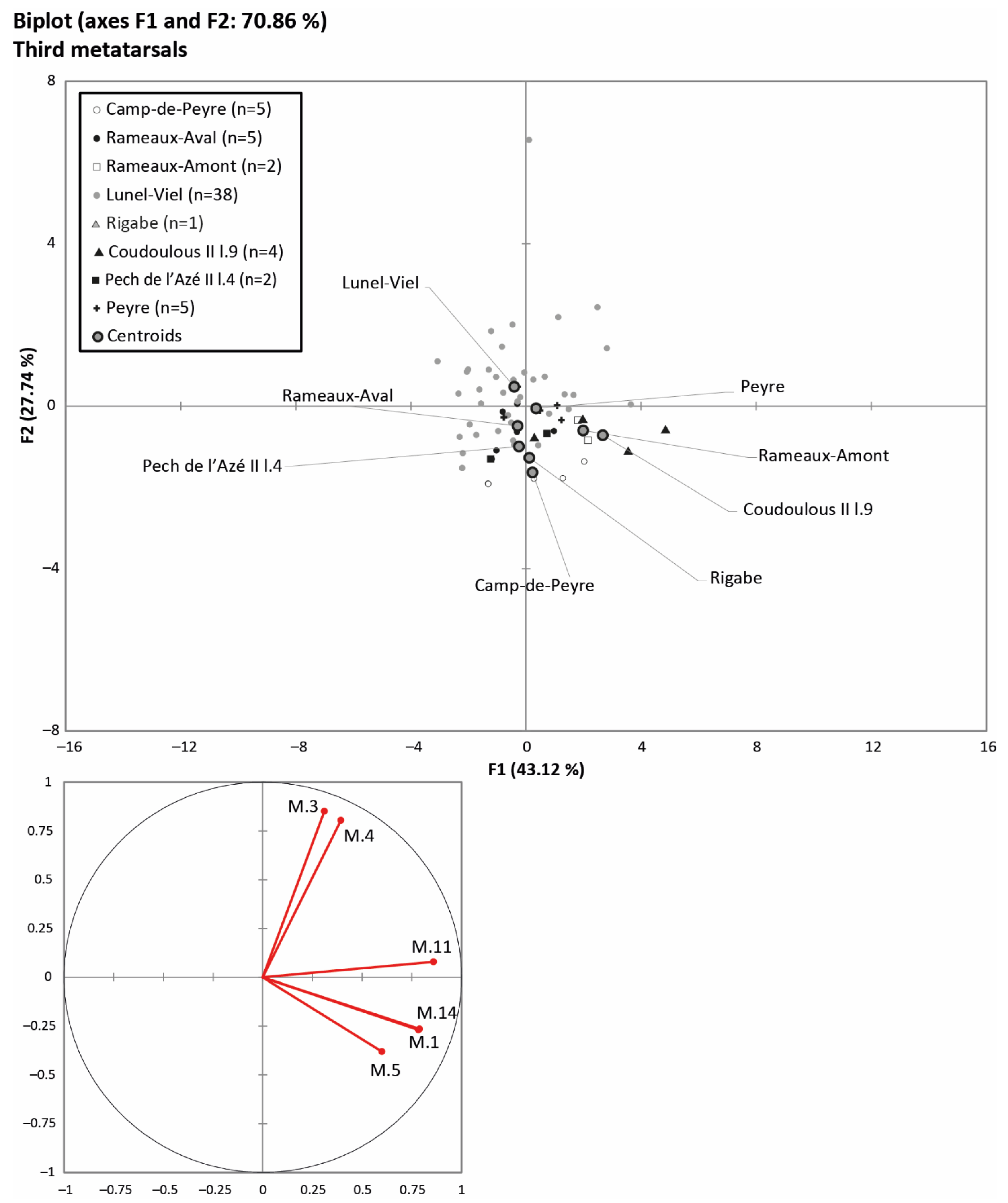
4.1. Horse Size and Ecomorphological Variation through Time
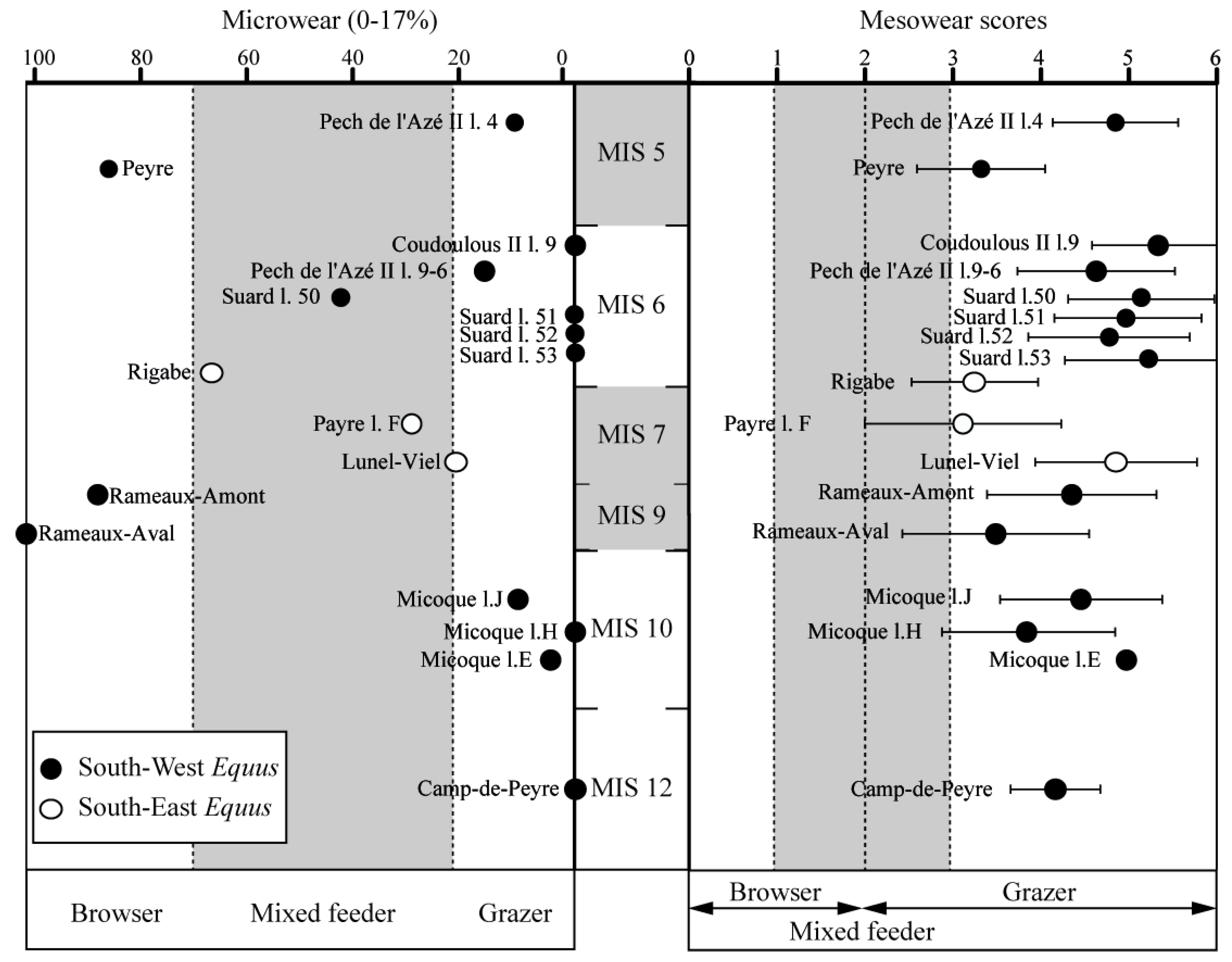
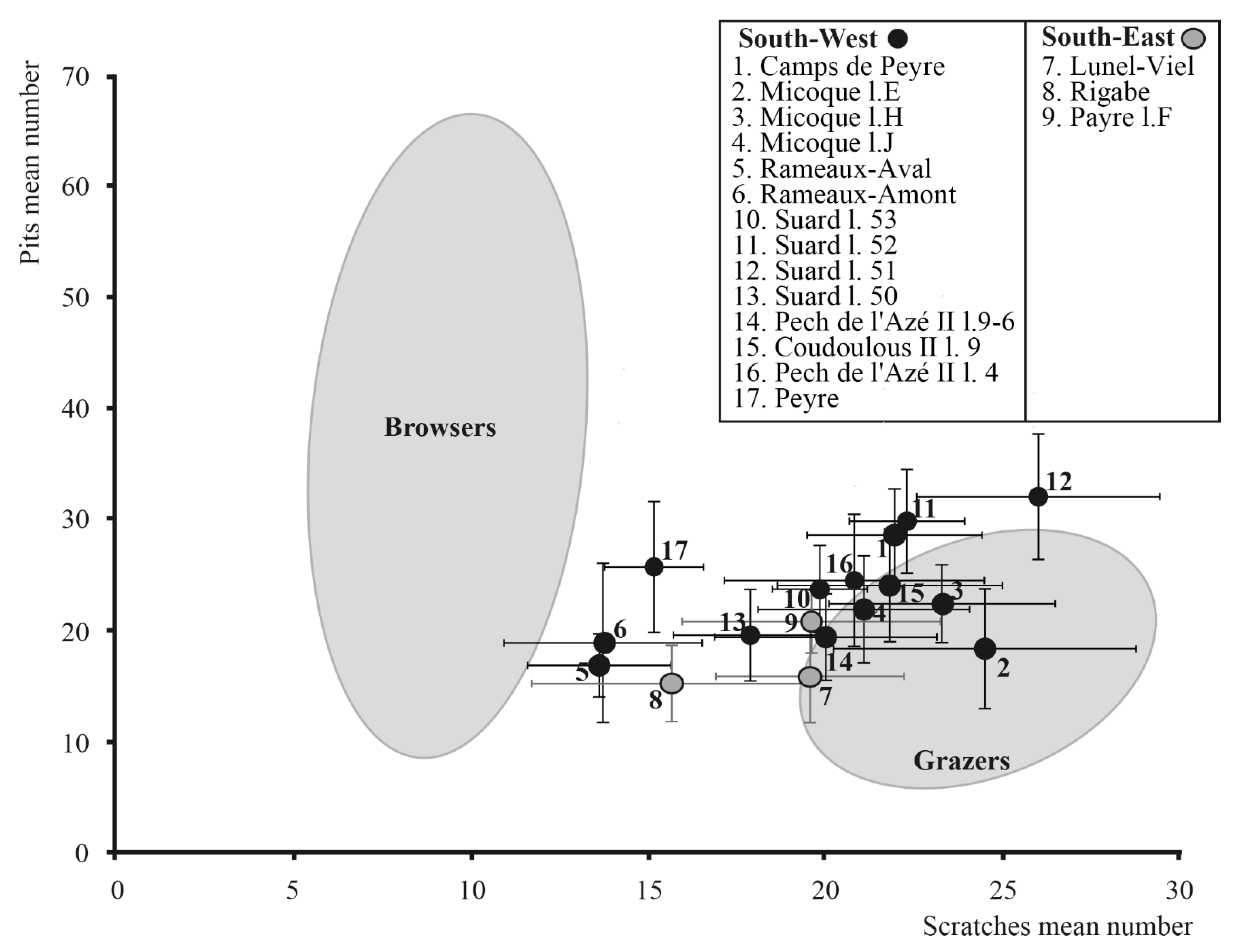
4.2. Dietary Traits of Horses
5. Discussion
5.1. Body Mass and Morphometrical Adaptations to Local Environments
5.2. Horse Dietary Traits at the End of Middle Pleistocene and the Beginning of Late Pleistocene in the South of France
5.3. Relations between Horse Dietary Habits, Body Mass, and Morphometrical Parameters
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ozenda, P. La Végétation Du Continent Européen; Delachaux et Niestlé: Lausanne, Switzerland, 1994; p. 271. [Google Scholar]
- Joannin, S. Changements Climatiques en Méditerranée à la Transition Pléistocène Inférieur-Moyen: Pollens, Isotopes Stables et Cyclostratigraphie. Ph.D. Thesis, Claude Bernard—Lyon I University, Lyon, France, 2007. [Google Scholar]
- Ozenda, P.; Borel, J.-L. An Ecological Map of Europe: Why and How? Compt. Rendus Acad. Sci. Ser. III Sci. Vie 2000, 323, 983–994. [Google Scholar] [CrossRef]
- Petit, R.J.; Aguinagalde, I.; de Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial Refugia: Hotspots But Not Melting Pots of Genetic Diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef]
- Couchoud, I. Etude Pétrographique et Isotopique Des Spéléothèmes Du Sud-Ouest de La France Formés En Contexte Archéologique: Contribution à La Connaissance Des Paléoclimats Régionaux Du Stade Isotopique 5. Ph.D. Thesis, Bordeaux I University, Bordeaux, France, 2006. [Google Scholar]
- Wainer, K.; Genty, D.; Blamart, D.; Hoffmann, D.; Couchoud, I. A New Stage 3 Millennial Climatic Variability Record from a SW France Speleothem. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 271, 130–139. [Google Scholar] [CrossRef]
- Couchoud, I.; Genty, D.; Hoffmann, D.; Drysdale, R.; Blamart, D. Millennial-Scale Climate Variability during the Last Interglacial Recorded in a Speleothem from South-Western France. Quat. Sci. Rev. 2009, 28, 3263–3274. [Google Scholar] [CrossRef]
- Masson-Delmotte, V. Les grandes oscillations du climat depuis 800,000 ans. In Des Climats et des Hommes; Berger, J.-F., Ed.; La Découverte: Paris, France, 2012; pp. 57–72. [Google Scholar]
- Magri, D.; Palombo, M.R. Early to Middle Pleistocene Dynamics of Plant and Mammal Communities in South West Europe. Quat. Int. 2013, 288, 63–72. [Google Scholar] [CrossRef]
- Colleoni, F.; Krinner, G.; Jakobsson, M.; Peyaud, V.; Ritz, C. Influence of Regional Parameters on the Surface Mass Balance of the Eurasian Ice Sheet during the Peak Saalian (140 Kya). Glob. Planet. Chang. 2009, 68, 132–148. [Google Scholar] [CrossRef]
- Colleoni, F.; Krinner, G.; Jakobsson, M. Sensitivity of the Late Saalian (140 Kyrs BP) and LGM (21 Kyrs BP) Eurasian Ice Sheet Surface Mass Balance to Vegetation Feedbacks. Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- van Asperen, E.N. Ecomorphological Adaptations to Climate and Substrate in Late Middle Pleistocene Caballoid Horses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 584–596. [Google Scholar] [CrossRef]
- Rivals, F.; Mihlbachler, M.C.; Solounias, N.; Mol, D.; Semprebon, G.M.; de Vos, J.; Kalthoff, D.C. Palaeoecology of the Mammoth Steppe Fauna from the Late Pleistocene of the North Sea and Alaska: Separating Species Preferences from Geographic Influence in Paleoecological Dental Wear Analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 286, 42–54. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Rivals, F. On the Question of Short-Term Neanderthal Site Occupations: Payre, France (MIS 8-7), and Taubach/Weimar, Germany (MIS 5). J. Anthropol. Res. 2011, 67, 47–75. [Google Scholar] [CrossRef]
- Van Asperen, E.N. Late Middle Pleistocene Horse Fossils from Northwestern Europe as Biostratigraphic Indicators. J. Archaeol. Sci. 2012, 39, 1974–1983. [Google Scholar] [CrossRef]
- Rivals, F.; Julien, M.-A.; Kuitems, M.; Van Kolfschoten, T.; Serangeli, J.; Drucker, D.G.; Bocherens, H.; Conard, N.J. Investigation of Equid Paleodiet from Schöningen 13 II-4 through Dental Wear and Isotopic Analyses: Archaeological Implications. J. Hum. Evol. 2015, 89, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Rivals, F.; Lister, A.M. Dietary Flexibility and Niche Partitioning of Large Herbivores through the Pleistocene of Britain. Quat. Sci. Rev. 2016, 146, 116–133. [Google Scholar] [CrossRef]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing Equid Body Mass Estimate Equations on Modern Zebras—With Implications to Understanding the Relationship of Body Size, Diet, and Habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Prat, F. Recherches sur les Equidés Pléistocènes en France. Ph.D. Thesis, Bordeaux Faculty of Sciences, Bordeaux, France, 1968. [Google Scholar]
- Forsten, A. Size Decrease in Late Pleistocene-Holocene Caballoid Horses (Genus Equus), Intra-or Interspecific Evolution? A Discussion of Alternatives. Quat. Int. 1993, 19, 71–75. [Google Scholar] [CrossRef]
- Magniez, P.; Boulbes, N. Environment during the Middle to Late Palaeolithic Transition in Southern France: The Archaeological Sequence of Tournal Cave (Bize-Minervois, France). Quat. Int. 2014, 337, 43–63. [Google Scholar] [CrossRef]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene Stack of 57 Globally Distributed Benthic Δ18O Records. Paleoceanography 2005, 20, PA1003. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Argant, A.; Boudadi-Maligne, M.; Crégut-Bonnoure, E.; Croitor, R.; Fernandez, P.; Fourvel, J.-B.; Fosse, P.; Guadelli, J.-L.; Labe, B.; et al. Pleistocene Herbivores and Carnivores from France: An Updated Overview of the Literature, Sites and Taxonomy. Ann. Paléontol. 2020, 106, 102384. [Google Scholar] [CrossRef]
- Vernet, J.-L.; Mercier, N.; Bazile, F.; Brugal, J.-P. Travertins et terrasses de la moyenne vallée du Tarn à Millau (Sud du Massif Central, Aveyron, France). Quat. Rev. Assoc. Fr. Pour Létude Quat. 2008, 19, 3–10. [Google Scholar] [CrossRef]
- Uzunidis-Boutillier, A. Grands Herbivores de La Fin Du Pléistocène Moyen Au Début Du Pléistocène Supérieur Dans Le Sud de La France: Implications Anthropologiques Pour La Lignée Néandertalienne. Ph.D. Thesis, Aix-Marseille University, Aix-en-Provence, France, 2017. [Google Scholar]
- Jacobs, Z.; Jankowski, N.R.; Dibble, H.L.; Goldberg, P.; McPherron, S.J.P.; Sandgathe, D.; Soressi, M. The Age of Three Middle Palaeolithic Sites: Single-Grain Optically Stimulated Luminescence Chronologies for Pech de l’Azé I, II and IV in France. J. Hum. Evol. 2016, 95, 80–103. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Aouadi, N.; Mai, B.T.; Cochard, D.; Costamagno, S.; Ellwood, B.; Girard, M.; Jaubert, J.; Jeannet, M.; Kervazo, B.; et al. La Grotte de Coudoulous II (Tour-de-Faure, Lot); Unpublished Excavation Report, Bilan 1997—2000; Pgm P1 Min. de la Culture, SRA Midi-Pyrénées, 2000; 138p. [Google Scholar]
- Uzunidis, A.; Brugal, J.-P. Les grands herbivores (Bovinés, Équidés, Rhinocérotidés, Proboscidiens) de la fin du Pléistocène Moyen: La couche 9 de Coudoulous II (Lot, Quercy, Sud-Ouest France). PALEO 2018, 29, 223–249. [Google Scholar] [CrossRef]
- Schvoerer, M.; Rouanet, J.-F.; Navailles, H.; Debénath, A. Datation Absolue Par Thermoluminescence Des Restes Humains Antéwürmiens de l’abri Suard à La Chaise-de-Vouthon (Charente). Comptes Rendus Acad. Sci. Paris 1977, 284, 1979–1982. [Google Scholar]
- Schwarcz, H.P.; Debénath, A. Datations Absolues Des Restes Humains de La Chaise-de-Vouthon (Charente) Au Moyen Des Déséquilibres Des Séries. Comptes Rendus Acad. Sci. Paris 1979, 288, 1155–1157. [Google Scholar]
- Blackwell, B.; Schwarcz, H.P.; Debénath, A. Absolute Dating of Hominids and Palaeolithic Artifacts of the Cave of La Chaise-de-Vouthon (Charente), France. J. Archaeol. Sci. 1983, 10, 493–513. [Google Scholar] [CrossRef]
- David, P.; Prat, F. Considérations Sur Les Faunes de La Chaise (Commune de Vouthon, Charente), Abris Suard et Bourgeois-Delaunay. Bull. Assoc. Fr. Pour Létude Quat. 1965, 2, 222–232. [Google Scholar] [CrossRef]
- Bonifay, E. Etude Géologique de La Grotte de Rigabe, Artigues (Var). In Proceedings of the Congrès Préhistorique de Monaco, XVIe Session, Monaco, 28 August–5 September 1959; Imprimerie Monnoyer: Le Mans, France, 1959; pp. 265–291. [Google Scholar]
- Moncel, M.-H. Les différentes phases d’occupations humaines à Payre et hypothèses sur les types d’occupation. In Le Site de Payre: Occupations Humaines dans la Vallée du Rhône à la fin du Pléistocène Moyen et au début du Pléistocène Supérieur; Patou-Mathis, M., Moncel, M.-H., Eds.; Société Préhistorique Française: Paris, France, 2008; pp. 309–314. [Google Scholar]
- Brugal, J.-P.; Giuliani, C.; Fosse, P.; Fourvel, J.-B.; Magniez, P.; Pelletier, M.; Uzunidis, A. Preliminary Data on the Middle Pleistocene Site of Lunel Viel I (Hérault, France). Alp. Mediterr. Quat. 2021, 34, 1–13. [Google Scholar] [CrossRef]
- Bonifay, M.-F. Le Cheval du pléistocène moyen des grottes de Lunel-Viel (Hérault). Equus mosbachensis palustris n. ssp. Gall. Préhistoire 1980, 23, 233–281. [Google Scholar] [CrossRef]
- Rouzaud, F.; Soulier, M.; Brugal, J.-P.; Jaubert, J. L’igue Des Rameaux (Saint-Antonin-Noble-Val, Tarn-et-Garonne). Un Nouveau Gisement Du Pléistocène Moyen. Premiers Résultats. PALEO 1990, 2, 89–106. [Google Scholar] [CrossRef][Green Version]
- Argant, A.; Brugal, J.-P. The Cave Lion Panthera (Leo) Spelaea and Its Evolution: Panthera Spelaea Intermedia Nov. Subspecies. Acta Zool. Crac. 2017, 60, 58–103. [Google Scholar] [CrossRef]
- Falguères, C.; Bahain, J.-J.; Saleki, H. U-Series and ESR Dating of Teeth from Acheulian and Mousterian Levels at La Micoque (Dordogne, France). J. Archaeol. Sci. 1997, 24, 537–545. [Google Scholar] [CrossRef]
- Langlois, A. Au Sujet du Cheval de La Micoque (Dordogne) et des Comportements Humains de Subsistance au Pléistocène Moyen Dans le Nord-est de l’Aquitaine. Ph.D. Thesis, Bordeaux I University, Bordeaux, France, 2004. [Google Scholar]
- Langlois, A. Le Cheval du gisement Pléistocène moyen de La Micoque (Les Eyzies-de-Tayac, Dordogne): Equus mosbachensis micoquii nov. ssp. PALEO 2005, 17, 73–110. [Google Scholar] [CrossRef]
- Guadelli, J.-L.; Prat, F. Le Cheval Du Gisement Pléistocène Moyen de Camp-de-Peyre (Sauveterre-La-Lémance, Lot-et-Garonne): Equus mosbachensis campdepeyri n. ssp. PALEO 1995, 7, 85–121. [Google Scholar] [CrossRef]
- Marquet, J.-C. Paléoenvironnement et Chronologie Des Sites Du Domaine Atlantique Fraçais d’âge Pléistocene Moyen et Supérieur d’après l’étude Des Rongeurs. Ph.D. Thesis, Dijon University, Dijon, France, 1993. [Google Scholar]
- Le Tensorer, J.-M. Le Paléolithique de l’Agenais; Cahiers du Quaternaire; Editions du Centre National de la Recherche Scientifique: Paris, France, 1981; 526p. [Google Scholar]
- Texier, J.-P.; Bertran, P. Nouvelle Interprétation Paléoenvironnementale et Chronostratigraphique Du Site Paléolithique de La Micoque (Dordogne): Implications Archéologiques. Comptes Rendus Acad. Sci. Paris 1993, 316, 1611–1617. [Google Scholar]
- Texier, J.-P. Histoire Géologique de Sites Préhistoriques Classiques Du Périgord: Une Vision Actualisée La Micoque, La Grotte Vaufrey, Le Pech de l’Azé I et II, La Ferrassie, l’abri Castanet, Le Flageolet, Laugerie Haute; Documents Préhistoriques; Comité des Travaux Historiques et Scientifiques: Paris, France, 2009; p. 193. [Google Scholar]
- Coumont, M.-P. Taphonomie Préhistorique: Mammifères Fossiles en Contexte d’avens Pièges. Apport Pour L’étude des Archeofaunes. Ph.D. Thesis, Aix-Marseille I University, Aix-en-Provence, France, 2006. [Google Scholar]
- Jeannet, M.; Mein, P. Les Muridae (Mammalia, Rodentia) Du Pléistocène Moyen de l’Igue Des Rameaux (Tarn-et-Garonne, France). PALEO 2016, 27, 177–205. [Google Scholar] [CrossRef]
- Fosse, P. L’hyène Des Cavernes Comme Agent d’accumulation Des Ossements à Lunel-Viel 1 (Hérault, France): Observations Préliminaires. In Outillage peu Élaboré en os et bois de Cervidés; CERDAC: Treignes, Belgium, 1994; pp. 91–96. [Google Scholar]
- Fosse, P. Taphonomie Paléolithique: Les Grands Mammifères de Soleilhac (Haute-Loire) et de Lunel-Viel 1 (Hérault). Ph.D. Thesis, Provence University Aix-Marseille I, Marseille, France, 1994. [Google Scholar]
- Fosse, P. La Grotte N° 1 de Lunel-Viel (Hérault, France): Repaire d’hyènes Du Pléistocène Moyen. Etude Taphonomique Du Matériel Osseux. PALEO 1996, 8, 47–79. [Google Scholar] [CrossRef]
- Uzunidis, A. Dental Wear Analyses of Middle Pleistocene Site of Lunel-Viel (Hérault, France): Did Equus and Bos Live in a Wetland? Quat. Int. 2020, 557, 39–46. [Google Scholar] [CrossRef]
- Desclaux, E.; El Hazzazi, N.; Vilette, P.; Dubar, M. Le contexte environnemental des occupations humaines. L’apport de la microfaune, des restes aviaires et de la malacofaune. In Le site de Payre. Occupations Humaines dans la Vallée du Rhône à la fin du Pléistocène Moyen et au Début du Pléistocène Supérieur; Moncel, M.-H., Ed.; Mémoires de la Société Préhistorique Française; Société Préhistorique Française: Paris, France, 2008; pp. 91–105. [Google Scholar]
- Ecker, M.; Bocherens, H.; Julien, M.-A.; Rivals, F.; Raynal, J.-P.; Moncel, M.-H. Middle Pleistocene Ecology and Neanderthal Subsistence: Insights from Stable Isotope Analyses in Payre (Ardèche, Southeastern France). J. Hum. Evol. 2013, 65, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Bocherens, H.; Díaz-Zorita Bonilla, M.; Daujeard, C.; Fernandes, P.; Raynal, J.-P.; Moncel, M.-H. Direct Isotopic Evidence for Subsistence Variability in Middle Pleistocene Neanderthals (Payre, Southeastern France). Quat. Sci. Rev. 2016, 154, 226–236. [Google Scholar] [CrossRef]
- Dessart, C. Etude de La Faune de La Grotte de Rigabe (Var) (Fouilles E. Bonifay); Post-Graduate Work, Aix-Marseille I-Université de Provence: Marseille, France, 2001. [Google Scholar]
- Paquereau, M.-M. Flore Des Deux Derniers Interglaciaires Dans Le Sud-Ouest de La France. L’Anthropologie 1976, 80, 201–228. [Google Scholar]
- Griggo, C. Significations Environnementales Des Communautés Animales Pléistocènes Reconnues à l’Abri Suard (Charente) et à La Grotte Du Bois-Ragot (Vienne). Ph.D. Thesis, Bordeaux 1 University, Bordeaux, France, 1995. [Google Scholar]
- Griggo, C. Etablissement de Courbes Climatiques Quantifiées à Partir Des Communautés Animales Pléistocènes Suivi d’une Application Aux Gisements de l’Abri Suard (Charente) et La Grotte de Bois-Ragot (Vienne). PALEO 1996, 8, 81–97. [Google Scholar] [CrossRef]
- Paquereau, M.-M. Etude Palynologique Du Wurm I Du Pech de l’Azé (Dordogne). Quaternaria 1969, XI, 227–233. [Google Scholar]
- Goldberg, P. Micromorphology of Pech-de-l’Azé II Sediments. J. Archaeol. Sci. 1979, 6, 17–47. [Google Scholar] [CrossRef]
- Lenoble, A. Le Rôle Du Ruissellement Dans La Formation Des Sites Préhistoriques: Approche Expérimentale. Ph.D. Thesis, Bordeaux 1 University, Bordeaux, France, 2003. [Google Scholar]
- Uzunidis, A. Les Grands Herbivores de La Couche 9 (Fin Pléistocène Moyen) de Coudoulous II (Tour-de-Faure, Lot): Paléontologie, Paléoécologie, Taphonomie; Post-Graduate Work, Aix-Marseille University: Aix-en-Provence, France, 2013. [Google Scholar]
- Astre, G. Brèche Pléistocène de Peyre (Aveyron), Avec Equus hydruntinus et Megaceros. Bull. Soc. Hist. Nat. Toulouse 1963, 98, 321–331. [Google Scholar]
- Brugal, J.-P. Recherches Préliminaires Sur Les Faunes Des Grands Causses et Vallées Périphériques. Paléobiol. Cont. 1981, 12, 145–158. [Google Scholar]
- Vernet, J.-L.; Ambert, P.; Andre, J.; Brugal, J.-P.; Casanova, J.; Michaux, J.; Semah, F. Les Travertins Pléistocènes de La Vallée Du Tarn, Dans Le Sud Du Massif Central: Intérêts Paléoécologiques. In Proceedings of the Paléoécologie, Lyon, France, 18–23 July 1983. [Google Scholar]
- Brugal, J.-P.; Jaubert, J. Les Gisements Paléontologiques Pléistocènes à Indice de Fréquentation Humaine: Un Nouveau Type de Comportement Prédateur ? PALEO 1991, 3, 15–41. [Google Scholar] [CrossRef]
- Brugal, J.-P. Un repaire de hyène des cavernes et de porc-épic à Peyre (Comprégnac, Aveyron). In Les Hommes et leurs Environnements Quaternaires dans les Causses de l’Aveyron; Ambert, P., Vernet, J.-L., Eds.; Association des amis du Musée de Millau: Millau, France, 1993; pp. 47–48. [Google Scholar]
- Sondaar, P.Y. The Osteology of the Manus of Fossil and Recent Equidae with Special Reference to Phylogeny and Function; Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afd. Natuurkunde; N.V. Noord-Hollandsche Utgevers Maatschappij: Amsterdam, The Netherlands, 1968. [Google Scholar]
- Gromova, V. Le Genre “Equus”, Premiere Partie: Revue et Description des Formes. In Annales du Centre d’Etudes et de Documentation Paléontologiques, 1955; n° 13; Pietresson de, S.-A., Translator; Travauxde l’Institut Paléontologique, Académie des Sciences de l’URSS: Paris, France, 1949; Volume 1, t. 17. [Google Scholar]
- Eisenmann, V. Sur Quelques Caractères Adaptatifs Du Squelette d’Equus et Leurs Implications Paléoécologiques. Bull. Muséum Hist. Nat. 1984, 6, 185–195. [Google Scholar]
- Kuzmina, I.E. Horses of North Eurasia from the Pliocene till the present time. In Proceedings of the Zoological Institute; Vereschagin, N.K., Ed.; Russian Academy of Sciences: Saint-Petersbourg, Russia, 1997; Volume 273. [Google Scholar]
- Cramer, B. Morphometrische Untersuchungen an Quartären Pferden in Mitteleuropa. Ph.D. Thesis, Geowissenschaftliche Fakultät, Eberhard-Karls-Universität, Tübingen, Germany, 2002. [Google Scholar]
- Levine, M.A. The use of crown height measurements and eruption-wear sequences to age horse teeth. In Ageing and Sexing of Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; BAR British Series; BAR Publishing: Oxford, UK, 1982; pp. 223–250. [Google Scholar]
- Van Asperen, E.N. Implications of Age Variation and Sexual Dimorphism in Modern Equids for Middle Pleistocene Equid Taxonomy. Int. J. Osteoarchaeol. 2013, 23, 1–12. [Google Scholar] [CrossRef]
- Eisenmann, V. Vera Eisenmann. Available online: https://vera-eisenmann.com/ (accessed on 20 May 2021).
- Eisenmann, V.; Alberdi, M.T.; De Giuli, C.; Staesche, U. Methodology. In Studying Fossil Horses; Woodburne, M.-O., Sondaar, P.Y., Eds.; E. J. Brill: Leiden, The Netherlands, 1988; Volume 1, pp. 1–71. [Google Scholar]
- Eisenmann, V. Les métapodes d’Equus sensu lato (Mammalia, Perissodactyla). Géobios 1979, 12, 863–886. [Google Scholar] [CrossRef]
- Scott, K.M. Postcranial dimensions of ungulates as predictors of body mass. In Body Size in Mammalian Palaeobiology—Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: New York, NY, USA, 1990; pp. 301–355. [Google Scholar]
- Simpson, G.G. Large Pleistocene Felines of North America; American Museum Novitates; American Museum of Natural History: New York, NY, USA, 1941. [Google Scholar]
- Eisenmann, V. Proportions Squelettiques de Chevaux Quaternaires et Actuels. Géobios 1991, 13, 25–32. [Google Scholar] [CrossRef]
- Ackermans, N.L.; Martin, L.F.; Codron, D.; Hummel, J.; Kircher, P.R.; Richter, H.; Kaiser, T.M.; Clauss, M.; Hatt, J.-M. Mesowear Represents a Lifetime Signal in Sheep (Ovis Aries) within a Long-Term Feeding Experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 553, 109793. [Google Scholar] [CrossRef]
- Fortelius, M.; Solounias, N. Functional Characterization of Ungulate Molars Using the Abrasion-Attrition Wear Gradient: A New Method for Reconstructing Paleodiets. Am. Mus. Novit. 2000, 2000, 1–36. [Google Scholar] [CrossRef]
- Grine, F.E. Dental Evidence for Dietary Differences in Australopithecus and Paranthropus: A Quantitative Analysis of Permanent Molar Microwear. J. Hum. Evol. 1986, 15, 783–822. [Google Scholar] [CrossRef]
- Hoffman, J.M.; Fraser, D.; Clementz, M.T. Controlled Feeding Trials with Ungulates: A New Application of in Vivo Dental Molding to Assess the Abrasive Factors of Microwear. J. Exp. Biol. 2015, 218, 1538–1547. [Google Scholar] [CrossRef]
- Merceron, G.; Ramdarshan, A.; Blondel, C.; Boisserie, J.-R.; Brunetiere, N.; Francisco, A.; Gautier, D.; Milhet, X.; Novello, A.; Pret, D. Untangling the Environmental from the Dietary: Dust Does Not Matter. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef]
- Teaford, M.F. A Review of Dental Microwear and Diet in Modern Mammals. Scanning Microsc. 1988, 2, 1149–1166. [Google Scholar] [PubMed]
- Winkler, D.E.; Schulz-Kornas, E.; Kaiser, T.M.; Codron, D.; Leichliter, J.; Hummel, J.; Martin, L.F.; Clauss, M.; Tütken, T. The Turnover of Dental Microwear Texture: Testing the “Last Supper” Effect in Small Mammals in a Controlled Feeding Experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 557, 109930. [Google Scholar] [CrossRef]
- Rivals, F.; Moncel, M.-H.; Patou-Mathis, M. Seasonality and Intra-Site Variation of Neanderthal Occupations in the Middle Palaeolithic Locality of Payre (Ardèche, France) Using Dental Wear Analyses. J. Archaeol. Sci. 2009, 36, 1070–1078. [Google Scholar] [CrossRef]
- Rivals, F.; Mihlbachler, M.C.; Solounias, N. Effect of Ontogenetic-Age Distribution in Fossil and Modern Samples on the Interpretation of Ungulate Paleodiets Using the Mesowear Method. J. Vertebr. Paleontol. 2007, 27, 763–767. [Google Scholar] [CrossRef]
- Saarinen, J.; Lister, A.M. Dental Mesowear Reflects Local Vegetation and Niche Separation in Pleistocene Proboscideans from Britain. J. Quat. Sci. 2016, 31, 799–808. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Solounias, N. Extending the Tooth Mesowear Method to Extinct and Extant Equids. Geodiversitas 2003, 25, 321–345. [Google Scholar]
- Mihlbachler, M.C.; Rivals, F.; Solounias, N.; Semprebon, G.M. Dietary Change and Evolution of Horses in North America. Science 2011, 331, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Rivals, F.; Rindel, D.; Belardi, J.B. Dietary Ecology of Extant Guanaco (Lama Guanicoe) from Southern Patagonia: Seasonal Leaf Browsing and Its Archaeological Implications. J. Archaeol. Sci. 2013, 40, 2971–2980. [Google Scholar] [CrossRef]
- Uzunidis, A.; Rivals, F.; Brugal, J.-P. Relation between Morphology and Dietary Traits in Horse Jugal Upper Teeth during the Middle Pleistocene in Southern France. Quat. Rev. Assoc. Fr. Pour Létude Quat. 2017, 28, 303–312. [Google Scholar] [CrossRef]
- Rivals, F.; Uzunidis, A.; Sanz, M.; Daura, J. Faunal Dietary Response to the Heinrich Event 4 in Southwestern Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 473, 123–130. [Google Scholar] [CrossRef]
- Solounias, N.; Semprebon, G. Advances in the Reconstruction of Ungulate Ecomorphology with Application to Early Fossil Equids. Am. Mus. Novit. 2002, 3366, 49. [Google Scholar] [CrossRef]
- Semprebon, G.; Godfrey, L.R.; Solounias, N.; Sutherland, M.R.; Jungers, W.L. Can Low-Magnification Stereomicroscopy Reveal Diet? J. Hum. Evol. 2004, 115–144. [Google Scholar] [CrossRef]
- Xafis, A.; Nagel, D.; Bastl, K. Which Tooth to Sample? A Methodological Study of the Utility of Premolar/Non-Carnassial Teeth in the Microwear Analysis of Mammals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 229–240. [Google Scholar] [CrossRef]
- Uzunidis, A.; Pineda, A.; Jiménez-Manchón, S.; Xafis, A.; Ollivier, V.; Rivals, F. The Impact of Sediment Abrasion on Tooth Microwear Analysis: An Experimental Study. Archaeol. Anthropol. Sci. 2021, 13, 134. [Google Scholar] [CrossRef]
- Walker, A.; Hoeck, H.N.; Perez, L. Mecrowear of Mammalian Teeth as an Indicator of Diet. Science 1978, 201, 908–910. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F. Was Grass More Prevalent in the Pronghorn Past? An Assessment of the Dietary Adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 253, 332–347. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Sise, P.J.; Coombs, M.C. Potential Bark and Fruit Browsing as Revealed by Stereomicrowear Analysis of the Peculiar Clawed Herbivores Known as Chalicotheres (Perissodactyla, Chalicotherioidea). J. Mamm. Evol. 2011, 18, 33–55. [Google Scholar] [CrossRef]
- Kaczensky, P.; Ganbaatar, O.; Wehrden, H.V.; Walzer, C. Resource Selection by Sympatric Wild Equids in the Mongolian Gobi. J. Appl. Ecol. 2008, 45, 1762–1769. [Google Scholar] [CrossRef]
- Uzunidis, A. Large Ungulates Mobility and Neanderthal Subsistence Behaviours: A Preliminary Tooth Microwear Analysis. J. Archaeol. Sci. Rep. 2020, 29, 102084. [Google Scholar] [CrossRef]
- Foronova, I. Late Quaternary Equids (Genus Equus) of South-Western and Southcentral Siberia. In Proceedings of the Equids in Time and Space: Papers in Honour of Véra Eisenmann: Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, UK, 23–28 August 2002; Mashkour, M., Ed.; Août 2002; Oxbow Books: Oxford, UK, 2006; pp. 20–30. [Google Scholar]
- Forsten, A. Middle Pleistocene Replacement of Stenonoid Horses by Caballoid Horses—Ecological Implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. Elsevier Sci. Publ. BV 1988, 65, 23–33. [Google Scholar] [CrossRef]
- Bro-Jørgensen, J. Dense Habitats Selecting for Small Body Size: A Comparative Study on Bovids. Oikos 2008, 117, 729–737. [Google Scholar] [CrossRef]
- Lindstedt, S.L.; Boyce, M.S. Seasonality, Fasting Endurance, and Body Size in Mammals. Am. Nat. 1985, 125, 873–878. [Google Scholar] [CrossRef]
- El Hazzazi, N. Apport Du Genre Arvicola (Rodentia, Mammalia) Dans La Connaissance de La Chronostratigraphie Des Sites Pléistocène Moyen et Supérieur: Orgnac 3, Payre 2 et l’abri Des Pêcheurs (Ardèche, France). In Proceedings of the Theoretical and Methodological Problems—Archaeometry—Paleoecology, Forli, Italy, 8–14 September 1996; Arias, C., Bietti, A., Castelletti, L., Peretto, C., Eds.; A.B.A.C.O. Edizioni: Forli, Italy, 1998; Volume 1, pp. 125–133. [Google Scholar]
- Moncel, M.-H.; Debard, E.; Desclaux, E.; Dubois, J.-M.; Lamarque, F.; Patou-Mathis, M.; Villette, P. Le Cadre de Vie Des Hommes Du Paléolithique Moyen (Stades Isotopiques 6 et 5) Dans Le Site de Payre (Rompon, Ardèche): D’une Grotte à Un Abri Sous Roche Effondré. Bull. Soc. Préhistorique Fr. 2002, 99, 249–273. [Google Scholar] [CrossRef]
- Schulz, E.; Kaiser, T.M. Historical Distribution, Habitat Requirements and Feeding Ecology of the Genus Equus (Perissodactyla). Mammal Rev. 2013, 43, 111–123. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Franz-Odendaal, T.A. A Mixed-Feeding Equus Species from the Middle Pleistocene of South Africa. Quat. Res. 2004, 62, 316–323. [Google Scholar] [CrossRef]
- Valli, A.M.F.; Palombo, M.R.; Alberdi, M.T. How Homogeneous Are Microwear Patterns on a Fossil Horse Tooth? Preliminary Test on a Premolar of Equus Altidens from Barranco Leon 5 (Spain). Alp. Mediterr. Quat. 2012, 25, 25–33. [Google Scholar]
- Kelly, A.; Miller, J.H.; Wooller, M.J.; Seaton, C.T.; Druckenmiller, P.; DeSantis, L. Dietary Paleoecology of Bison and Horses on the Mammoth Steppe of Eastern Beringia Based on Dental Microwear and Mesowear Analyses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 572, 110394. [Google Scholar] [CrossRef]
- Rivals, F.; Kitagawa, K.; Julien, M.-A.; Patou-Mathis, M.; Bessudnov, A.A.; Bessudnov, A.N. Straight from the Horse’s Mouth: High-Resolution Proxies for the Study of Horse Diet and Its Relation to the Seasonal Occupation Patterns at Divnogor’ye 9 (Middle Don, Central Russia). Quat. Int. 2018, 474, 146–155. [Google Scholar] [CrossRef]
- Delpech, F.; Le Tensorer, J.-M.; Pineda, R.; Prat, F. Un Nouveau Gisement Du Pleistocène Moyen: Camp-de-Peyre à Sauveterre-La-Lémance (Lot-et-Garonne). Comptes Rendus Acad. Sci. Paris D 1978, 286, 1101–1103. [Google Scholar]
- Saarinen, J.; Eronen, J.; Fortelius, M.; Seppä, H.; Lister, A.M. Patterns of Diet and Body Mass of Large Ungulates from the Pleistocene of Western Europe, and Their Relation to Vegetation. Palaeontol. Electron. 2016, 19.3.32A, 1–58. [Google Scholar] [CrossRef]
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; ISBN 978-0-8117-0496-0. [Google Scholar]
- Guthrie, R.D. Origin and Causes of the Mammoth Steppe: A Story of Cloud Cover, Woolly Mammal Tooth Pits, Buckles, and inside-out Beringia. Quat. Sci. Rev. 2001, 20, 549–574. [Google Scholar] [CrossRef]
- Saarinen, J. Ecometrics of Large Herbivorous Land Mammals in Relation to Climatic and Environmental Changes during the Pleistocene. Ph.D. Thesis, Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland, 2014. [Google Scholar]
- Blackburn, T.M.; Gaston, K.J.; Loder, N. Geographic Gradients in Body Size: A Clarification of Bergmann’s Rule. Divers. Distrib. 1999, 5, 165–174. [Google Scholar] [CrossRef]
- Boulbes, N.; van Asperen, E.N. Biostratigraphy and Palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef]
- Richerson, P.J.; Boyd, R.; Bettinger, R.L. Was Agriculture Impossible during the Pleistocene but Mandatory during the Holocene? A Climate Change Hypothesis. Am. Antiq. 2001, 66, 387–411. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Region | Age | Reference for Dating | Location | Equus Specie | Reference for Palaeontological Determination | Number of Studied Bones | Number of Studied Teeth |
|---|---|---|---|---|---|---|---|---|
| Peyre | South-West | MIS 5 | [24] | MMSH | Equus caballus | [25] | 138 | 27 |
| Pech de l’Azé II l.4-2 | South-West | MIS 5 | [26] | Les Eyzies MNP | Equus caballus | [19,25] | 18 | 31 |
| Coudoulous II l.9 | South-West | MIS 6 | [27,28] | MMSH | Equus mosbachensis | [25,28] | 115 | 25 |
| Pech de l’Azé II l. 9-6 | South-West | MIS 6 | [26] | Les Eyzies MNP | Equus mosbachensis | [19,25] | 6 | 63 |
| Suard l.53-50 | South-West | MIS 6 | [29,30,31] | Angoulême museum | Equus caballus piveteaui | [25,32] | 190 | 116 |
| Rigabe | South-East | MIS 6 | [33] | Les Eyzies MNP | Equus mosbachensis palustris | [25] | 29 | 18 |
| Payre l. F | South-East | MIS 7 | [34] | Orgnac museum | Equus mosbachensis | [25] | 26 | 8 |
| Lunel-Viel | South-East | MIS 7 | [35] | Les Eyzies MNP | Equus mosbachensis palustris | [25,36] | 1109 | 65 |
| Rameaux-Amont | South-West | MIS 9 | [25,37,38] | MMSH | Equus mosbachensis | [25] | 247 | 151 |
| Rameaux-Aval | South West | MIS 9 | [25,37,38] | MMSH | Equus mosbachensis | [25] | 190 | 8 |
| La Micoque l. E, H, J | South-West | MIS 10 | [39] | Les Eyzies MNP | Equus mosbachensis | [25,40,41] | 38 | 34 |
| Camp-de-Peyre | South-West | MIS 12 | [42] | Les Eyzies MNP | Equus mosbachensis campdepeyri | [25,42] | 164 | 38 |
| Site/Bone | Humerus | Radius | Femur | Tibia | Metacarpal | Metatarsal | Phalange I A | Phalange I P | Phalange III | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | m | n | m | n | m | n | m | n | m | n | m | n | m | n | m | n | m | |
| Camp-de-Peyre | 1 | 300.15 | 3 | 355.43 | 3 | 373.27 | 4 | 242.69 | 5 | 287.78 | 7 | 86.9 | 1 | 82.65 | 2 | 85.03 | ||
| La Micoque | 1 | 90.73 | 1 | 95.18 | 1 | 77.65 | ||||||||||||
| Rameaux-Aval | 2 | 354.08 | 5 | 240.75 | 7 | 283.65 | 6 | 87.68 | 3 | 87.28 | 2 | 77.45 | ||||||
| Rameaux-Amont | 1 | 316.14 | 1 | 368 | 2 | 410.44 | 6 | 247.89 | 3 | 293.79 | 9 | 86.16 | 7 | 88.51 | 2 | 87.7 | ||
| Lunel-Viel | 2 | 305.51 | 15 | 338.48 | 1 | 409.74 | 33 | 233.81 | 45 | 277.75 | 29 | 86.7 | 14 | 85.4 | 6 | 78.78 | ||
| Payre l.F | 1 | 249 | 1 | 99.4 | 1 | 92 | ||||||||||||
| Suard | 3 | 95.14 | 13 | 87.7 | 4 | 85.27 | ||||||||||||
| Pech de l’Azé II l.9-6 | 1 | 72.81 | ||||||||||||||||
| Rigabe | 1 | 237.99 | 2 | 284.88 | ||||||||||||||
| Coudoulous II l.9 | 1 | 381.19 | 1 | 399.68 | 2 | 249 | 4 | 299.175 | 3 | 87.4 | 4 | 90.25 | 1 | 88.9 | ||||
| Pech de l’Azé II l.4 | 2 | 290.43 | ||||||||||||||||
| Peyre | 4 | 239.71 | 5 | 285.09 | 4 | 88.9 | 4 | 89.52 | 1 | 90.26 | ||||||||
| Site | 1 | 3 | 4 | 5 | 6 | 10 | 11 | 12 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Camp-de-Peyre | n | 4 | 7 | 7 | 4 | 5 | 4 | 4 | 4 | 4 |
| m | 242.70 | 37.92 | 29.42 | 54.29 | 36.95 | 53.33 | 52.97 | 38.82 | 32.99 | |
| s | 4.43 | 1.98 | 0.99 | 2.16 | 1.40 | 3.07 | 2.00 | 0.42 | 1.97 | |
| min | 238.25 | 35.98 | 28.18 | 51.34 | 34.55 | 51.05 | 50.60 | 38.21 | 30.47 | |
| max | 248.16 | 40.28 | 30.83 | 56.30 | 37.99 | 57.86 | 55.26 | 39.18 | 34.84 | |
| Micoque | n | 1 | 1 | 3 | 5 | 1 | 1 | 1 | 1 | |
| m | 43.65 | 26.49 | 55.19 | 35.88 | 52.03 | 53.03 | 40.60 | 34.18 | ||
| s | 3.04 | 1.21 | ||||||||
| min | 52.61 | 34.50 | ||||||||
| max | 58.54 | 37.67 | ||||||||
| Rameaux-Aval | n | 5 | 8 | 8 | 7 | 7 | 8 | 6 | 6 | 6 |
| m | 240.76 | 38.36 | 28.78 | 55.26 | 36.15 | 51.74 | 52.83 | 39.55 | 32.90 | |
| s | 4.08 | 4.69 | 2.69 | 1.97 | 2.11 | 2.66 | 0.97 | 1.18 | 0.96 | |
| min | 237.20 | 29.85 | 24.18 | 52.31 | 33.31 | 47.26 | 51.70 | 37.69 | 31.71 | |
| max | 247.40 | 42.71 | 31.18 | 57.66 | 38.86 | 54.10 | 54.19 | 40.72 | 33.80 | |
| Rameaux-Amont | n | 6 | 9 | 9 | 8 | 7 | 7 | 6 | 5 | 7 |
| m | 247.89 | 40.23 | 30.54 | 55.86 | 37.60 | 54.62 | 54.42 | 40.59 | 33.05 | |
| s | 12.18 | 2.03 | 0.88 | 2.43 | 2.05 | 3.06 | 2.55 | 1.98 | 1.88 | |
| min | 235.99 | 37.60 | 29.25 | 52.13 | 34.74 | 51.44 | 51.10 | 38.17 | 29.59 | |
| max | 267.10 | 42.77 | 31.75 | 60.13 | 40.83 | 59.53 | 56.97 | 42.57 | 35.01 | |
| Lunel-Viel | n | 33 | 43 | 44 | 37 | 38 | 37 | 37 | 34 | 37 |
| m | 233.81 | 39.54 | 30.26 | 54.13 | 36.35 | 51.77 | 52.07 | 40.47 | 32.90 | |
| s | 5.92 | 3.52 | 2.41 | 3.26 | 2.56 | 2.33 | 2.46 | 1.74 | 1.89 | |
| min | 221.20 | 26.16 | 24.26 | 46.82 | 31.34 | 47.17 | 45.51 | 37.53 | 27.79 | |
| max | 242.90 | 45.48 | 40.69 | 58.80 | 39.73 | 58.15 | 57.65 | 46.46 | 39.81 | |
| Payre l.F | n | 1 | 1 | 1 | 1 | 4 | 2 | 2 | 2 | 2 |
| m | 249.00 | 41.50 | 29.30 | 54.40 | 37.43 | 52.45 | 53.30 | 41.25 | 34.35 | |
| s | 2.06 | 0.35 | 0.14 | 1.20 | 0.07 | |||||
| min | 35.00 | 52.20 | 53.20 | 40.40 | 34.30 | |||||
| max | 40.00 | 52.70 | 53.40 | 42.10 | 34.40 | |||||
| Suard | n | 5 | 6 | 13 | 15 | 2 | 2 | 2 | 2 | |
| m | 42.87 | 31.02 | 56.30 | 36.81 | 56.28 | 55.42 | 41.23 | 34.26 | ||
| s | 1.40 | 0.61 | 2.01 | 1.15 | 0.34 | 0.21 | 0.57 | 0.30 | ||
| min | 41.39 | 30.20 | 53.67 | 33.88 | 56.04 | 55.27 | 40.83 | 34.05 | ||
| max | 45.15 | 32.02 | 60.05 | 38.61 | 56.52 | 55.56 | 41.63 | 34.47 | ||
| Pech de l’Azé II | n | 1 | 1 | 2 | ||||||
| l. 9-6 | m | 28.74 | 57.49 | 37.88 | ||||||
| s | 0.71 | |||||||||
| min | 37.38 | |||||||||
| max | 38.38 | |||||||||
| Rigabe | n | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| m | 237.99 | 37.33 | 28.67 | 34.16 | 49.36 | 49.48 | 38.65 | 31.36 | ||
| Coudoulous II l.9 | n | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| m | 249.00 | 41.32 | 31.10 | 58.30 | 38.70 | 54.90 | 55.60 | 39.20 | 34.26 | |
| s | 1.41 | 2.91 | 0.42 | 2.83 | 2.12 | 1.56 | 2.83 | 1.70 | 0.21 | |
| min | 248.00 | 39.26 | 30.80 | 56.30 | 37.20 | 53.80 | 53.60 | 38.00 | 34.11 | |
| max | 250.00 | 43.37 | 31.40 | 60.30 | 40.20 | 56.00 | 57.60 | 40.40 | 34.40 | |
| Peyre | n | 4 | 4 | 4 | 2 | 2 | 4 | 4 | 3 | 4 |
| m | 239.72 | 39.75 | 30.45 | 57.54 | 37.12 | 50.24 | 50.90 | 38.92 | 32.40 | |
| s | 7.05 | 0.46 | 1.42 | 0.66 | 0.09 | 2.03 | 1.73 | 0.67 | 1.21 | |
| min | 233.59 | 39.22 | 29.08 | 57.07 | 37.05 | 47.22 | 48.36 | 38.19 | 30.64 | |
| max | 249.32 | 40.22 | 32.34 | 58.00 | 37.18 | 51.53 | 52.08 | 39.49 | 33.37 |
| Site | 1 | 3 | 4 | 5 | 6 | 10 | 11 | 12 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Camp-de-Peyre | n | 5 | 8 | 8 | 6 | 5 | 7 | 7 | 7 | 7 |
| m | 287.78 | 37.91 | 36.07 | 56.91 | 48.62 | 54.81 | 54.19 | 42.17 | 35.76 | |
| s | 7.59 | 1.52 | 1.17 | 2.96 | 2.27 | 2.14 | 1.98 | 1.69 | 1.29 | |
| min | 282.58 | 35.25 | 34.53 | 51.79 | 46.00 | 52.42 | 51.57 | 39.37 | 33.95 | |
| max | 301.08 | 40.35 | 37.85 | 59.46 | 51.09 | 58.65 | 57.01 | 43.95 | 37.15 | |
| Micoque | n | 2 | ||||||||
| m | 38.52 | |||||||||
| s | 3.71 | |||||||||
| min | 35.89 | |||||||||
| max | 41.14 | |||||||||
| Rameaux-Aval | n | 7 | 7 | 7 | 5 | 5 | 7 | 7 | 6 | 7 |
| m | 283.65 | 37.17 | 36.12 | 55.96 | 49.60 | 54.31 | 52.69 | 39.76 | 33.48 | |
| s | 5.96 | 3.04 | 2.74 | 1.08 | 2.49 | 1.64 | 2.00 | 1.18 | 1.07 | |
| min | 274.11 | 30.55 | 30.98 | 54.72 | 47.03 | 51.57 | 48.57 | 37.84 | 31.40 | |
| max | 290.80 | 39.80 | 38.56 | 57.36 | 53.71 | 55.82 | 54.99 | 41.32 | 34.90 | |
| Rameaux-Amont | n | 3 | 12 | 13 | 5 | 4 | 6 | 6 | 6 | 6 |
| m | 293.79 | 40.31 | 35.35 | 56.00 | 49.40 | 54.94 | 54.28 | 41.83 | 34.71 | |
| s | 13.38 | 1.83 | 4.71 | 2.14 | 2.70 | 2.88 | 1.23 | 2.07 | 1.29 | |
| min | 278.79 | 37.41 | 28.98 | 53.71 | 45.85 | 50.13 | 52.90 | 38.89 | 32.79 | |
| max | 304.50 | 43.13 | 44.71 | 58.41 | 52.38 | 58.38 | 55.86 | 43.87 | 36.07 | |
| Lunel-Viel | n | 44 | 51 | 50 | 42 | 41 | 47 | 48 | 45 | 48 |
| m | 277.63 | 41.00 | 37.73 | 55.59 | 49.45 | 54.37 | 53.79 | 42.12 | 33.75 | |
| s | 7.63 | 3.07 | 2.46 | 3.65 | 3.24 | 2.66 | 2.45 | 4.76 | 1.42 | |
| min | 262.61 | 36.24 | 33.61 | 40.47 | 38.32 | 50.30 | 48.43 | 38.84 | 30.89 | |
| max | 294.75 | 50.28 | 47.27 | 65.34 | 56.30 | 60.87 | 61.46 | 71.77 | 37.58 | |
| Suard | n | 3 | 2 | 6 | 6 | 2 | 2 | 2 | 2 | |
| m | 40.41 | 35.54 | 58.11 | 48.14 | 55.28 | 57.59 | 44.06 | 35.86 | ||
| s | 1.84 | 4.07 | 1.54 | 2.12 | 0.34 | 0.23 | 1.22 | 0.28 | ||
| min | 38.39 | 32.66 | 55.19 | 45.65 | 55.04 | 57.42 | 43.20 | 35.66 | ||
| max | 42.00 | 38.41 | 59.84 | 51.45 | 55.52 | 57.75 | 44.92 | 36.05 | ||
| Pech de l’Azé II | n | 1 | 1 | |||||||
| l. 9-6 | m | 56.84 | 52.37 | |||||||
| Rigabe | n | 2 | 3 | 3 | 1 | 2 | 3 | 3 | 2 | 2 |
| m | 284.88 | 39.05 | 35.53 | 56.22 | 47.58 | 53.14 | 50.82 | 43.89 | 35.61 | |
| s | 1.00 | 1.61 | 1.94 | 0.90 | 1.98 | 2.15 | 1.12 | 1.07 | ||
| min | 284.17 | 37.23 | 34.39 | 46.94 | 51.31 | 48.38 | 43.10 | 34.85 | ||
| max | 285.59 | 40.28 | 37.77 | 48.21 | 55.24 | 52.44 | 44.68 | 36.36 | ||
| Coudoulous II l.9 | n | 4 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| m | 299.18 | 40.86 | 37.21 | 59.45 | 49.45 | 57.42 | 57.76 | 44.86 | 37.37 | |
| s | 9.06 | 0.63 | 1.33 | 3.90 | 1.03 | 2.56 | 3.11 | 1.84 | 1.83 | |
| min | 292.20 | 40.00 | 35.59 | 56.44 | 48.18 | 54.20 | 53.16 | 42.30 | 34.68 | |
| max | 311.80 | 41.62 | 39.30 | 65.44 | 50.90 | 60.31 | 60.42 | 46.70 | 39.34 | |
| Pech de l’Azé II | n | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| l.4 | m | 290.44 | 38.16 | 36.36 | 53.93 | 46.33 | 55.16 | 52.66 | 41.41 | 34.97 |
| s | 4.87 | 0.32 | 2.32 | 2.86 | 0.62 | 0.64 | 2.07 | 0.22 | 0.06 | |
| min | 286.99 | 37.93 | 34.72 | 51.90 | 45.89 | 54.70 | 51.19 | 41.25 | 34.93 | |
| max | 293.88 | 38.38 | 38.00 | 55.95 | 46.77 | 55.61 | 54.12 | 41.56 | 35.01 | |
| Peyre | n | 5 | 7 | 7 | 7 | 6 | 6 | 6 | 4 | 6 |
| m | 285.10 | 40.34 | 37.40 | 56.27 | 46.78 | 55.65 | 53.10 | 42.16 | 34.94 | |
| s | 9.90 | 0.72 | 1.51 | 1.30 | 1.77 | 2.05 | 0.92 | 1.01 | 1.63 | |
| min | 268.84 | 38.90 | 35.54 | 54.55 | 44.57 | 52.68 | 52.03 | 40.72 | 32.10 | |
| max | 292.30 | 40.93 | 38.91 | 58.75 | 49.12 | 58.74 | 54.59 | 42.93 | 36.48 |
| Site | Mesowear | Microwear | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | MWS | N | NP | NS | LP% | G% | SWS | HC% | 0–17% | ||
| Peyre | M | 27 | 3.33 | 14 | 25.71 | 15.21 | 50 | 28.57 | 0.92 | 7.14 | 85.71 |
| SD | 0.73 | 5.89 | 1.40 | ||||||||
| Pech de l’Azé II l.4-2 | M | 31 | 4.87 | 18 | 24.50 | 20.86 | 0 | 11.11 | 1 | 27.78 | 11.11 |
| SD | 0.72 | 5.94 | 3.67 | ||||||||
| Coudoulous II l.9 | M | 25 | 5.36 | 11 | 24.05 | 21.86 | 63.63 | 0 | 1 | 0 | 0 |
| SD | 0.76 | 5.07 | 3.17 | ||||||||
| Pech de l’Azé II l. 9-6 | M | 63 | 4.65 | 42 | 18.13 | 20.21 | 0 | 23.81 | 1 | 27.78 | 16.67 |
| SD | 0.9 | 4.25 | 2.63 | ||||||||
| Suard l. 50 | M | 12 | 5.17 | 7 | 19.57 | 17.93 | 57.14 | 14.28 | 1 | 0 | 42.86 |
| SD | 0.83 | 4.14 | 2.17 | ||||||||
| Suard l. 51 | M | 70 | 5.01 | 28 | 32.04 | 26.05 | 3.57 | 0 | 1 | 3.57 | 0 |
| SD | 0.84 | 5.66 | 3.42 | ||||||||
| Suard l. 52 | M | 30 | 4.80 | 26 | 29.83 | 22.35 | 3.84 | 3.84 | 0.92 | 0 | 0 |
| SD | 0.92 | 4.66 | 1.62 | ||||||||
| Suard l. 53 | M | 4 | 5.25 | 9 | 23.72 | 19.89 | 0 | 0 | 0.89 | 0 | 0 |
| SD | 0.96 | 3.91 | 1.34 | ||||||||
| Rigabe | M | 18 | 4.94 | 12 | 15.21 | 15.71 | 75 | 50 | 0.5 | 0 | 66.67 |
| SD | 0.73 | 3.44 | 3.95 | ||||||||
| Payre l. F * | M | 8 | 3.13 | 10 | 20.80 | 19.65 | 20 | 10 | 1.1 | 63.63 | 30 |
| SD | 1.13 | 2.84 | 3.64 | ||||||||
| Lunel-Viel | M | 65 | 4.88 | 36 | 15.85 | 19.61 | 31.43 | 8.33 | 1 | 2.78 | 22.22 |
| SD | 0.93 | 4.17 | 2.65 | ||||||||
| Rameaux-Amont | M | 151 | 4.37 | 24 | 18.88 | 13.77 | 37.5 | 12.5 | 1.04 | 4.17 | 87.5 |
| SD | 0.97 | 7.19 | 2.79 | ||||||||
| Rameaux-Aval | M | 8 | 3.50 | 3 | 16.83 | 13.67 | 66.66 | 33.33 | 1 | 0 | 100 |
| SD | 1.07 | 2.84 | 2.02 | ||||||||
| La Micoque l. J | M | 21 | 4.48 | 19 | 21.89 | 21.13 | 94.74 | 42.10 | 1.05 | 10.53 | 10.53 |
| SD | 0.93 | 4.82 | 2.99 | ||||||||
| La Micoque l. H | M | 8 | 3.88 | 15 | 22.40 | 23.33 | 46.66 | 0 | 1 | 0 | 0 |
| SD | 0.99 | 3.52 | 3.19 | ||||||||
| La Micoque l. E | M | 5 | 5 | 22 | 18.34 | 24.55 | 0 | 0 | 0.72 | 0 | 4.55 |
| SD | 0 | 5.41 | 4.27 | ||||||||
| Camp-de-Peyre | M | 38 | 4.18 | 7 | 28.57 | 22.00 | 0 | 0 | 0.71 | 14.28 | 0 |
| SD | 0.51 | 4.17 | 2.47 | ||||||||
| Group | n | m | s | p-Values |
|---|---|---|---|---|
| Western cohorts | 9 | 504.91 | 35.67 | 0.8636 |
| Eastern cohorts | 3 | 492.88 | 17.89 | |
| Cold IS | 6 | 502.22 | 37.91 | 0.9372 |
| Temperate IS | 6 | 501.59 | 28.13 |
| Group | n | m | s |
|---|---|---|---|
| G-G | 6 | 502.64 | 37.57 |
| G-MF | 3 | 492.88 | 17.89 |
| G-B | 3 | 509.45 | 38.91 |
| Kruskal-Wallis Test p-Values | |||
| G-G/G-MF | G-G/G-B | G-MF/G-B | |
| −0.3651 | 0.3651 | −0.3086 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uzunidis, A. Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments. Quaternary 2021, 4, 23. https://doi.org/10.3390/quat4030023
Uzunidis A. Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments. Quaternary. 2021; 4(3):23. https://doi.org/10.3390/quat4030023
Chicago/Turabian StyleUzunidis, Antigone. 2021. "Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments" Quaternary 4, no. 3: 23. https://doi.org/10.3390/quat4030023
APA StyleUzunidis, A. (2021). Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments. Quaternary, 4(3), 23. https://doi.org/10.3390/quat4030023