
Climate and Vegetation Change, Hillslope Soil Erosion, and the Complex Nature of Late Quaternary Environmental Transitions, Eastern Mojave Desert, USA

Abstract
1. Introduction
2. Study Site and Methods
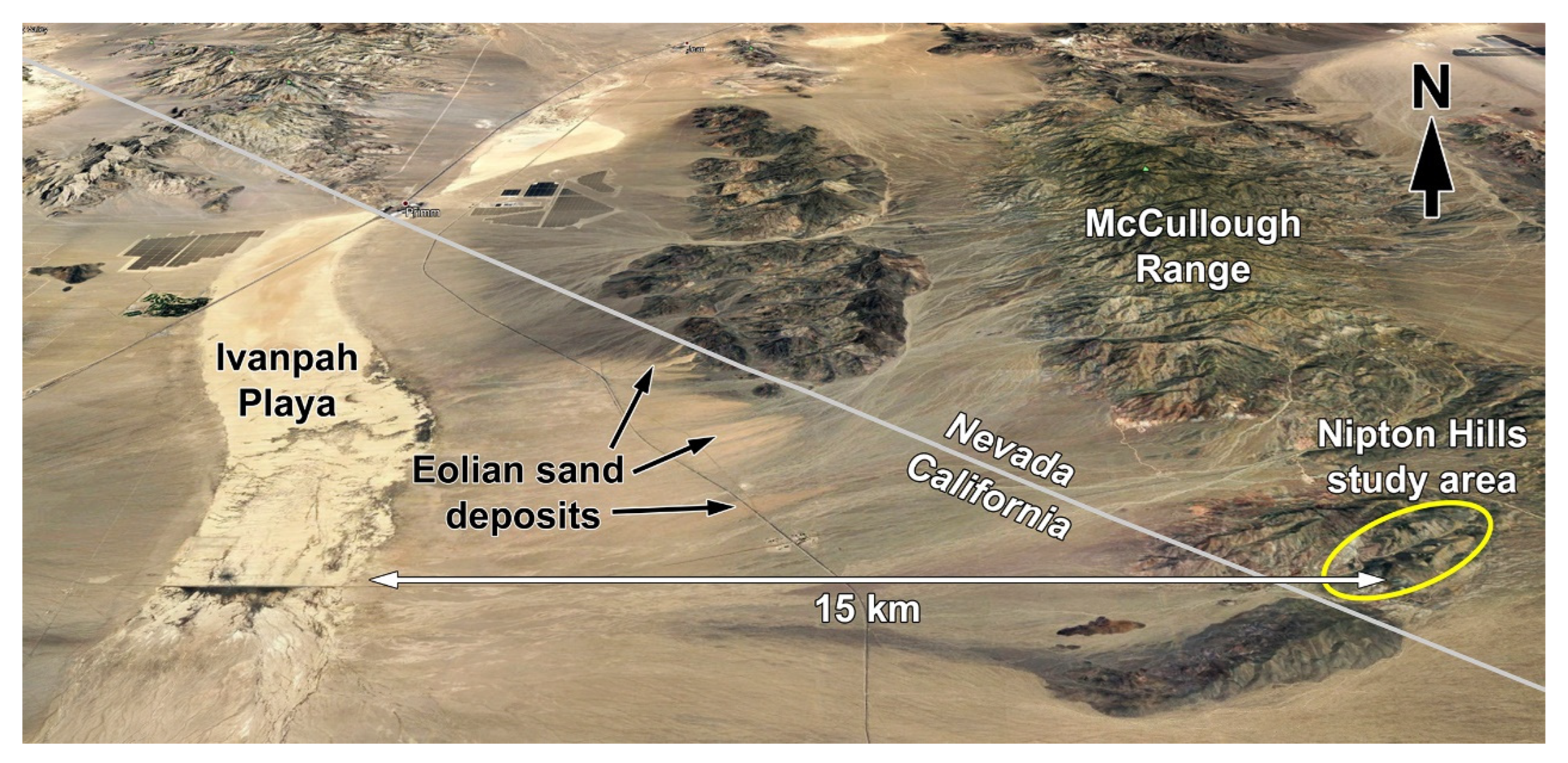
2.1. Location and Geology
2.2. Climate
2.3. Methods
3. Results
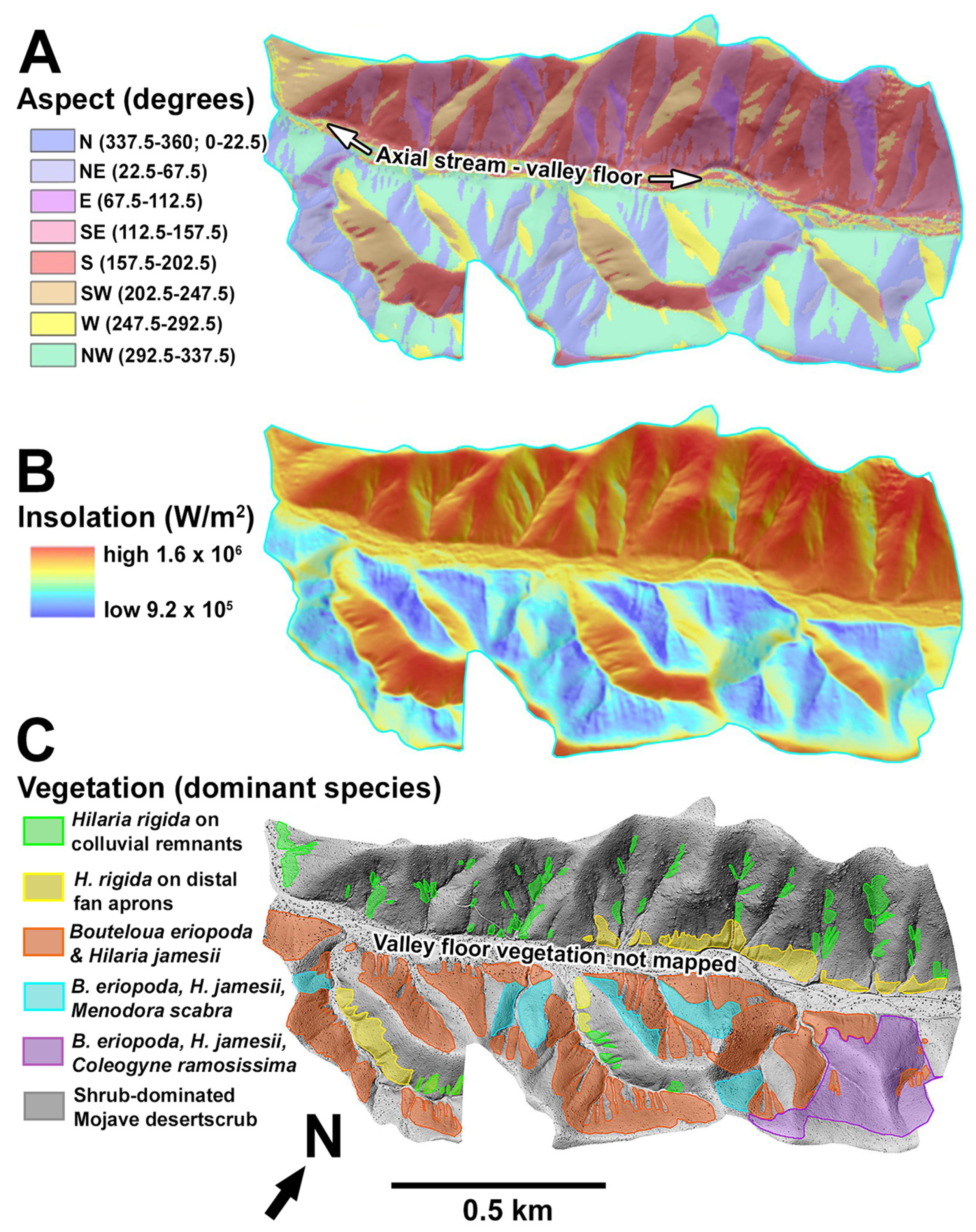
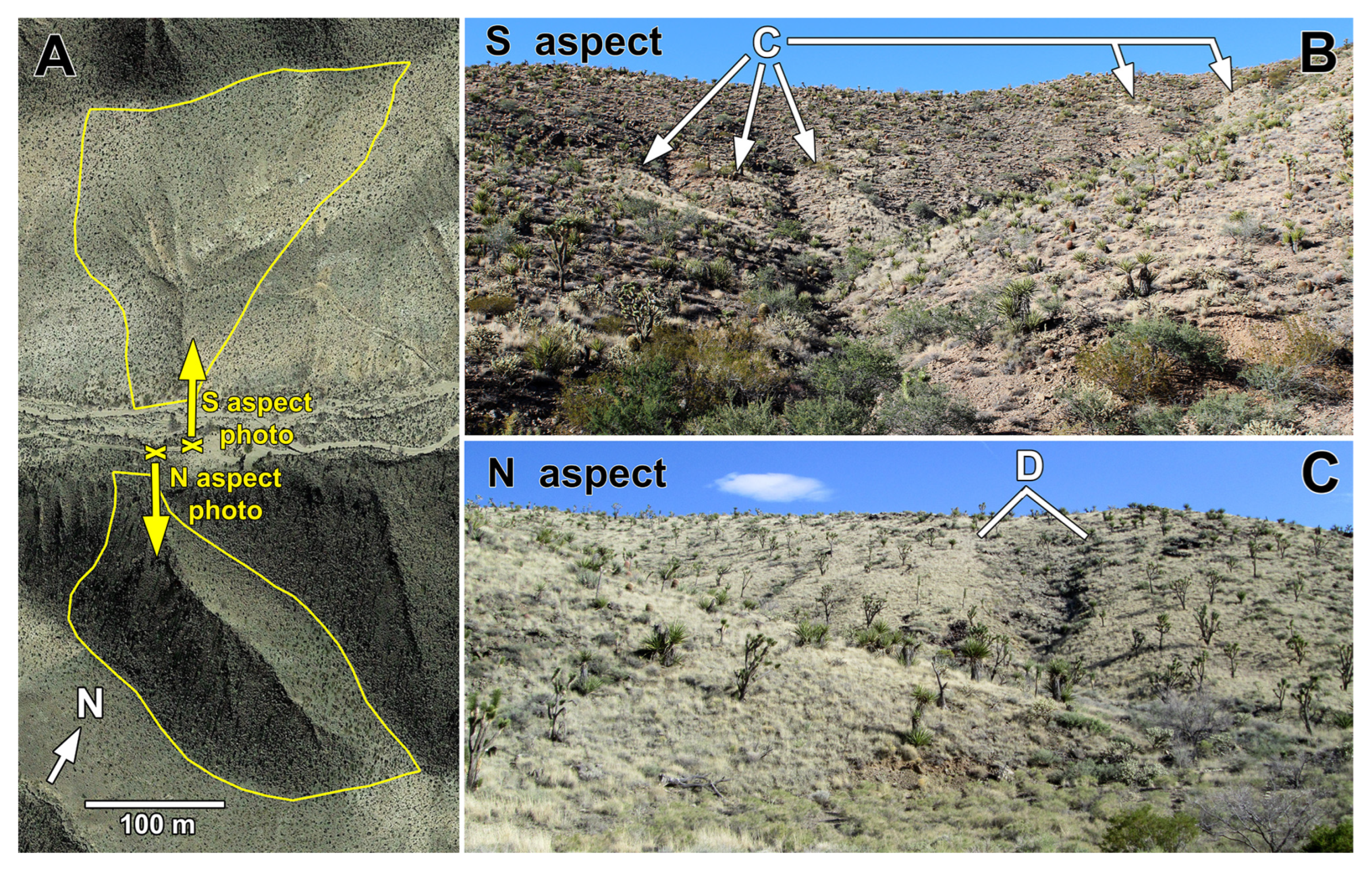
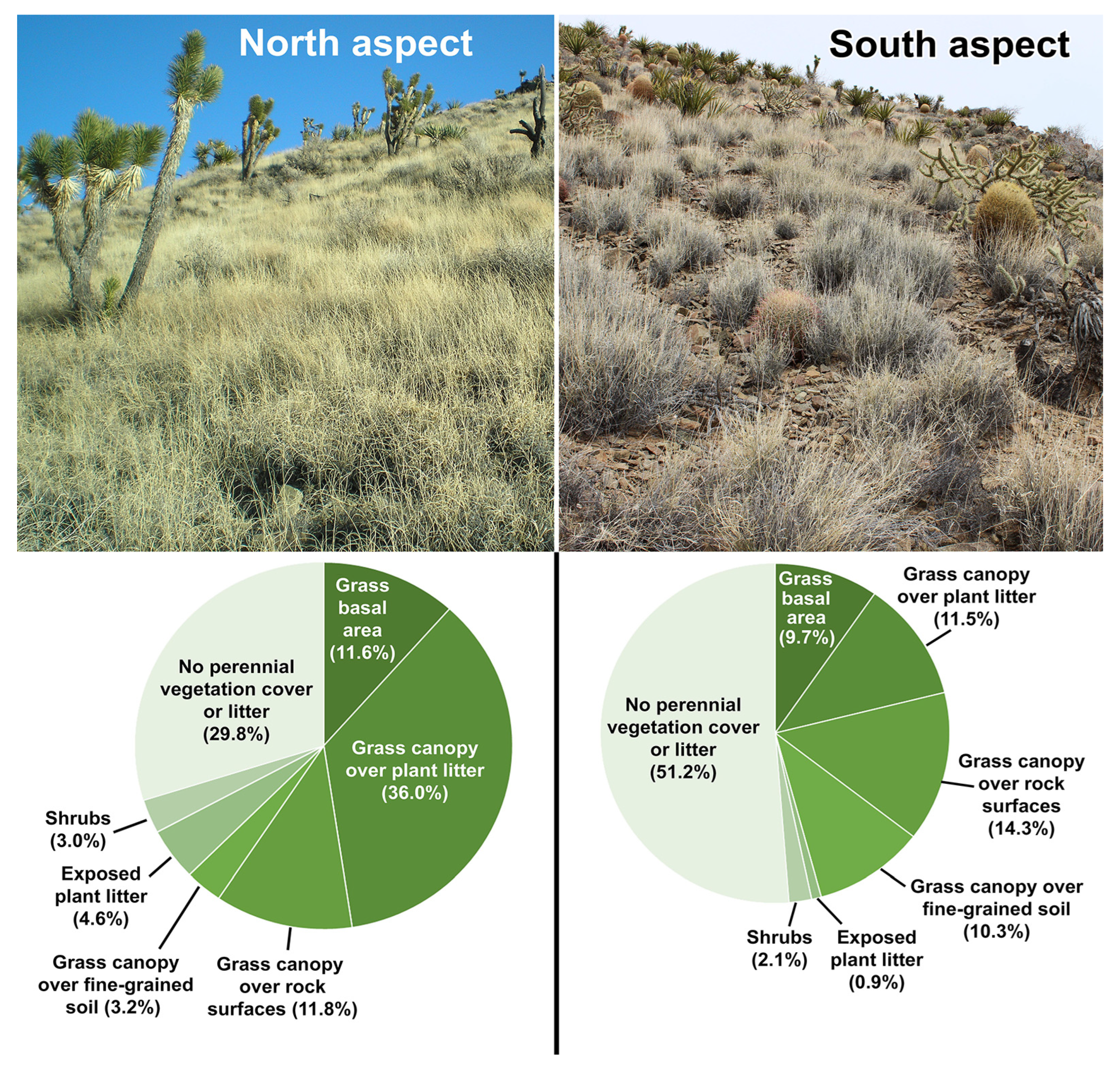
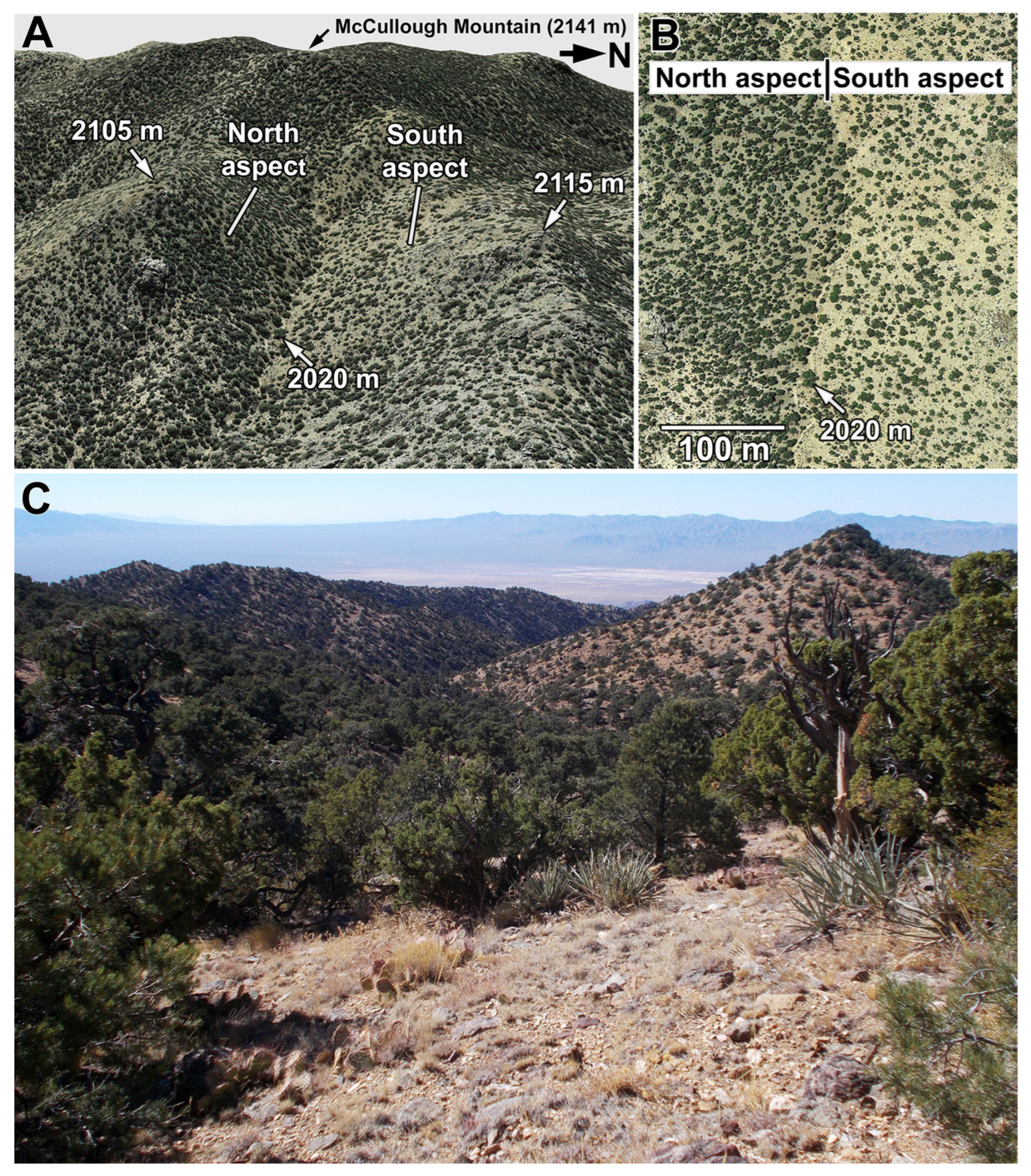
3.1. Vegetation Contrasts of Opposite Slope Aspects
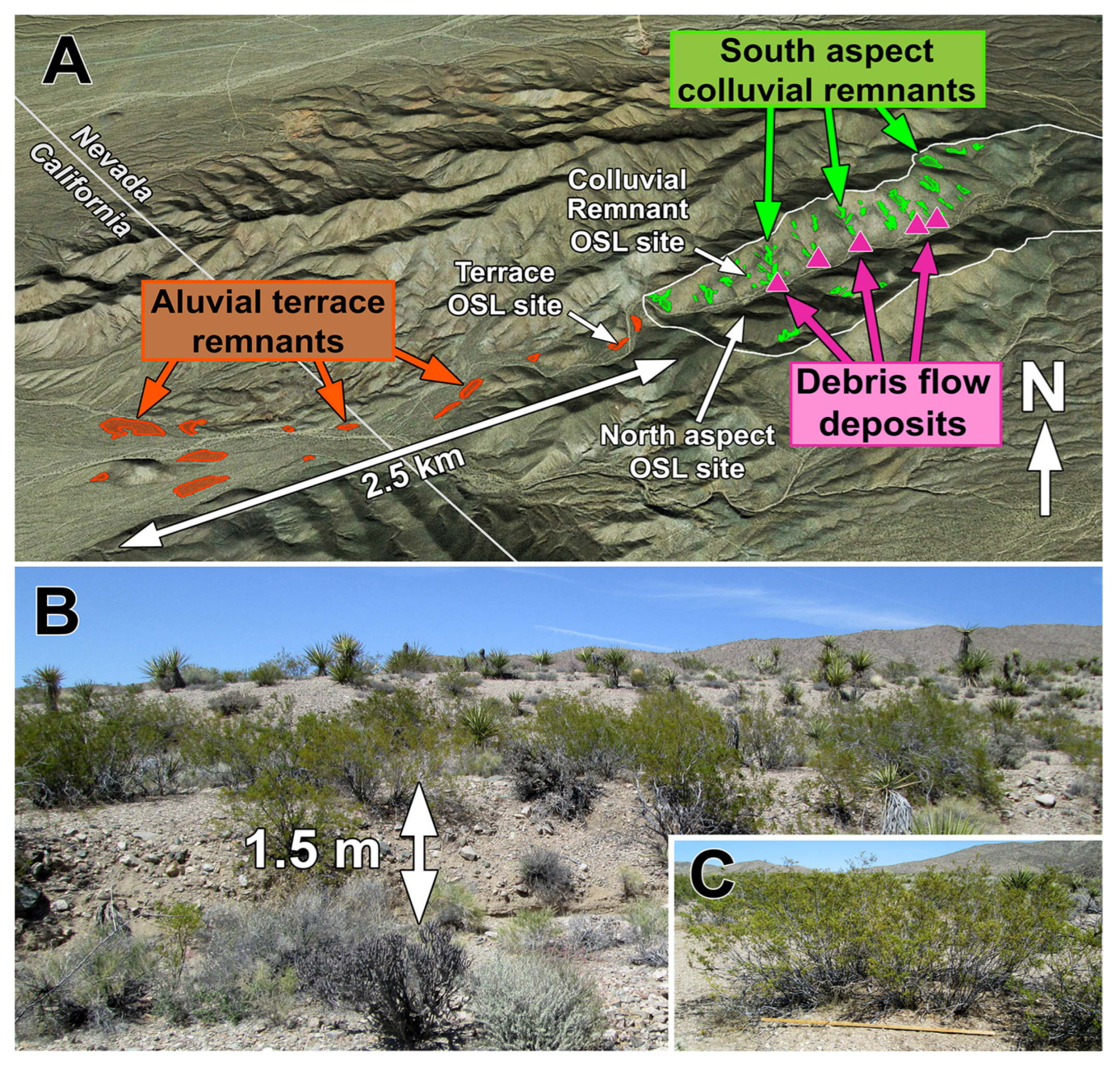
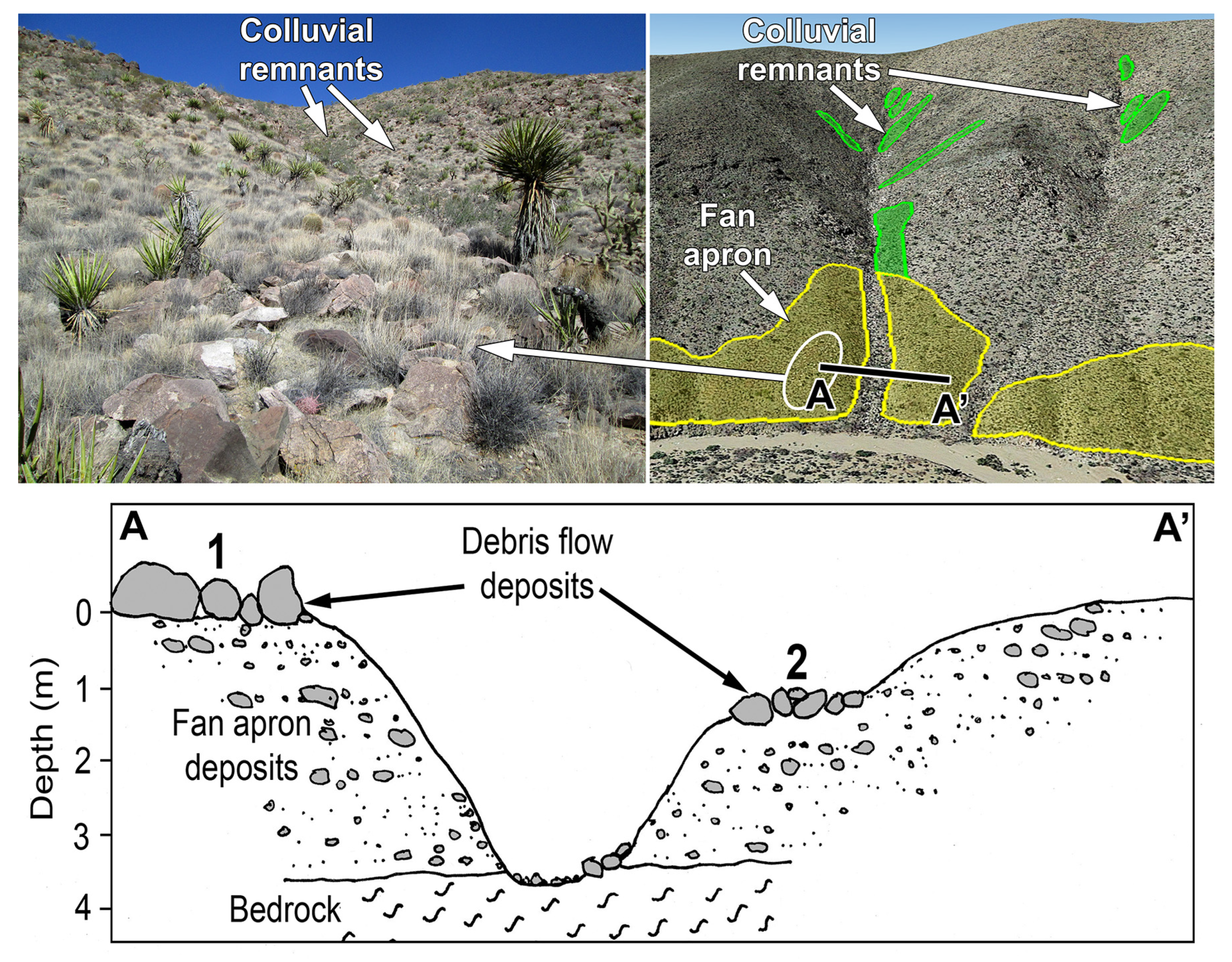
3.2. Hillslope Soils
3.2.1. Soil Characteristics
3.2.2. Formation and Age of Hillslope Soils
3.2.3. Timing of Major Episode of Hillslope Erosion
3.2.4. Extent of Hillslope Soil Erosion on South Aspects
3.2.5. Effective Ground Cover and Soil Erosion
3.3. Root Systems, the Distribution of Soil Moisture, and Soil Moisture Extraction by Plants
3.4. Modern Distributions of Predominant Perennial Grasses in Relation to Climate
4. Discussion
4.1. Reconstruction of Late Quaternary Vegetation Changes
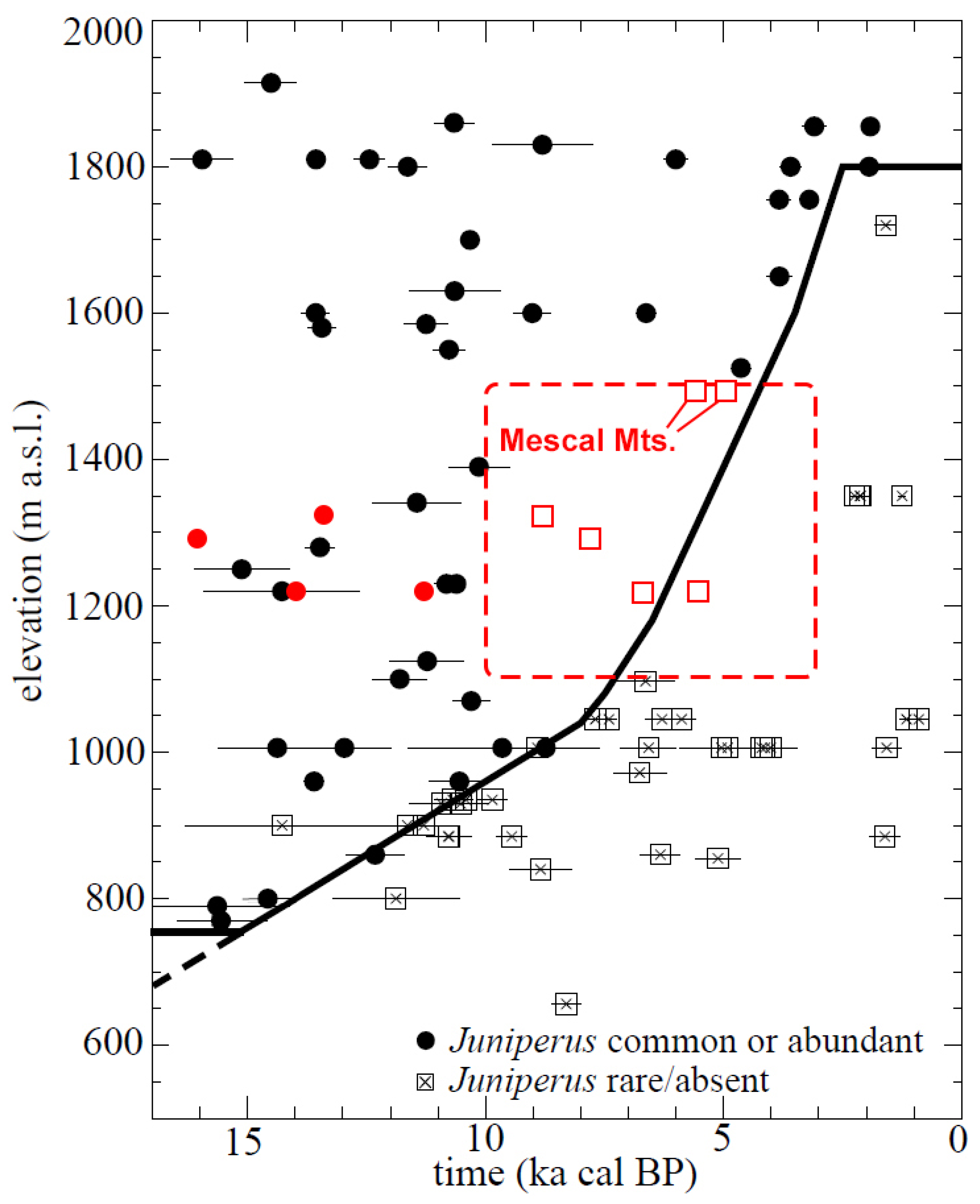
4.1.1. Late Pleistocene Vegetation at Nipton Hills
4.1.2. Initial Vegetation Transitions in the Early Holocene Driven by Climate Change
4.1.3. Subsequent Vegetation Transitions of the Middle-to Late Holocene
4.2. Climate, Vegetation Change, and Drivers of Fluvial System Behavior
4.3. Landscape Responses along Gradients of Precipitation Amount and Seasonality
4.4. Predominant Climate Drivers of Landscape Change
4.5. Effects of Future Climate Change
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, P.V.; Jorgensen, C.D. Pleistocene wood rat middens and climatic change in Mohave Desert: A record of juniper woodlands. Science 1964, 143, 1171–1173. [Google Scholar] [CrossRef] [PubMed]
- Wells, P.V.; Berger, R. Late Pleistocene history of coniferous woodland in the Mojave Desert. Science 1967, 155, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- King, T.J., Jr. Late Pleistocene—Early Holocene history of coniferous woodlands in the Lucerne Valley region, Mohave desert, California. Gr. Basin Nat. 1976, 36, 227–238. [Google Scholar]
- Betancourt, J.L.; Van Devender, T.R.; Martin, P.S. (Eds.) Packrat Middens: The Last 40,000 Years of Biotic Change; University of Arizona Press: Tucson, AZ, USA, 1990. [Google Scholar]
- Spaulding, W.G. Vegetational and climatic development of the Mojave Desert: The last glacial maximum to the present. In Packrat Middens: The Last 40,000 Years of Biotic Change; Betancourt, J.L., Van Devender, T.R., Martin, P.S., Eds.; University of Arizona Press: Tucson, AZ, USA, 1990; pp. 166–199. [Google Scholar]
- Harvey, A.M.; Wigand, P.E.; Wells, S.G. Response of alluvial fan systems to the late Pleistocene to Holocene climatic transition: Contrasts between the margins of pluvial Lakes Lahontan and Mojave, Nevada and California, USA. Catena 1999, 36, 255–281. [Google Scholar] [CrossRef]
- Pelletier, J.D. The linkages among hillslope-vegetation changes, elevation, and the timing of late-Quaternary fluvial-system aggradation in the Mojave Desert revisited. Earth Surf. Dyn. 2014, 2, 455–468. [Google Scholar] [CrossRef]
- Davis, M.B.; Woods, K.D.; Webb, S.L.; Futyma, R.P. Dispersal versus climate: Expansion of Fagus and Tsuga into the Upper Great Lakes region. Vegetatio 1986, 67, 93–103. [Google Scholar] [CrossRef]
- McAuliffe, J.R.; Van Devender, T.R. A 22,000-year record of vegetation change in the north-central Sonoran Desert. Palaeogeog. Palaeoclim. Palaeoecol. 1998, 141, 253–275. [Google Scholar] [CrossRef]
- Cole, K. Past rates of change, species richness, and a model of vegetational inertia in the Grand Canyon, Arizona. Amer. Nat. 1985, 125, 289–303. [Google Scholar] [CrossRef]
- Cole, K.L. In defense of inertia. Amer. Nat. 1986, 127, 727–728. [Google Scholar] [CrossRef]
- McFadden, L.D.; Wells, S.G.; Jercinovich, M.J. Influences of eolian and pedogenic processes on the origin and evolution of desert pavements. Geology 1987, 15, 504–508. [Google Scholar] [CrossRef]
- Adelsberger, K.A.; Smith, J.R. Desert pavement development and landscape stability on the Eastern Libyan Plateau, Egypt. Geomorphology 2009, 107, 178–194. [Google Scholar] [CrossRef]
- McDonald, E.V.; Pierson, F.B.; Flerchinger, G.N.; McFadden, L.D. Application of a soil-water balance model to evaluate the influence of Holocene climate change on calcic soils, Mojave Desert, California, USA. Geoderma 1996, 74, 167–192. [Google Scholar] [CrossRef]
- Meadows, D.G.; Young, M.H.; McDonald, E.V. Influence of relative surface age on hydraulic properties and infiltration on soils associated with desert pavements. Catena 2008, 72, 169–178. [Google Scholar] [CrossRef]
- McAuliffe, J.R.; McDonald, E.V. Holocene environmental change and vegetation contraction in the Sonoran Desert. Quat. Res. 2006, 65, 204–215. [Google Scholar] [CrossRef]
- McAuliffe, J.R. Soil horizon development and the tempo and modes of vegetation change during the Holocene in a Sonoran Desert basin, USA. Holocene 2019, 29, 1263–1272. [Google Scholar] [CrossRef]
- McAuliffe, J.R. The Sonoran Desert: Landscape complexity and ecological diversity. In Ecology of Sonoran Desert Plants and Plant Communities; Robichaux, R., Ed.; University of Arizona Press: Tucson, AZ, USA, 1999; pp. 87–104. [Google Scholar]
- McAuliffe, J.R. Desert soils. In A Natural History of the Sonoran Desert, 2nd ed.; Dimmitt, M.A., Comas, P.W., Brewer, L.M., Eds.; Arizona-Sonora Desert Museum Press & Univ. of California Press: Berkeley, CA, USA, 2015; pp. 85–100. [Google Scholar]
- McAuliffe, J.R.; Sundt, P.C.; Valiente-Banuet, A.; Casas, A.; Viveros, J.L. Pre-Columbian soil erosion, persistent ecological changes, and collapse of a subsistence agricultural economy in the semi-arid Tehuacán Valley, Mexico’s “Cradle of Maize”. J. Arid Environ. 2001, 47, 47–75. [Google Scholar] [CrossRef]
- McAuliffe, J.R.; McFadden, L.D.; Roberts, L.M.; Wawrzyniec, T.F.; Scuderi, L.A.; Meyer, G.A.; King, M.P. Non-equilibrium hillslope dynamics and irreversible landscape changes at a shifting pinyon-juniper woodland. Glob. Planet. Chang. 2014, 122, 1–13. [Google Scholar] [CrossRef]
- Persico, L.; McFadden, L.; McAuliffe, J.; Rittenour, T.; Stahlecker, T.; Dunn, S.; Brody, W. Late Quaternary geochronologic record of soil formation and erosion: Effects of climate change on Mojave Desert hillslopes. Geology 2021, 50, 54–59. [Google Scholar] [CrossRef]
- Miller, D.M. Surficial geologic map of the Ivanpah 30′ × 60′ Quadrangle, San Bernardino County, California, and Clark County, Nevada: U.S. Geological Survey Scientific Investigations Map 3206, Scale 1:100,000; U.S. Geological Survey: Menlo Park, CA, USA, 2012; 14p.
- McAuliffe, J.R. Perennial Grass-Dominated Plant Communities of the Eastern Mojave Desert Region. Desert Plants 2016, 32, 1–90. Available online: https://repository.arizona.edu/handle/10150/622004 (accessed on 1 January 2020).
- Rowlands, P.; Johnson, H.; Ritter, E.; Endo, A. The Mojave desert. In Reference Handbook on the Deserts of North America; Bender, G.L., Ed.; Greenwood Press: Westport, London, UK, 1982; pp. 103–162. [Google Scholar]
- Corbosiero, K.L.; Dickinson, M.J.; Bosart, L.F. The contribution of eastern North Pacific tropical cyclones to the rainfall climatology of the southwest United States. Mon. Weather Rev. 2009, 137, 2415–2435. [Google Scholar] [CrossRef]
- Ritchie, E.A.; Wood, K.M.; Gutzler, D.S.; White, S.R. The influence of eastern Pacific tropical cyclone remnants on the southwestern United States. Mon. Weather Rev. 2011, 139, 192–210. [Google Scholar] [CrossRef]
- Walker, M.; Head, M.J.; Lowe, J.; Berkelhammer, M.; Björck, S.; Cheng, H.; Cwynar, L.C.; Fisher, D.; Gkinis, V.; Long, A.; et al. Subdividing the Holocene Series/Epoch: Formalization of stages/ages and subseries/subepochs, and designation of GSSPs and auxiliary stratotypes. J. Quat. Sci. 2019, 34, 173–186. [Google Scholar] [CrossRef]
- Gile, L.H.; Peterson, F.F.; Grossman, R.B. Morphological and genetic sequences of carbonate accumulation in desert soils. Soil Sci. 1966, 101, 347–360. [Google Scholar] [CrossRef]
- Reheis, M.C.; Goodmacher, J.C.; Harden, J.W.; McFadden, L.D.; Rockwell, T.K.; Shroba, R.R.; Sowers, J.M.; Taylor, E.M. Quaternary soils and dust deposition in southern Nevada and California. Geol. Soc. Amer. Bull. 1995, 107, 1003–1022. [Google Scholar] [CrossRef]
- Ugolini, F.C.; Hillier, S.; Certini, G.; Wilson, M.J. The contribution of aeolian material to an Aridisol from southern Jordan as revealed by mineralogical analysis. J. Arid Environ. 2008, 72, 1431–1447. [Google Scholar] [CrossRef]
- Hirmas, D.R.; Graham, R.C.; Kendrick, K.J. Pedogenesis and soil-geomorphic relationships in an arid mountain range, Mojave Desert, California. Soil Sci. Soc. Amer. J. 2011, 75, 192–206. [Google Scholar] [CrossRef]
- Hirmas, D.R.; Graham, R.C.; Kendrick, K.J. Soil-geomorphic significance of lands surface characteristics in an arid mountain range, Mojave Desert, USA. Catena 2011, 87, 408–420. [Google Scholar] [CrossRef]
- Persico, L.P.; McFadden, L.D.; Frechette, J.D.; Meyer, G.A. Rock type and dust influx control accretionary soil development on hillslopes in the Sandia Mountains, New Mexico, USA. Quat. Res. 2011, 76, 411–416. [Google Scholar] [CrossRef]
- Goossens, D. Field experiments of aeolian dust accumulation on rock fragment substrata. Sedimentology 1995, 42, 391–402. [Google Scholar] [CrossRef]
- Blank, R.R.; Young, J.A.; Lugaski, T. Pedogenesis on talus slopes, the Buckskin range, Nevada, USA. Geoderma 1996, 71, 121–142. [Google Scholar] [CrossRef]
- Chadwick, O.A.; Davis, J.O. Soil-forming intervals caused by eolian sediment pulses in the Lahontan basin, northwestern Nevada. Geology 1990, 18, 243–246. [Google Scholar] [CrossRef]
- Reheis, M.C.; Kihl, R. Dust deposition in southern Nevada and California, 1984–1989: Relations to climate, source area, and source lithology. J. Geophys. Res. Atmos. 1995, 100, 8893–8918. [Google Scholar] [CrossRef]
- Sweeney, M.R.; McDonald, E.V.; Etyemezian, V. Quantifying dust emissions from desert landforms, eastern Mojave Desert, USA. Geomorphology 2011, 135, 21–34. [Google Scholar] [CrossRef]
- McDonald, E.V.; McFadden, L.D.; Wells, S.G.; Enzel, Y.; Lancaster, N. Regional response of alluvial fans to the Pleistocene-Holocene climatic transition, Mojave Desert, California. In Paleoenvironments and Paleohydrology of the Mojave and Southern Great Basin Deserts; Enzel, Y., Wells, S.G., Lancaster, N., Eds.; Geological Society of America: Boulder, CO, USA, 2003; Volume 368, pp. 189–205. [Google Scholar]
- Yan, Y.; Xu, X.; Xin, X.; Yang, G.; Wang, X.; Yan, R.; Chen, B. Effect of vegetation coverage on aeolian dust accumulation in a semiarid steppe of northern China. Catena 2011, 87, 351–356. [Google Scholar] [CrossRef]
- Vasek, F.C. Creosote bush: Long-lived clones in the Mojave Desert. Am. J. Bot. 1980, 67, 246–255. [Google Scholar] [CrossRef]
- McAuliffe, J.R.; Hamerlynck, E.P.; Eppes, M.C. Landscape dynamics fostering the development and persistence of long-lived creosotebush (Larrea tridentata) clones in the Mojave Desert. J. Arid Environ. 2007, 69, 96–126. [Google Scholar] [CrossRef]
- Helms, J.G.; McGill, S.F.; Rockwell, T.K. Calibrated, late Quaternary age indices using clast rubification and soil development on alluvial surfaces in Pilot Knob Valley, Mojave Desert, southeastern California. Quat. Res. 2003, 60, 377–393. [Google Scholar] [CrossRef]
- McAuliffe, J.R. The interface between precipitation and vegetation: The importance of soils in arid and semiarid environments. In Changing Precipitation Regimes and Terrestrial Ecosystems: A North American Perspective; Weltzin, J., McPherson, G.R., Eds.; University of Arizona Press: Tucson, AZ, USA, 2003; pp. 9–27. [Google Scholar]
- Sternberg, P.D.; Anderson, M.A.; Graham, R.C.; Beyers, J.L.; Tice, K.R. Root distribution and seasonal water status in weathered granitic bedrock under chaparral. Geoderma 1996, 72, 89–98. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Witty, J.H.; Graham, R.C.; Hubbert, K.R.; Doolittle, J.A.; Wald, J.A. Contributions of water supply from the weathered bedrock zone to forest soil quality. Geoderma 2003, 114, 389–400. [Google Scholar] [CrossRef]
- McCormick, E.L.; Dralle, D.N.; Hahm, W.J.; Tune, A.K.; Schmidt, L.M.; Chadwick, K.D.; Rempe, D.M. Widespread Woody Plant Use of Water Stored in Bedrock. Nature 2021, 597, 225–229. Available online: https://www.fs.fed.us/psw/publications/dralle/psw2021dralle005mccormick.pdf (accessed on 1 January 2020). [CrossRef] [PubMed]
- Sala, O.E.; Lauenroth, W.K.; Golluscio, R.A. Plant functional types in temperate semi-arid areas. In Plant Functional Types; Smith, T.M., Shugart, H.H., Woodward, F.I., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 217–233. [Google Scholar]
- Schenk, H.J.; Jackson, R.B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef]
- Holmgren, C.A.; Betancourt, J.L.; Rylander, K.A. A long-term vegetation history of the Mojave–Colorado desert ecotone at Joshua Tree National Park. J. Quat. Sci. 2010, 25, 222–236. [Google Scholar] [CrossRef]
- Koehler, P.A.; Anderson, R.S.; Spaulding, W.G. Development of vegetation in the central Mojave Desert of California during the late Quaternary. Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 215, 297–311. [Google Scholar] [CrossRef]
- Longwell, C.R.; Pampeyan, E.H.; Bowyer, B.; Roberts, R.J. Geology and Mineral Deposits of Clark County, Nevada; Nevada Bureau of Mines and Geology Bulletin 62; Mackay School of Mines, University of Nevada-Reno: Reno, NV, USA, 1965. [Google Scholar]
- Spaulding, W.G. A middle Holocene vegetation record from the Mojave Desert of North America and its paleoclimatic significance. Quat. Res. 1991, 35, 427–437. [Google Scholar] [CrossRef]
- Steponaitis, E.; Andrews, A.; McGee, D.; Quade, J.; Hsieh, Y.T.; Broecker, W.S.; Shuman, B.N.; Burns, S.J.; Cheng, H. Mid-Holocene drying of the US Great Basin recorded in Nevada speleothems. Quat. Sci. Rev. 2015, 127, 174–185. [Google Scholar] [CrossRef]
- Bull, W.B. Geomorphic Responses to Climate Change; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Antinao, J.L.; McDonald, E. A reduced relevance of vegetation change for alluvial aggradation in arid zones. Geology 2013, 41, 11–14. [Google Scholar] [CrossRef]
- Enzel, Y.; Amit, R.; Grodek, T.; Ayalon, A.; Lekach, J.; Porat, N.; Bierman, P.; Blum, J.D.; Erel, Y. Late Quaternary weathering, erosion, and deposition in Nahal Yael, Israel: An “impact of climatic change on an arid watershed”? Geol. Soc. Amer. Bull. 2012, 124, 705–722. [Google Scholar] [CrossRef]
- Antinao, J.L.; McDonald, E. An enhanced role for the Tropical Pacific on the humid Pleistocene–Holocene transition in southwestern North America. Quat. Sci. Rev. 2013, 78, 319–341. [Google Scholar] [CrossRef]
- Johnson, H.B. Vegetation and plant communities of southern California deserts—A functional view. In Plant Communities of Southern California; Latting, J., Ed.; Special Publication No. 2; California Native Plant Society: Sacramento, CA, USA, 1976; pp. 125–164. [Google Scholar]
- Schlesinger, W.H.; Reynolds, J.F.; Cunningham, G.L.; Huenneke, L.F.; Jarrell, W.M.; Virginia, R.M.; Whitford, W.G. Biological feedbacks in global desertification. Science 1990, 247, 1043–1048. [Google Scholar] [CrossRef]
- Abrahams, A.D.; Parsons, A.J.; Wainwright, J. Effects of vegetation change on interrill runoff and erosion, Walnut Gulch, southern Arizona. Geomorphology 1995, 13, 37–48. [Google Scholar] [CrossRef]
- Gutiérrez-Jurado, H.A.; Vivoni, E.R.; Cikowski, C.; Harrison, J.B.; Bras, R.L.; Istanbulluoglu, E. On the observed ecohydrological dynamics of a semiarid basin with aspect delimited ecosystems. Water Resour. Res. 2013, 49, 8263–8284. [Google Scholar] [CrossRef]
- Wells, S.G.; McFadden, L.D.; Dohrenwend, J.C. Influence of late Quaternary climatic changes on geomorphic and pedogenic processes on a desert piedmont, eastern Mojave Desert, California. Quat. Res. 1987, 27, 130–146. [Google Scholar] [CrossRef]
- Miller, D.M.; Schmidt, K.M.; Mahan, S.A.; McGeehin, J.P.; Owen, L.A.; Barron, J.A.; Lehmkuhl, F.; Löhrer, R. Holocene landscape response to seasonality of storms in the Mojave Desert. Quat. Internat. 2010, 215, 45–61. [Google Scholar] [CrossRef]
- Rein, B.; Lückge, A.; Reinhardt, L.; Sirocko, F.; Wolf, A.; Dullo, W.C. El Niño variability off Peru during the last 20,000 years. Paleoceanography 2005, 20, PA4003. Available online: https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/2004PA001099 (accessed on 1 January 2022). [CrossRef]
- Moy, C.M.; Seltzer, G.O.; Rodbell, D.T.; Anderson, D.M. Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature 2002, 420, 162–165. [Google Scholar] [CrossRef]
- Jin, F.F.; Boucharel, J.; Lin, I.I. Eastern Pacific tropical cyclones intensified by El Niño delivery of subsurface ocean heat. Nature 2014, 516, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Hereford, R.; Webb, R.H. Historic variation of warm-season rainfall, southern Colorado Plateau, southwestern USA. Clim. Chang. 1992, 22, 239–256. [Google Scholar] [CrossRef]
- McAuliffe, J.R.; Scuderi, L.A.; McFadden, L.D. Tree-ring record of hillslope soil erosion and valley floor dynamics: Landscape responses to climate variation during the last 400 years in the Colorado Plateau, northeastern Arizona. Glob. Planet. Chang. 2006, 50, 184–201. [Google Scholar] [CrossRef]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehman, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| North Aspect | South Aspect | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Thick Colluvial Mantle | Thin Colluvial Mantle | Older Colluvial Remnant | Distal Fan Apron | ||||||
| Sample Sites: | N1 | N2 | N3 | S1 | S2 | S3 | S4 | S5 | S6 |
| Perennial grass species | |||||||||
| Hilaria jamesii | 67 | 41 | 54 | 42 | |||||
| Bouteloua eriopoda | 15 | 16 | 3 | 3 | |||||
| Hilaria rigida | 43 | 45 | 48 | ||||||
| Muhlenbergia porteri | 2 | ||||||||
| Tridens muticus | 3 | ||||||||
| Dasyochloa pulchella | 1 | 1 | |||||||
| Total % cover (perennial grasses) | 67 | 58 | 70 | 1 | 4 | 46 | 45 | 42 | 51 |
| Shrub species | |||||||||
| Menodora spinescens | + | + | |||||||
| Ephedra nevadensis | 2 | 3 | + | 1 | 4 | 4 | |||
| Eriogonum fasciculatum | 9 | 6 | |||||||
| Encelia virginensis | 1 | 7 | |||||||
| Krameria erecta | 2 | 3 | 4 | 4 | |||||
| Acacia [= Senegalia] greggii | 1 | 1 | |||||||
| Larrea tridentata | 2 | ||||||||
| Aloysia wrightii | 2 | + | + | ||||||
| Adenophyllum cooperi | 4 | ||||||||
| Coleogyne ramosissima | 2 | ||||||||
| Total % cover (shrubs) | 2 | 3 | <1 | 20 | 21 | 8 | <1 | 8 | <1 |
| Succulent species | |||||||||
| Cylindropuntia ramosissima | + | 9 | + | + | + | + | + | ||
| Ferocactus cylindraceus | 1 | 1 | + | + | + | + | |||
| Yucca brevifolia var. jaegeriana | + | + | + | ||||||
| Yucca schidigera | 2 | 2 | + | + | + | + | |||
| Yucca baccata | + | 1 | |||||||
| Total % cover (succulents) | <1 | <1 | <1 | 13 | 3 | <1 | <1 | <1 | <1 |
| Total % Perennial Plant Cover | 69 | 61 | 70 | 34 | 28 | 54 | 45 | 50 | 51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McAuliffe, J.R.; McFadden, L.D.; Persico, L.P.; Rittenour, T.M. Climate and Vegetation Change, Hillslope Soil Erosion, and the Complex Nature of Late Quaternary Environmental Transitions, Eastern Mojave Desert, USA. Quaternary 2022, 5, 43. https://doi.org/10.3390/quat5040043
McAuliffe JR, McFadden LD, Persico LP, Rittenour TM. Climate and Vegetation Change, Hillslope Soil Erosion, and the Complex Nature of Late Quaternary Environmental Transitions, Eastern Mojave Desert, USA. Quaternary. 2022; 5(4):43. https://doi.org/10.3390/quat5040043
Chicago/Turabian StyleMcAuliffe, Joseph R., Leslie D. McFadden, Lyman P. Persico, and Tammy M. Rittenour. 2022. "Climate and Vegetation Change, Hillslope Soil Erosion, and the Complex Nature of Late Quaternary Environmental Transitions, Eastern Mojave Desert, USA" Quaternary 5, no. 4: 43. https://doi.org/10.3390/quat5040043
APA StyleMcAuliffe, J. R., McFadden, L. D., Persico, L. P., & Rittenour, T. M. (2022). Climate and Vegetation Change, Hillslope Soil Erosion, and the Complex Nature of Late Quaternary Environmental Transitions, Eastern Mojave Desert, USA. Quaternary, 5(4), 43. https://doi.org/10.3390/quat5040043