


Preservation of Mammalian Teeth and Bones Influences Identification of Terminal Pleistocene to Middle Holocene Hunter-Gatherer Subsistence at Ban Rai Rockshelter, Northwest Thailand

Abstract
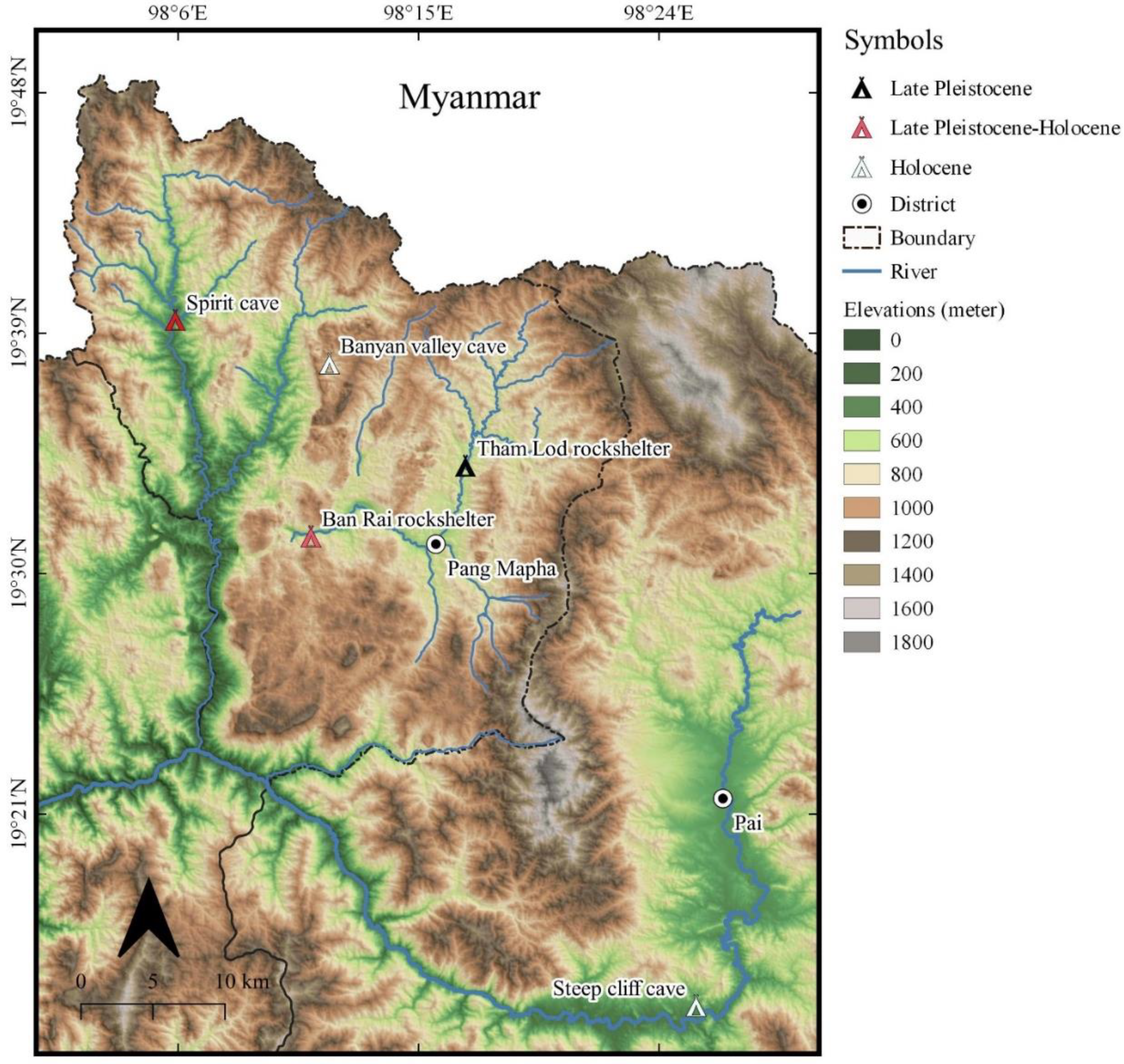
:1. Introduction
2. Site Excavation, Chronology, and Stratigraphy
2.1. The First Cultural Phase
2.2. The Second Cultural Phase
3. Materials and Methods
3.1. Study Materials
3.2. Zooarchaeological Methods
4. Results
4.1. Taxonomic Composition
4.2. Taphonomic Insights
5. Discussion
5.1. Sample Size Influence, Tooth Preservation, and Area 3
5.2. Long-Term Subsistence Change
5.3. Changes in Paleoenvironments and Paleofaunas
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conrad, C. Mainland Southeast Asia in the Longue Durée: A Zooarchaeological Test of the “Broad Spectrum Revolution” in Northern Thailand. Ph.D. Thesis, The University of New Mexico, Albuquerque, New Mexico, 2018. [Google Scholar]
- Conrad, C.; Higham, C.; Eda, M.; Marwick, B. Palaeoecology and forager subsistence strategies during the Pleistocene-Holocene transition: A reinvestigation of the zooarchaeological assemblage from Spirit Cave, Mae Hong Son Province, Thailand. Asian Perspect. 2016, 55, 2–27. [Google Scholar] [CrossRef]
- Gorman, C.F. Excavations at Spirit Cave, North Thailand: Some Interim Interpretations. Asian Perspect. 1970, 13, 79–107. [Google Scholar]
- Reynolds, T.E.G. Excavations at Banyan Valley Cave, Northeast Thailand: A Report on the 1972 Season. Asian Perspect. 1992, 31, 77–97. [Google Scholar]
- Shoocongdej, R. Paleoenvironment during Late Pleistocene to Late Holocene in highland Pang Mapha District, Mae Hong Son Province. In The Proceeding of the Conference on People, Culture, and Paleoenvironment in Highland Pang Mapha, Mae Hong Son Province, Silpakorn University (Tha Phra Palace), 20’21 February 2003; Silpakorn University: Bangkok, Thailand, 2003; pp. 292–317. (In Thai) [Google Scholar]
- Shoocongdej, R. Late Pleistocene activities at the Tham Lod rockshelter in highland Pang Mapha, Mae Hong Son province, northwestern Thailand. In Uncovering Southeast Asia’s Past: Selected Papers from the 10th International Conference of the European Association of Southeast Asian Archaeologists, the British Museum, London, 14’17 September 2004; Bacus, E.A., Glover, I.C., Pigott, V., Eds.; NUS Press: Singapore, 2006; pp. 22–37. [Google Scholar]
- Shoocongdej, R.; Phumijumnong, N.; Chintakanon, K.; Pureepatpong, N.; Hoontrakul, U.; Treerayapiwat, C. Final Report of Highland Archaeology in Pang Mapha Distric, Mae Hong Son Province Phase 2, 2007; Volume 2. Archaeology, Submitted to Thailand Research Fund (TRF) (in Thai)
- Shoocongdej, R.; Wattanapituksakul, A. Faunal assemblages and demography during the Late Pleistocene (MIS 2-1) to Early Holocene in Highland Pang Mapha, Northwest Thailand. Quat. Int. 2020, 563, 51–63. [Google Scholar] [CrossRef]
- Treerayapiwat, C. Patterns of habitation and burial activity in the Ban Rai rockshelter, northwestern Thailand. Asian Perspect. 2005, 44, 231–245. [Google Scholar] [CrossRef]
- Conrad, C.; Shoocongdej, R.; Marwick, B.; White, J.C.; Thongcharoenchaikit, C.; Higham, C.; Feathers, J.K.; Tumpeesuwan, S.; Castillo, C.C.; Fuller, D.Q.; et al. Re-evaluating Pleistocene–Holocene occupation of cave sites in north-west Thailand: New radiocarbon and luminescence dating. Antiquity 2021, 96, 280–297. [Google Scholar] [CrossRef]
- Gorman, C. The Hoabinhian and After: Subsistence Patterns in Southeast Asia during the Late Pleistocene and Early Recent Periods. World Archaeol. 1971, 2, 300–320. [Google Scholar] [CrossRef]
- Suraprasit, K.; Jaeger, J.-J.; Shoocongdej, R.; Chaimanee, Y.; Wattanapituksakul, A.; Bocherens, H. Long-Term Isotope Evidence on the Diet and Habitat Breadth of Pleistocene to Holocene Caprines in Thailand: Implications for the Extirpation and Conservation of Himalayan Gorals. Front. Ecol. Evol. 2020, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Suraprasit, K.; Shoocongdej, R.; Chintakanon, K.; Bocherens, H. Late Pleistocene human paleoecology in the highland savanna ecosystem of mainland Southeast Asia. Sci. Rep. 2021, 11, 16756. [Google Scholar] [CrossRef]
- Wattanapituksakul, A.; Filoux, A.; Amphansri, A.; Tumpeesuwan, S. Late Pleistocene Caprinae assemblages of Tham Lod rockshelter (Mae Hong Son province, northwest Thailand). Quat. Int. 2018, 493, 212–226. [Google Scholar] [CrossRef]
- Bacon, A.-M.; Duringer, P.; Westaway, K.; Joannes-Boyau, R.; Zhao, J.; Bourgon, N.; Dufour, E.; Pheng, S.; Tep, S.; Ponche, J.-L.; et al. Testing the savannah corridor hypothesis during MIS2: The Boh Dambang hyena site in southern Cambodia. Quat. Int. 2018, 464, 417–439. [Google Scholar] [CrossRef]
- Heaney, L.R. A synopsis of climatic and vegetational change in Southeast Asia. Clim. Change 1991, 19, 53–61. [Google Scholar] [CrossRef]
- Louys, J.; Roberts, P. Environmental drivers of megafauna and hominin extinction in Southeast Asia. Nature 2020, 586, 402–406. [Google Scholar] [CrossRef]
- Suraprasit, K.; Jongautchariyakul, S.; Yamee, C.; Pothichaiya, C.; Bocherens, H. New fossil and isotope evidence for the Pleistocene zoogeographic transition and hypothesized savanna corridor in peninsular Thailand. Quat. Sci. Rev. 2019, 221, 105861. [Google Scholar] [CrossRef]
- Wurster, C.M.; Bird, M.I.; Bull, I.D.; Creed, F.; Bryant, C.; Dungait, J.A.J.; Paz, V. Forest contraction in north equatorial Southeast Asia during the Last Glacial Period. Proc. Natl. Acad. Sci. 2010, 107, 15508–15511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kealhofer, L.; Penny, D. A combined pollen and phytolith record for fourteen thousand years of vegetation change in northeastern Thailand. Rev. Palaeobot. Palynol. 1998, 103, 83–93. [Google Scholar] [CrossRef]
- Penny, D. A 40,000 year palynological record from north-east Thailandl; implications for biogeography and palaeo-environmental reconstruction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 171, 97–128. [Google Scholar] [CrossRef]
- Sun, X.; Li, X.; Luo, Y.; Chen, X. The vegetation and climate at the last glaciation on the emerged continental shelf of the South China Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 160, 301–316. [Google Scholar] [CrossRef]
- Bien, M.N.; Chia, L.P. Cave and rock-shelter deposits in Yunnan. Bull. Geol. Soc. China 1938, 18, 325–347. [Google Scholar] [CrossRef]
- Von Koenigswald, G. The relationship between the fossil mammalian faunae of Java and China, with special reference to early man. Peking Nat. Hist. Bul. 1939, 13, 293–298. [Google Scholar]
- Kahlke, H.D. On the complex of the Stegodon-Ailuropoda-fauna of southern China and the chronological position of Gigantopithecus blacki V. Koenigswald. Vert. PalAs. 1961, 6, 83–108. [Google Scholar]
- Louys, J. Quaternary extinctions in Southeast Asia. In Mass Extinction; Springer: Berlin, Germany, 2007; pp. 159–189. [Google Scholar]
- Suraprasit, K.; Jaeger, J.-J.; Chaimanee, Y.; Chavasseau, O.; Yamee, C.; Tian, P.; Panda, S. The Middle Pleistocene vertebrate fauna from Khok Sung (Nakhon Ratchasima, Thailand): Biochronological and paleobiogeograpgical implications. Zookeys 2016, 613, 1–157. [Google Scholar]
- Bacon, A.-M.; Westaway, K.; Antoine, P.O.; Duringer, P.; Blin, A.; Demeter, F.; Ponche, J.-L.; Zhao, J.-X.; Barnes, L.; Sayavongkhamdy, T.; et al. Late Pleistocene mammalian assemblages of Southeast Asia: New dating, mortality profiles and evolution of the predator-prey relationships in an environmental context. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 422, 101–127. [Google Scholar] [CrossRef] [Green Version]
- Sathiamurthy, E.; Voris, H.K. Maps of Holocene sea level transgression and submerged lakes on the Sunda Shelf. Nat. Hist. J. Chulalongkorn Univ. 2006, 2, 1–43. [Google Scholar]
- Fontaine, H.; Hoang, T.T.; Jaunngam, S.; Kavinate, S.; Salyapongse, S.; Sutheethorn, V.; Vachard, D. Paleontology and Stratigraphy of the Northwest Thailand: Paleogeographical Implications; Department of Mineral Resources: Bangkok, Thailand, 2009. [Google Scholar]
- Grave, P. Beyond the Mandala: Buddhist Landscapes and Upland-Lowland Interaction in North-West Thailand AD 1200–1650. World Archaeol. 1995, 27, 243–265. [Google Scholar]
- Marwick, B. Human behavioural ecology and stone artefacts in northwest Thailand during the terminal Pleistocene and Holocene. In Proceedings of the Southeast Asian Prehistory, the 11th EurASEAA Conference, Musée de Bougon, Paris, 25–30 September 2006; pp. 37–49. [Google Scholar]
- Shoocongdej, R.; Venunan, P.; Chintakanon, K.; Kutanan, W.; Wonnapinij, P.; Surat, W.; Rodcharoen, P.; Wattanapituksakul, W.; Na Nongkai, W. Final Report of Prehistoric Population and Cultural Dynamics in Highland Pang Mapha, Mae Hong Son Province, 2021; Volume 2. Archaeology, Submitted to Thailand Science Research and Innovation (TSRI) (in Thai)
- Siraboot, D.; Pureepatpong, N. Result of animal bone analysis. In Final Report of Highland Archaeology in Pang Mapha District, Mae Hong Son Province, 2003; Volume 2, 238–248. Submitted to Thailand Research Fund (TRF) (in Thai)
- Amphansri, A. Animal bone. Final Report of Highland Archaeology in Pang Mapha District, Mae Hong Son Province Phase 2, 2007; Volume 2. Submitted to Thailand Research Fund (TRF) (in Thai) [Google Scholar]
- Coolidge, H.J., Jr. The Indo-Chinese Forest Ox or Kouprey. Mem. Mus. Comp. Zoölogy Harv. Coll. 1940, 54, 421–531. [Google Scholar]
- Hooijer, D.A. Fossil Bovidae from the Malay Archipelago and the Punjab; Rijksmuseum van Natuurlijke: Leiden, Netherlands, 1958; Volume 38. [Google Scholar]
- Guérin, C. Gallogoral (nov. gen.) meneghini (Rütimeyer, 1878): Un Rupicaprine du Villafranchien d’Europe Occidentale. Ph.D. Thesis, University de Lyon, Lylon, France, 1966. [Google Scholar]
- Thein, T. La Fauna Néolithique du Phnom Loang (Cambodge). Ph.D. Thesis, University Paris VI, Paris, France, 1974. [Google Scholar]
- De Vos, J.; Long, V.T. Systematic Discussion of the Lang Trang Fauna, Vietnam. 1993; Unpublished Report. [Google Scholar]
- Bacon, A.M.; Demeter, F.; Duringer, P.; Helm, C.; Bano, M.; Long, V.T.; Thuy, N.T.K.; Antoine, P.O.; Mai, B.T.; Huong, N.T.M.; et al. The Late Pleistocene Duoi U’Oi cave in northern Vietnam: Palaeontology, sedimentology, taphonomy and palaeoenvironments. Quat. Sci. Rev. 2008, 27, 1627–1654. [Google Scholar] [CrossRef]
- Bacon, A.M.; Duringer, P.; Antoine, P.-O.; Demeter, F.; Shackelford, L.; Sayavongkhamdy, T.; Sichanthongtip, P.; Khamdalavong, P.; Nokhamaomphu, S.; Sysuphanh, V.; et al. The Middle Pleistocene mammalian fauna from Tam Hang karstic deposit, northern Laos: New data and evolutionary hypothesis. Quat. Int. 2011, 245, 315–332. [Google Scholar] [CrossRef]
- Filoux, A.; Wattanapituksakul, A.; Lespes, C.; Thongcharoenchaikit, C. A Pleistocene mammal assemblage containing Ailuropoda and Pongo from Tham Prakai Phet cave, Chaiyaphum province, Thailand. Geobios 2015, 48, 341–349. [Google Scholar] [CrossRef]
- Lyman, R.L. Quantitative Paleozoology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Reitz, E.J.; Wing, E.S. Zooarchaeology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Cannon, M.D. A Mathematical Model of the Effects of Screen Size on Zooarchaeological Relative Abundance Measures. J. Archaeol. Sci. 1999, 26, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Cannon, M.D. Archaeofaunal Relative Abundance, Sample Size, and Statistical Methods. J. Archaeol. Sci. 2001, 28, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Lyman, R.L. On the variable relationship between NISP and NTAXA in bird remains and in mammal remains. J. Archaeol. Sci. 2015, 53, 291–296. [Google Scholar] [CrossRef]
- O’Hara, R.; Kotze, J. Do not log-transform count data. Nat. Preced. 2010. [Google Scholar] [CrossRef]
- Conrad, C.; Wattanapituksakul, A.; Shoocongdej, R. Compendium for the paper Preservation of Mammalian Teeth and Bones Influences Terminal Pleistocene to Middle Holocene Hunter-Gatherer Subsistence at Ban Rai Rockshelter, Northwest Thailand. 2022. Available online: https://github.com/cylerc/BanRai (accessed on 1 November 2022).
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Barton, H.; Piper, P.J.; Rabett, R.; Reeds, I. Composite hunting technologies from the Terminal Pleistocene and Early Holocene, Niah cave, Borneo. J. Archaeol. Sci. 2009, 36, 1708–1714. [Google Scholar] [CrossRef]
- Piper, P.J. Human cultural, technological and adaptive changes from the end of the Pleistocene to the mid-Holocene in Southeast Asia. In The Routledge Handbook of Bioarchaeology in Southeast Asia and the Pacific Islands; Routledge: London, UK, 2015; pp. 52–72. [Google Scholar]
- Shoocongdej, R. Forager Mobility Organization in Seasonal Tropical Environments: A View from Lang Kamnan Cave, Western Thailand. Ph.D. Dissertation, Department of Anthropology, University of Michigan, Ann Arbor, MI, USA, 1996. [Google Scholar]
- Shoocongdej, R. Forager mobility organization in seasonal tropical environments of western Thailand. World Archaeol. 2000, 32, 14–40. [Google Scholar] [CrossRef]
- Amano, N.; Moigne, A.-M.; Ingicco, T.; Sémah, F.; Awe, R.D.; Simanjuntak, T. Subsistence strategies and environment in Late Pleistocene–Early Holocene Eastern Java: Evidence from Braholo Cave. Quat. Int. 2016, 416, 46–63. [Google Scholar] [CrossRef]
- Ingicco, T.; Amano, N.; Setiagama, K.; Moigne, A.M.; Budiman Sémah, A.M.; Simanjuntak, T.; Sémah, F. From Food to Grave Good: Nonhuman Primate Exploitation in Early to Mid-Holocene Eastern Java (Indonesia). Curr. Anthropol. 2020, 61, 264–277. [Google Scholar] [CrossRef]
- Marwick, B.; Gagan, M.K. Late Pleistocene monsoon variability in northwest Thailand: An oxygen isotope sequence from the bivalve Margaritanopsis laosensis excavated in Mae Hong Son province. Quat. Sci. Rev. 2011, 30, 3088–3098. [Google Scholar] [CrossRef]
- White, J.C.; Penny, D.; Kealhofer, L.; Maloney, B. Vegetation changes from the late Pleistocene through the Holocene from three areas of archaeological significance in Thailand. Quat. Int. 2004, 113, 111–132. [Google Scholar] [CrossRef]
- Frère, S.; Auetrakulvit, P.; Zeitoun, V.; Sophady, H.; Forestier, H.; Tan, N.H. Doi Pha Kan (Thailand), Ban Tha Si (Thailand) and Laang Spean (Cambodia) Animal Bone Assemblages: A New Perception of Meat Supply Strategies for Early olocene Mainland Southeast Asia. In Proceedings of the Advancing Southeast Asian Archaeology 2019: Selected Papers from the Second SEAMEO SPAFA International Conference on Southeast Asian Archaeology, Noel Hidalgo Tan, Bangkok, Thailand, May 2016; pp. 100–108. [Google Scholar]
- Zeitoun, V.; Auetrakulvit, P.; Forestier, H.; Zazzo, A.; Davtian, G.; Nakbunlung, S.; Tiamtinkrit, C. Discovery of a Mesolithic burial near the painted rock-shelter of Ban Tha Si (Lampang province, Northern Thailand): Implications for regional mortuary practices. CR Palevol 2013, 12, 127–136. [Google Scholar] [CrossRef]
- Zeitoun, V.; Auetrakulvit, P.; Zazzo, A.; Pierret, A.; Frère, S.; Forestier, H. Discovery of an outstanding Hoabinhian site from the Late Pleistocene at Doi Pha Kan (Lampang province, northern Thailand). Archaeol. Res. Asia 2019, 18, 1–16. [Google Scholar] [CrossRef]
- Auetrakulvit, P. Faunes du Pléistocène final à l’Holocène de Thaïlande: Approche archéozoologique. Ph.D. Dissertation, Université Aix-Marseille I, Marseille, France, 2004. [Google Scholar]
- Morwood, M.J.; Sutikna, T.; Saptomo, E.W.; Westaway, K.E.; Jatmiko Due, R.A.; Moore, M.W.; Yuniawati, D.Y.; Hadi, P.; Zhao, J.; Turney, C.S.M.; et al. Climate, people and faunal succession on Java, Indonesia: Evidence from Song Gupuh. J. Archaeol. Sci. 2008, 35, 1776–1789. [Google Scholar] [CrossRef]
- Higham, C. Early Mainland Southeast Asia from first Humans to Angor; River Books Co., Ltd.: Bangkok, Thailand, 2014. [Google Scholar]
- Anderson, D.D. Lang Rongrien rockshelter: A Pleistocene-Early Holocene Archaeological Site from Krabi, Southwestern Thailand; The University Museum: Philadelphia, PA, USA, 1990. [Google Scholar]
- Mudar, K.; Anderson, D. New evidence for Southeast Asian Pleistocene foraging economies: Faunal remains from the early levels of Lang Rongrien Rockshelter, Krabi, Thailand. Asian Perspect. 2007, 46, 298–334. [Google Scholar] [CrossRef] [Green Version]
- Wohlfarth, B.; Klubseang, W.; Inthongkaew, S.; Fritz, S.C.; Blaauw, M.; Reimer, P.J.; Chabangborn, A.; Löwemark, L.; Chawchai, S. Holocene environmental changes in northeast Thailand as reconstructed from a tropical wetland. Glob. Planet. Change 2012, 92, 148–161. [Google Scholar] [CrossRef]
- Chawchai, S.; Chabangborn, A.; Kylander, M.; Löwemark, L.; Mörth, C.-M.; Blaauw, M.; Klubseang, W.; Reimer, P.J.; Fritz, S.C.; Wohlfarth, B. Lake Kumphawapi-An archive of Holocene paleoenvironmental and paleoclimatic changes in northeast Thailand. Quat. Sci. Rev. 2013, 68, 59–75. [Google Scholar] [CrossRef]
- Lekagul, B.; McNeely, J.A. Mammals of Thailand; Kurusapa Ladprao Press: Bangkok, Thailand, 1988. [Google Scholar]
- Francis, C.M. A Guide to the Mammals of Southeast Asia; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Laboratory Code | Area | Layer | Material | δ13C | Radiocarbon Years BP | ±SD | Calibrated Year BP | Context |
|---|---|---|---|---|---|---|---|---|
| Beta-168215 | 1 | 2 | Charcoal | −25.0 | 9400 | 120 | 11,110–10,250 | Hunter-Gatherer |
| Beta-168216 | 1 | 4 | Charcoal | −28.8 | 10,600 | 40 | 12,910–12,610, 12,490–12,350 | Hunter-Gatherer |
| Beta-168218 | 2 | 4 | Charcoal | −25.0 | 9410 | 80 | 11,060–10,950, 10,780–10,420 | Hunter-Gatherer |
| Beta-168217 | 2 | 4 | Charcoal | −30.8 | 9720 | 50 | 11,210–11,090, 10,920–10,890 | Hunter-Gatherer |
| Beta-168219 | 2 | 4 | Charcoal | −28.4 | 10,210 | 50 | 12,340–11,670 | Hunter-Gatherer |
| Beta-534121 | 3 | - | Wood | −24.6 | 1770 | 30 | 1744–1605 | Log Coffin |
| Beta-534122 | 3 | - | Wood | −24.4 | 1840 | 30 | 1864–1708 | Log Coffin |
| Beta-534118 | 3 | - | Wood | −24.3 | 1880 | 30 | 1884–1728 | Log Coffin |
| Beta-534119 | 3 | - | Wood | −25.9 | 1920 | 30 | 1947–1812 | Log Coffin |
| Beta-534120 | 3 | - | Wood | −25.5 | 2040 | 30 | 2067–1924 | Log Coffin |
| Beta-538749 | 3 | - | Sediment | - | 4550 | 30 | 5189–5053, 5318–5257 | Hunter-Gatherer |
| Beta-168220 | 3 | 3 | Charcoal | −28.0 | 7250 | 40 | 8340–8140 | Hunter-Gatherer |
| Beta-168222 | 3 | 5 | Charcoal | −29.7 | 8190 | 50 | 9290–9120 | Hunter-Gatherer |
| Beta-168221 | 3 | 5 | Charcoal | −27.7 | 8850 | 50 | 10,170–9720 | Hunter-Gatherer |
| Taxon | Common Name | Area 1 S1E4 | Area 1 S2E12 | Area 2 S1E6 | Area 2 S2E14 | Area 2 Baulk S2E14 | Area 3 N5E39 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | ||
| Mammalia | Mammal | 51 | 4 | 29 | 6 | 21 | 4 | 17 | 3 | 51 | 4 | 88 | 5 |
| Manis sp. | Pangolin | 1 | 1 | ||||||||||
| Nycticebus sp. | Loris | 1 | 1 | ||||||||||
| Macaca mulatta | Rhesus macaque | 13 | 6 | ||||||||||
| Macaca arctoides | Stump-tailed macaque | 11 | 7 | ||||||||||
| Macaca sp. | Macaque | 1 | 1 | 2 | 1 | 5 | 1 | 2 | 1 | 243 | 19 | ||
| Cercopithecidae | Old World monkeys | 4 | 2 | 1 | 1 | 8 | 3 | 1 | 1 | ||||
| Trachypithecus cf. phayrei | Phayre’s langur | 2 | 2 | ||||||||||
| Colobinae | Langur | 1 | 1 | 2 | 1 | 1 | 1 | 73 | 9 | ||||
| Hylobatidae | Gibbon | 10 | 2 | ||||||||||
| Ursus thibetanus | Asia black bear | 1 | 1 | ||||||||||
| Ursus spp. | Bear | 1 | 1 | 4 | 2 | ||||||||
| Arctonyx collaris | Hog badger | 1 | 1 | ||||||||||
| Arctictis binturong | Binturong | 1 | 1 | ||||||||||
| Canis sp.? | Dog | 8 | 2 | ||||||||||
| Panthera sp. | Tiger or leopard | 2 | 1 | ||||||||||
| Felidae | Felids | 6 | 3 | ||||||||||
| Carnivora | Carnivors | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 2 | 26 | 5 | ||
| Elephas sp. | Elephant | 4 | 1 | ||||||||||
| Rhinocerotidae | Rhinoceros | 10 | 2 | ||||||||||
| Sus scrofa | Eurasian wild pig | 1 | 1 | 3 | 3 | 8 | 4 | ||||||
| Muntiacus sp. | Muntjac | 8 | 3 | 2 | 2 | 13 | 4 | ||||||
| Muntiacus feae? | Fea’s muntjac | 4 | 2 | ||||||||||
| Panolia eldii | Eld’s deer | 10 | 3 | 2 | 2 | 3 | 1 | ||||||
| Rusa unicolor | Sambar deer | 30 | 7 | 20 | 5 | 30 | 4 | 22 | 4 | 33 | 7 | ||
| Panolia/Rusa sp. | Sambar/Eld’s deer | 18 | 4 | ||||||||||
| Axis sp. | Hog deer | 9 | 5 | 4 | 2 | 8 | 3 | 12 | 1 | 19 | 6 | ||
| Cervidae | Cervids | 3 | 2 | 1 | 1 | 19 | 4 | ||||||
| Bubalus arnee | Wild water buffalo | 1 | 1 | 1 | 1 | 5 | 3 | ||||||
| Bos gaurus | Gaur | 1 | 1 | 2 | 2 | ||||||||
| Bos javanicus | Banteng | 1 | 1 | ||||||||||
| Bos sp. | Wild cattle | 2 | 2 | 4 | 2 | 6 | 3 | ||||||
| Bovinae | Bovid | 1 | 1 | 1 | 1 | ||||||||
| Capricornis sumatraensis | Sumatran serow | 2 | 2 | 5 | 3 | 1 | 1 | 8 | 2 | ||||
| Naemorhedus griseus | Chinese goral | 1 | 1 | ||||||||||
| Artiodactyla | Even-toed ungulates | 1 | 1 | ||||||||||
| Callosciurus finlaysonii | Variable squirrel | 1 | 1 | ||||||||||
| Petauristinae | Flying squirrel | 1 | 1 | ||||||||||
| Sciuridae | Squirrel | 5 | 2 | ||||||||||
| Leopoldamys sabanus | Long-tailed giant rat | 1 | 1 | ||||||||||
| Bandicota sp. | Bandicoot rat | 1 | 1 | ||||||||||
| Rhizomyidae | Bamboo rat | 3 | 1 | 1 | 1 | 2 | 2 | 6 | 2 | ||||
| Hystricidae | Old World porcupine | 19 | 3 | ||||||||||
| Rodentia | Rodent | 29 | 6 | ||||||||||
| Aves | Bird | 1 | 1 | ||||||||||
| Lacertilia | Lizards | 3 | 2 | ||||||||||
| Testudinata | Turtle | 22 | 5 | 30 | 6 | 2 | 2 | 12 | 3 | 15 | 4 | 53 | 4 |
| Indotestudo elongata | Elongated tortoise | 28 | 2 | ||||||||||
| Cyprinidae | Carp or minnow | 2 | 2 | 1 | 1 | 2 | 1 | 61 | 15 | ||||
| Eumalacostraca | Crab | 1 | 1 | ||||||||||
| Total | 155 | 43 | 96 | 29 | 119 | 33 | 90 | 24 | 74 | 15 | 818 | 150 | |
| Layer | Butchery Mark 1 | Bone Tool 1 | Burning | Weathering 1 | Chemical Process 1 | Gnawing Damage 1 | Root Etching 1 | Total |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 3 | 15 | 0 | 1 | 5 | 0 | 25 |
| 2 | 6 | 7 | 25 | 0 | 0 | 5 | 1 | 44 |
| 3 | 8 | 14 | 21 | 1 | 3 | 6 | 0 | 53 |
| 4 | 7 | 8 | 10 | 0 | 2 | 4 | 0 | 31 |
| 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 8 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Total | 22 | 32 | 72 | 1 | 6 | 20 | 1 | 154 |
| Layer | Butchery Mark 1 | Bone Tool 1 | Burning | Weathering 1 | Chemical Process 1 | Gnawing Damage 1 | Root Etching 1 | Total |
|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 12 | 11 | 0 | 0 | 6 | 0 | 32 |
| 2 | 0 | 9 | 21 | 0 | 0 | 4 | 1 | 35 |
| 3 | 0 | 4 | 13 | 0 | 1 | 3 | 0 | 21 |
| 4 | 5 | 27 | 37 | 0 | 0 | 7 | 0 | 76 |
| 5 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 3 |
| Total | 8 | 52 | 85 | 0 | 1 | 20 | 1 | 167 |
| Layer | Butchery Mark 1 | Bone Tool 1 | Burning | Weathering 1 | Chemical Process 1 | Gnawing Damage 1 | Root Etching 1 | Total |
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 2 | 14 | 0 | 1 | 4 | 0 | 23 |
| 2 | 1 | 2 | 21 | 0 | 1 | 5 | 0 | 30 |
| 3,4 | 7 | 12 | 72 | 0 | 4 | 11 | 0 | 106 |
| 5 | 12 | 24 | 38 | 0 | 0 | 3 | 0 | 77 |
| 6 | 1 | 2 | 13 | 0 | 1 | 1 | 0 | 18 |
| Total | 23 | 42 | 158 | 0 | 7 | 24 | 0 | 254 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wattanapituksakul, A.; Shoocongdej, R.; Conrad, C. Preservation of Mammalian Teeth and Bones Influences Identification of Terminal Pleistocene to Middle Holocene Hunter-Gatherer Subsistence at Ban Rai Rockshelter, Northwest Thailand. Quaternary 2022, 5, 51. https://doi.org/10.3390/quat5040051
Wattanapituksakul A, Shoocongdej R, Conrad C. Preservation of Mammalian Teeth and Bones Influences Identification of Terminal Pleistocene to Middle Holocene Hunter-Gatherer Subsistence at Ban Rai Rockshelter, Northwest Thailand. Quaternary. 2022; 5(4):51. https://doi.org/10.3390/quat5040051
Chicago/Turabian StyleWattanapituksakul, Athiwat, Rasmi Shoocongdej, and Cyler Conrad. 2022. "Preservation of Mammalian Teeth and Bones Influences Identification of Terminal Pleistocene to Middle Holocene Hunter-Gatherer Subsistence at Ban Rai Rockshelter, Northwest Thailand" Quaternary 5, no. 4: 51. https://doi.org/10.3390/quat5040051