

Chihuahuan Desert Vegetation Development during the Past 10,000 Years According to Pollen and Sediment Data at Upper Arroyo, Saltillo, Mexico

Abstract
:1. Introduction
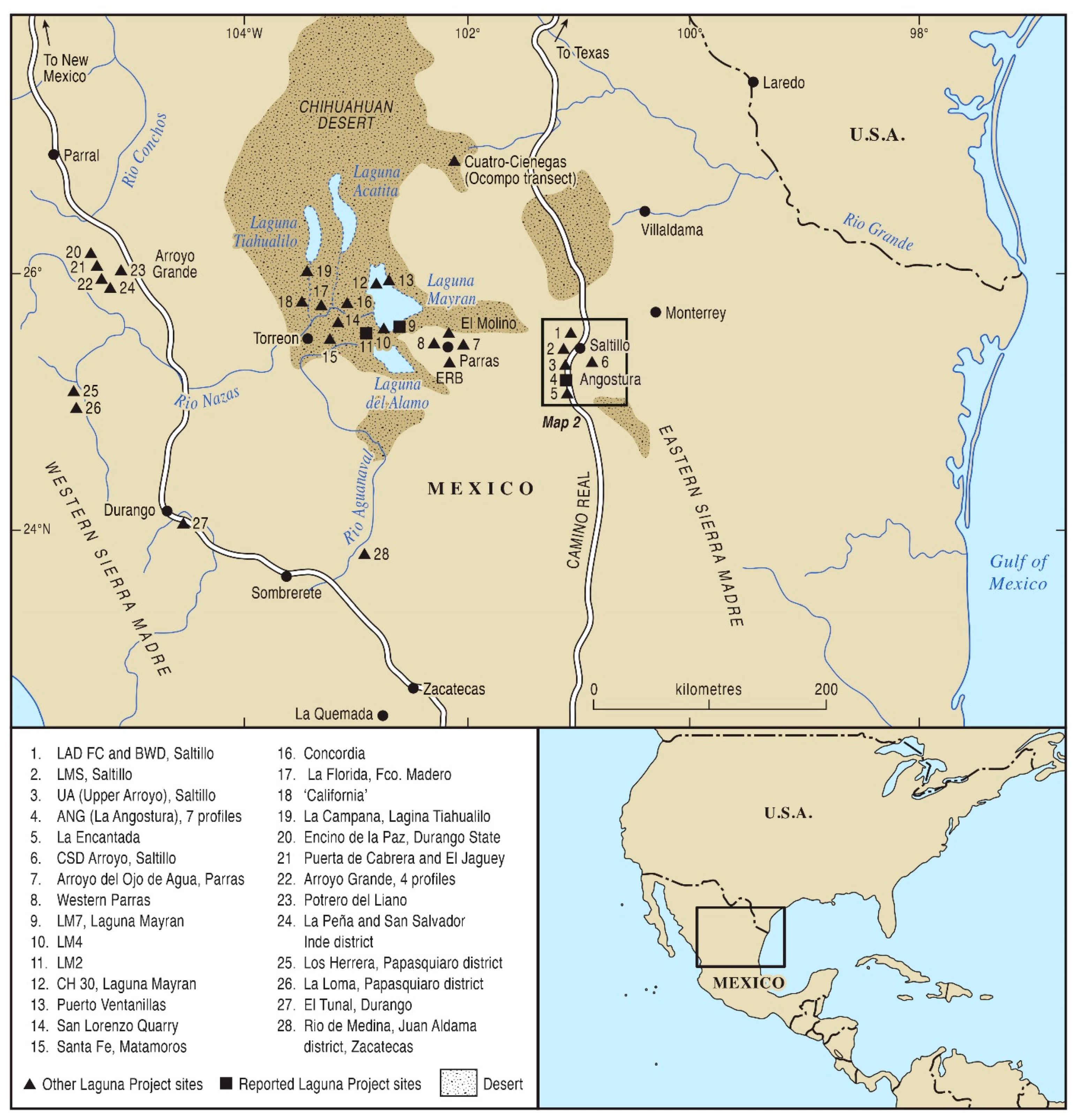
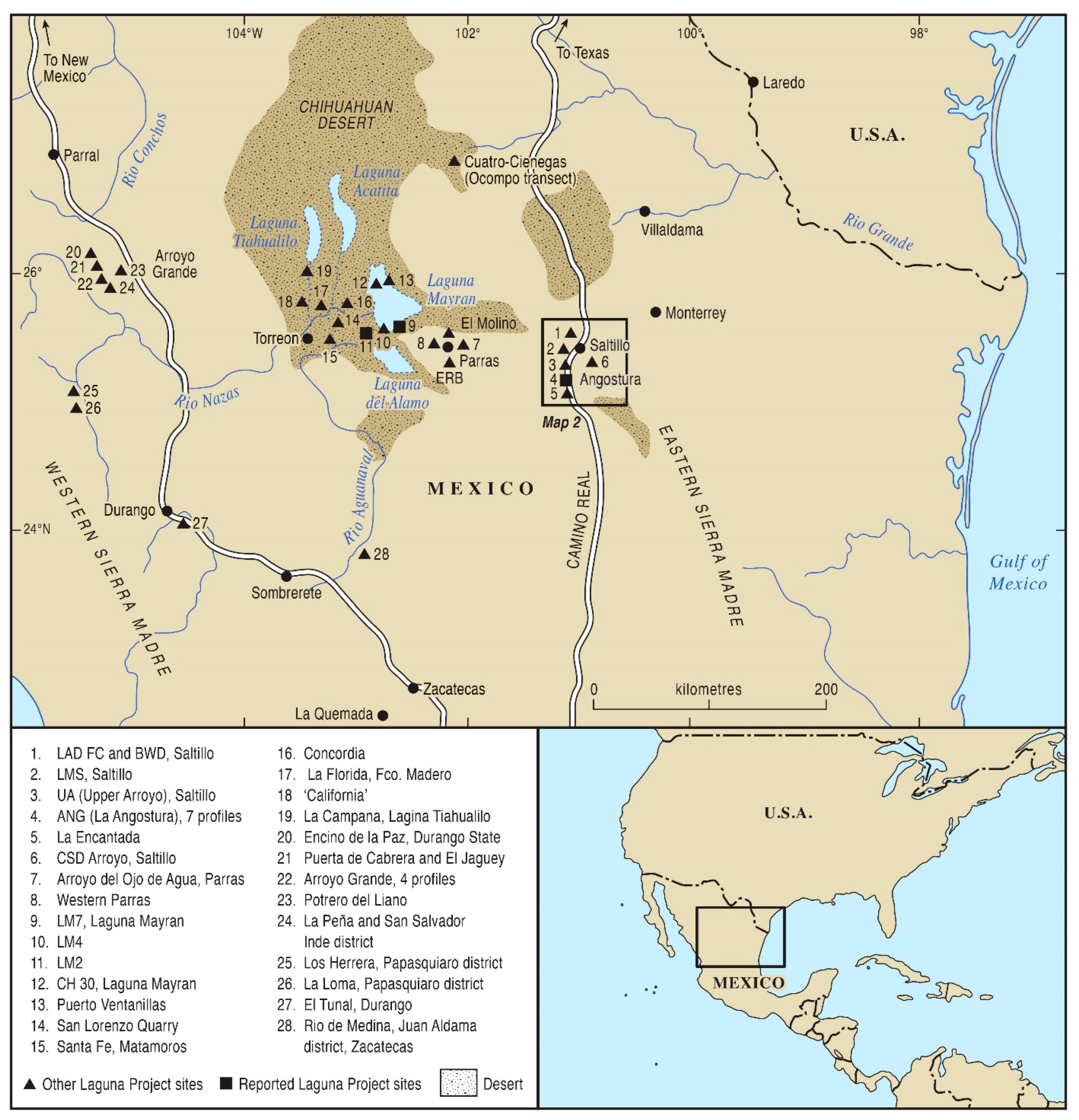
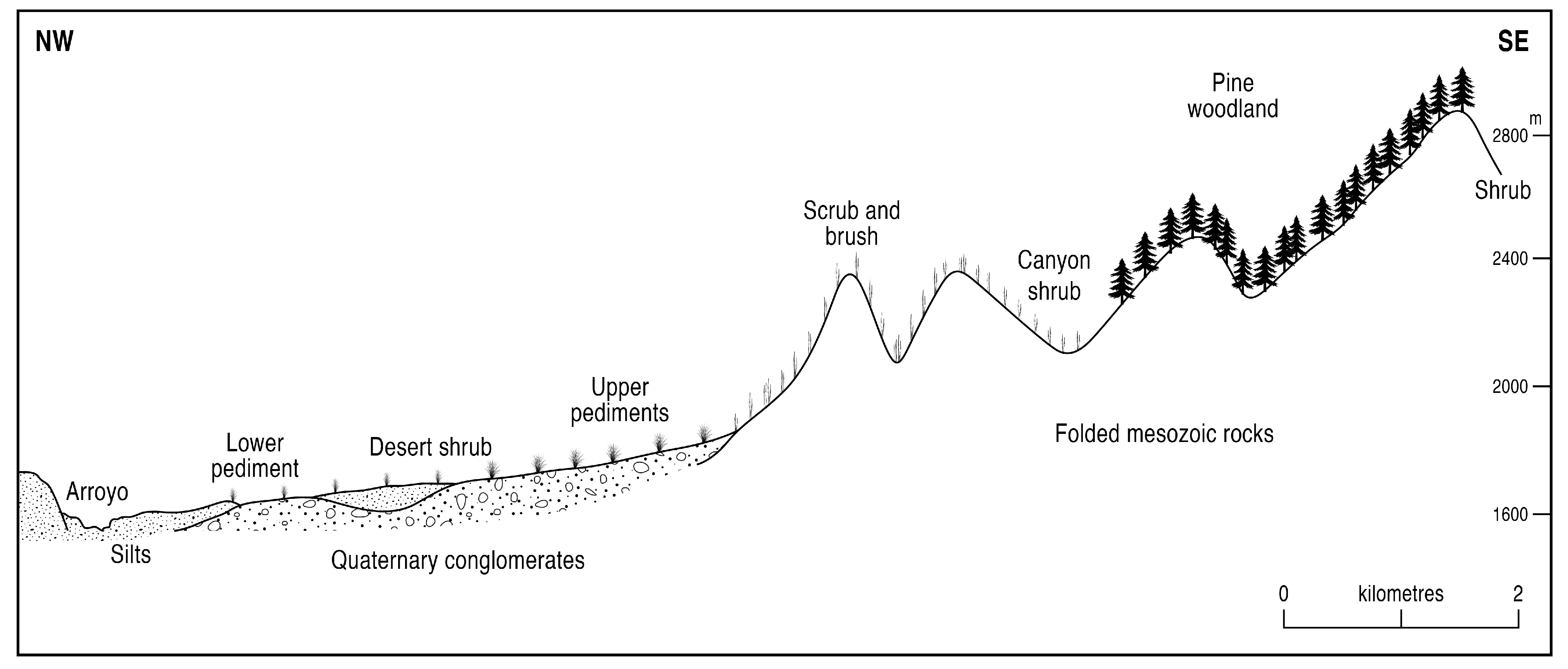
2. Study Region and Materials
3. Stratigraphic Pollen Studies in Northern Mexico
4. Field and Laboratory Methods
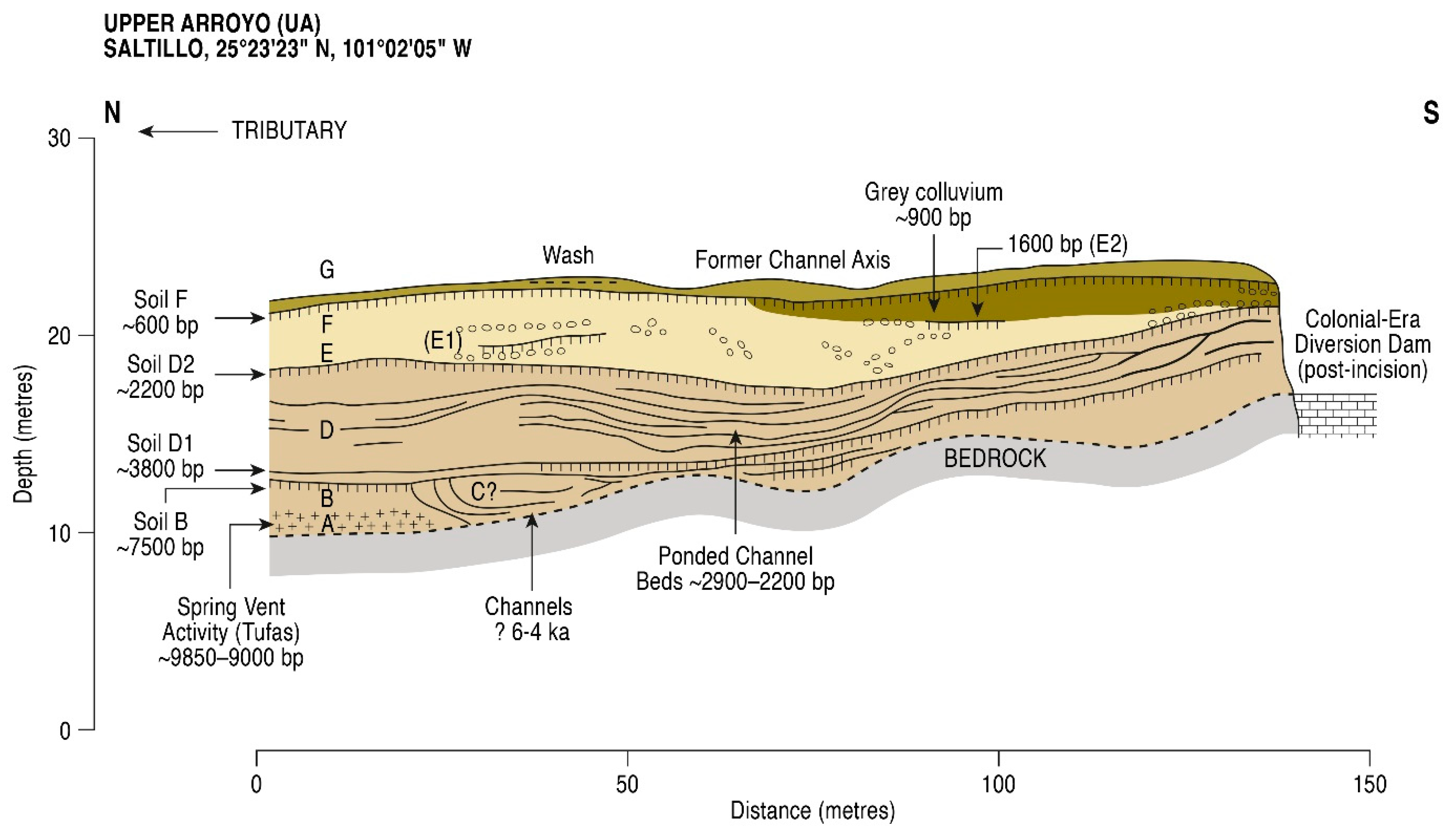
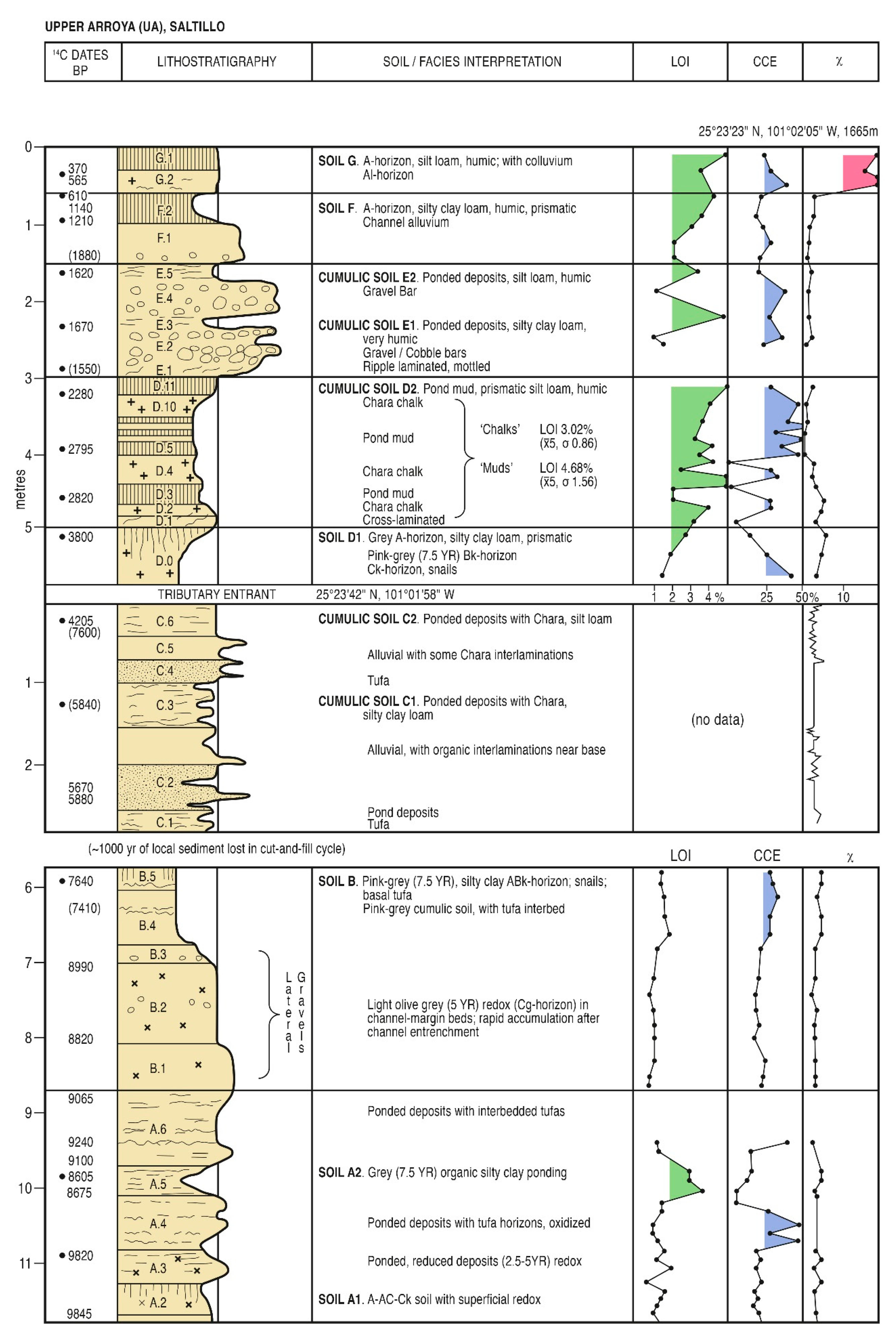
5. Results
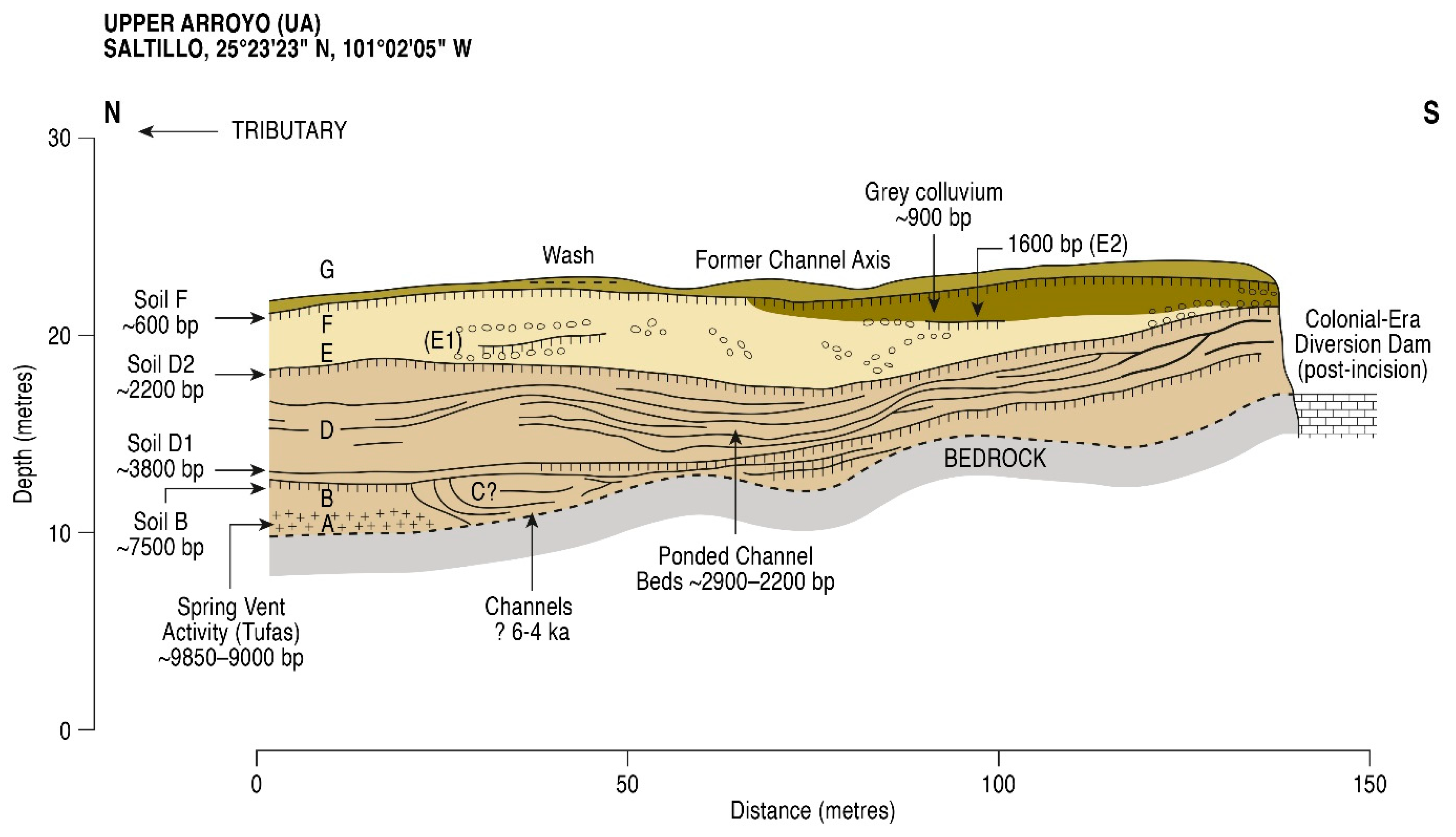
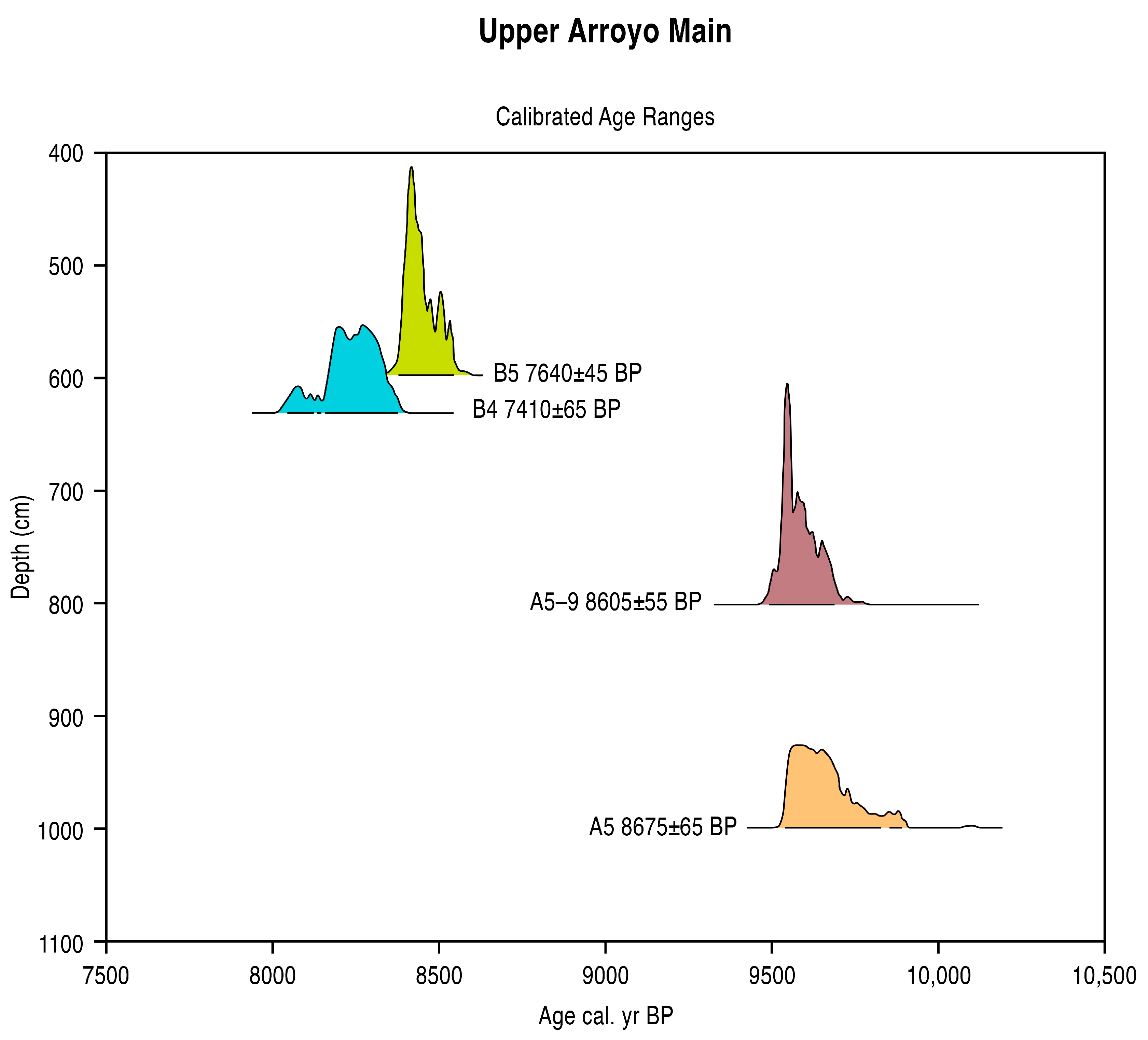
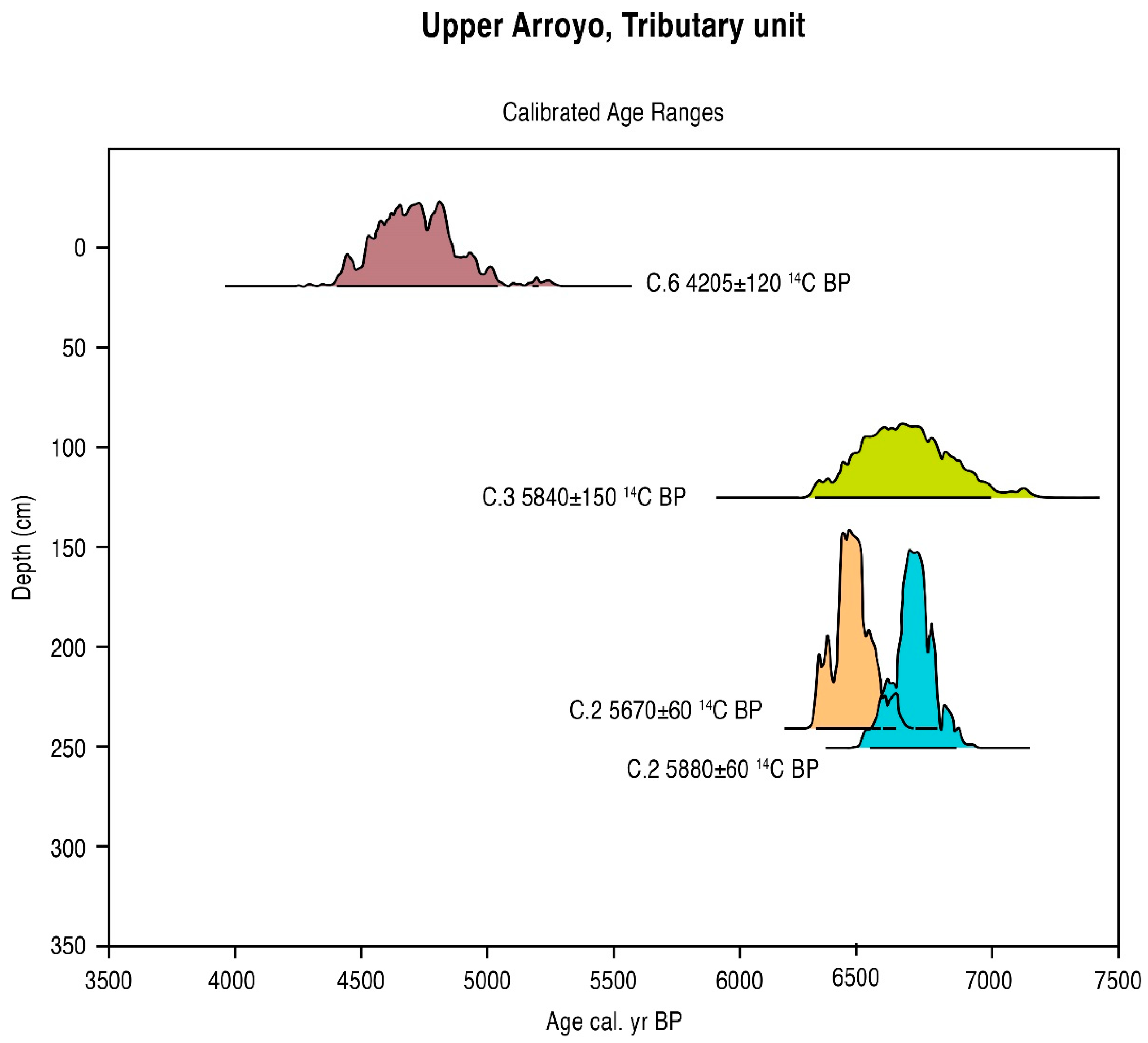
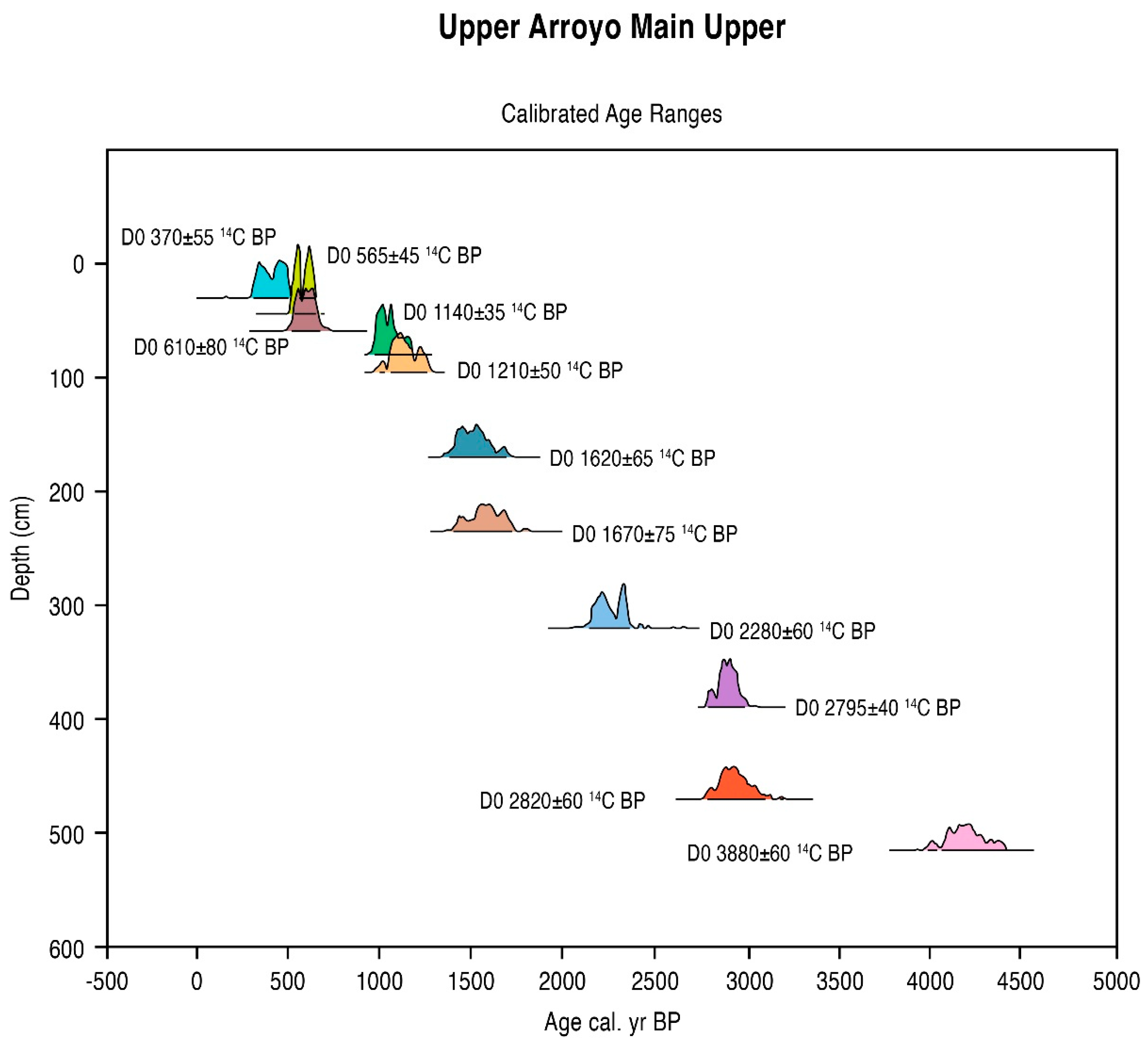
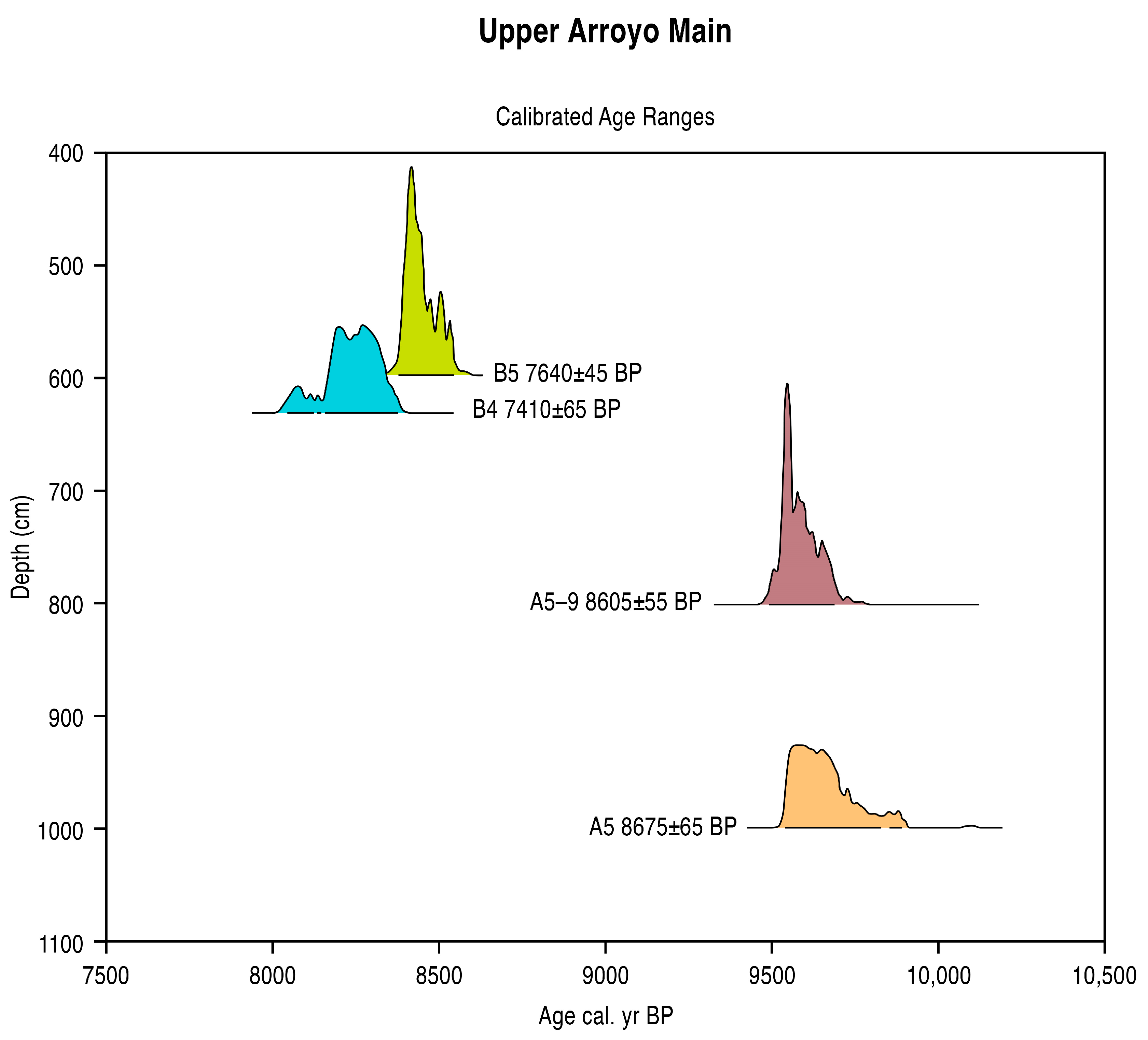
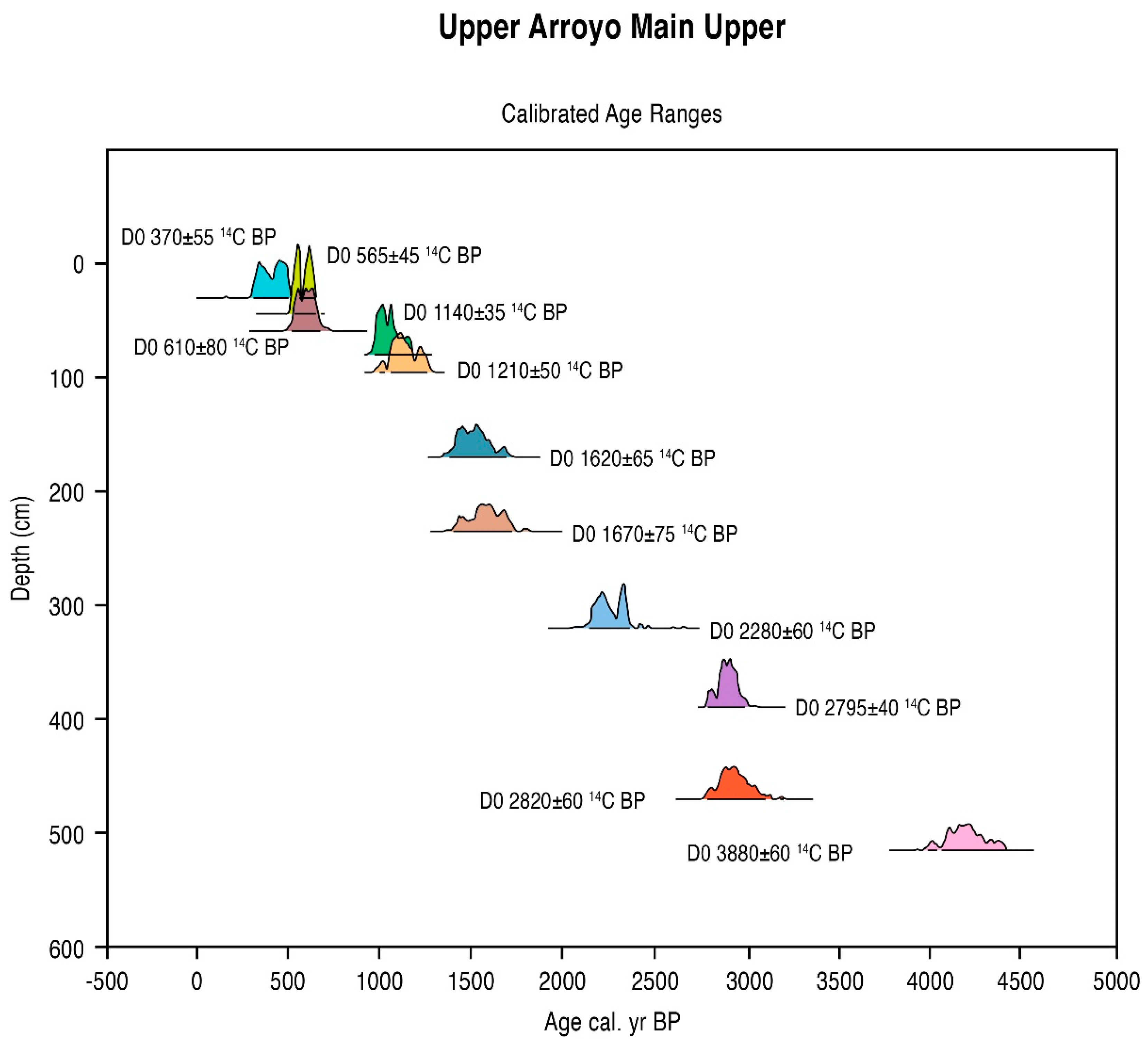
5.1. Radiometric and AMS Dating
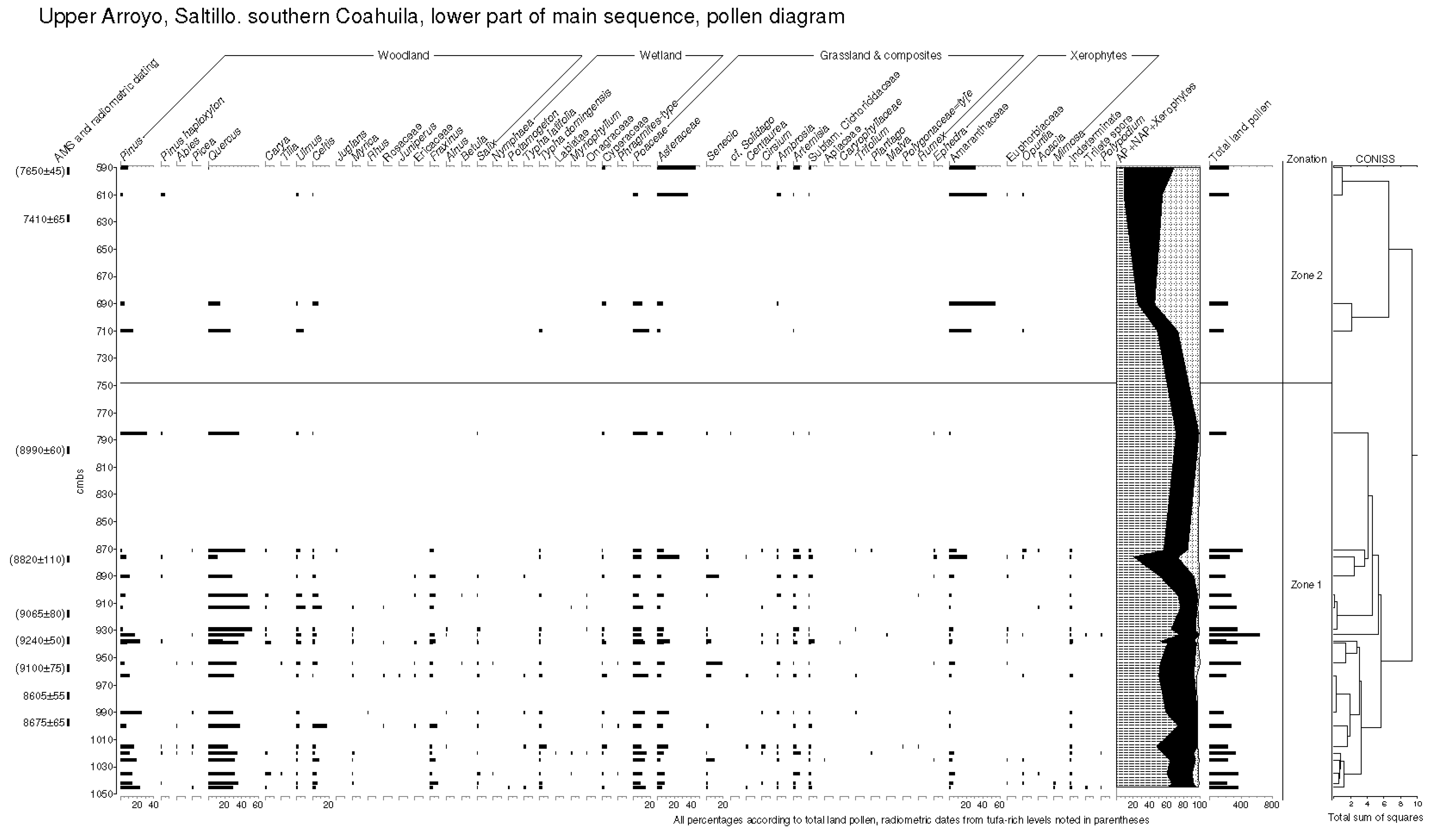
- The presence of spring vents in situ within a limestone bedrock geology (Figure 3), giving rise to a hard-water local hydrology. Note the presence of calcareous tufas;
- The presence of aquatic and semi-aquatic flora according to the pollen data of Zone 1 in the lower part of the main sequence (see Section 5.2.1 below) presented an ecology that would permit a hard-water error, as opposed to that of terrestrial plants;
- An apparent, non-random error factor of circa +1000 y calculated in this part of the sequence in the rejected date series (4). A systematic error resulting from hard-water (as opposed to a re-working of old, or intrusion of young sediments) would be expected to be expressed at a relatively constant differential;
- Pollen dating of the Terminal Pleistocene site of La Angostura at a 7 km distance from Upper Arroyo. An early dating of Upper Arroyo (i.e., an acceptance of the series of rejected dates) would place Zone 1 within the timeframe of the Artemisia steppe formation in the basin of Upper Arroyo. This vegetation unit is not apparent in the Zone 1 pollen data in the lower aspect of the Upper Arroyo sequence.
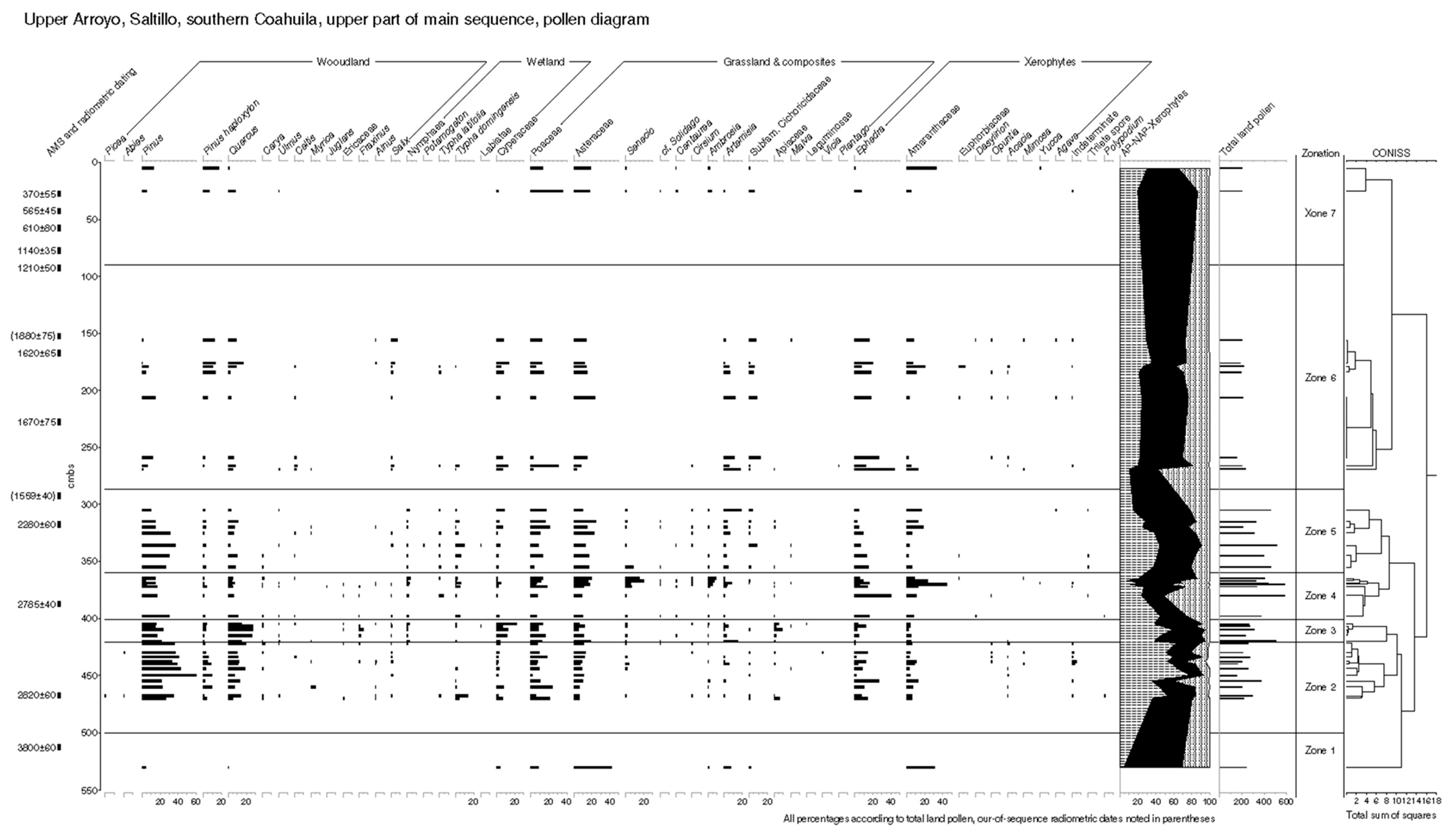
5.2. Palynological Analyses
5.2.1. Pollen Zonation and Sedimentary Interpretation of the Early Holocene Sequence
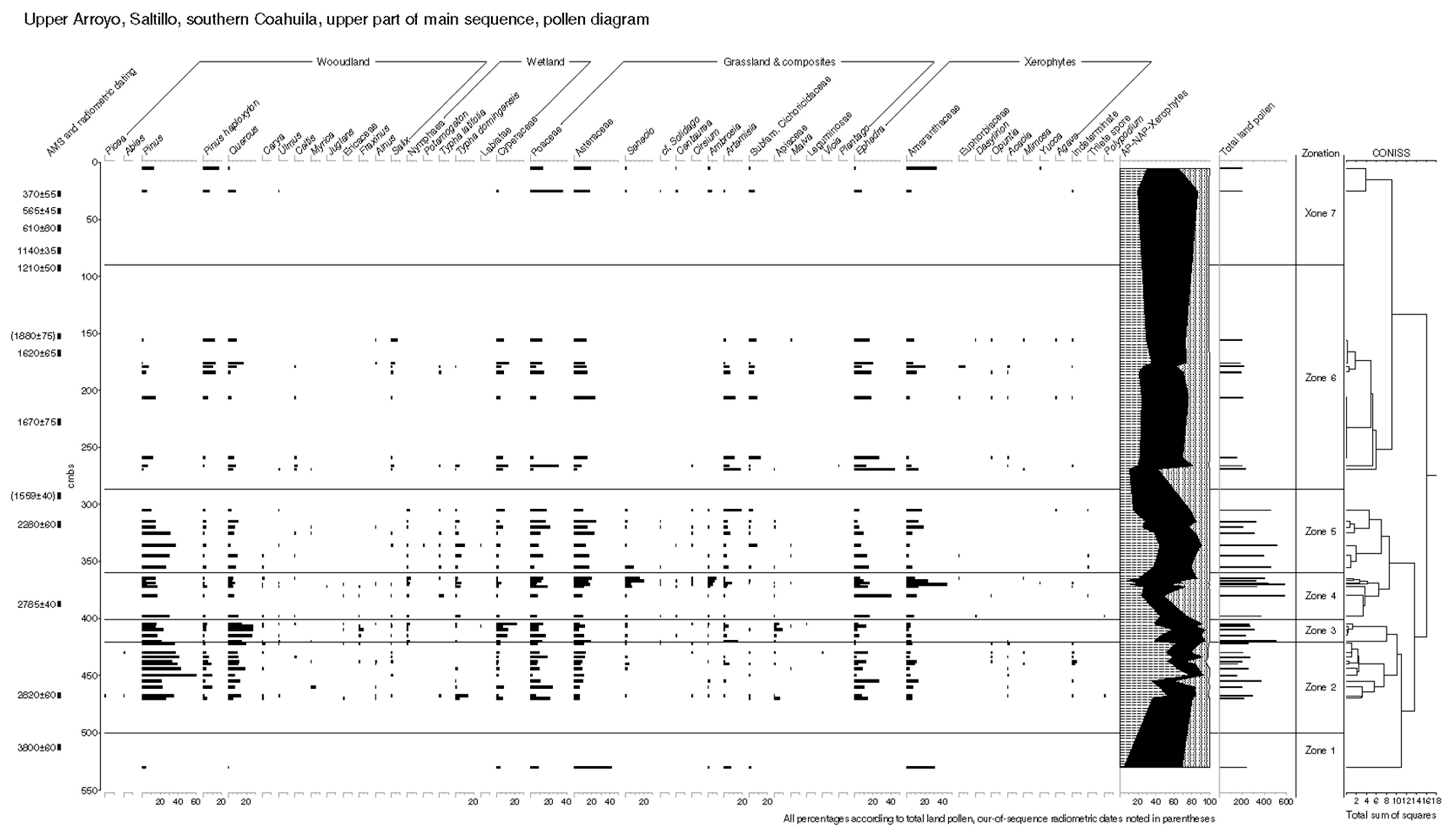
5.2.2. Pollen Zonation and Interpretation of the Mid-Holocene Sequence
5.2.3. Results: Zonation and Interpretation of Late-Holocene Sequence
6. Discussion
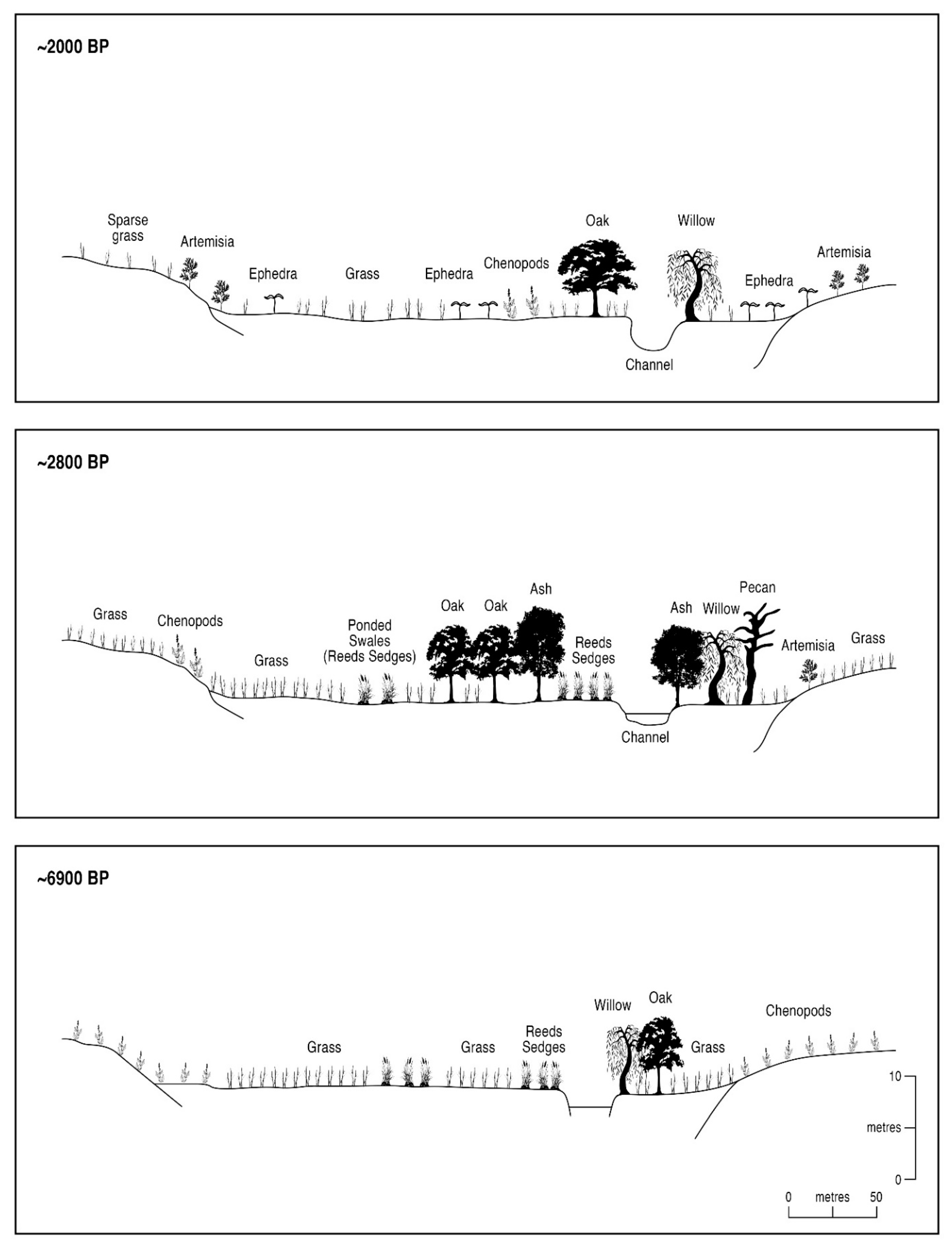
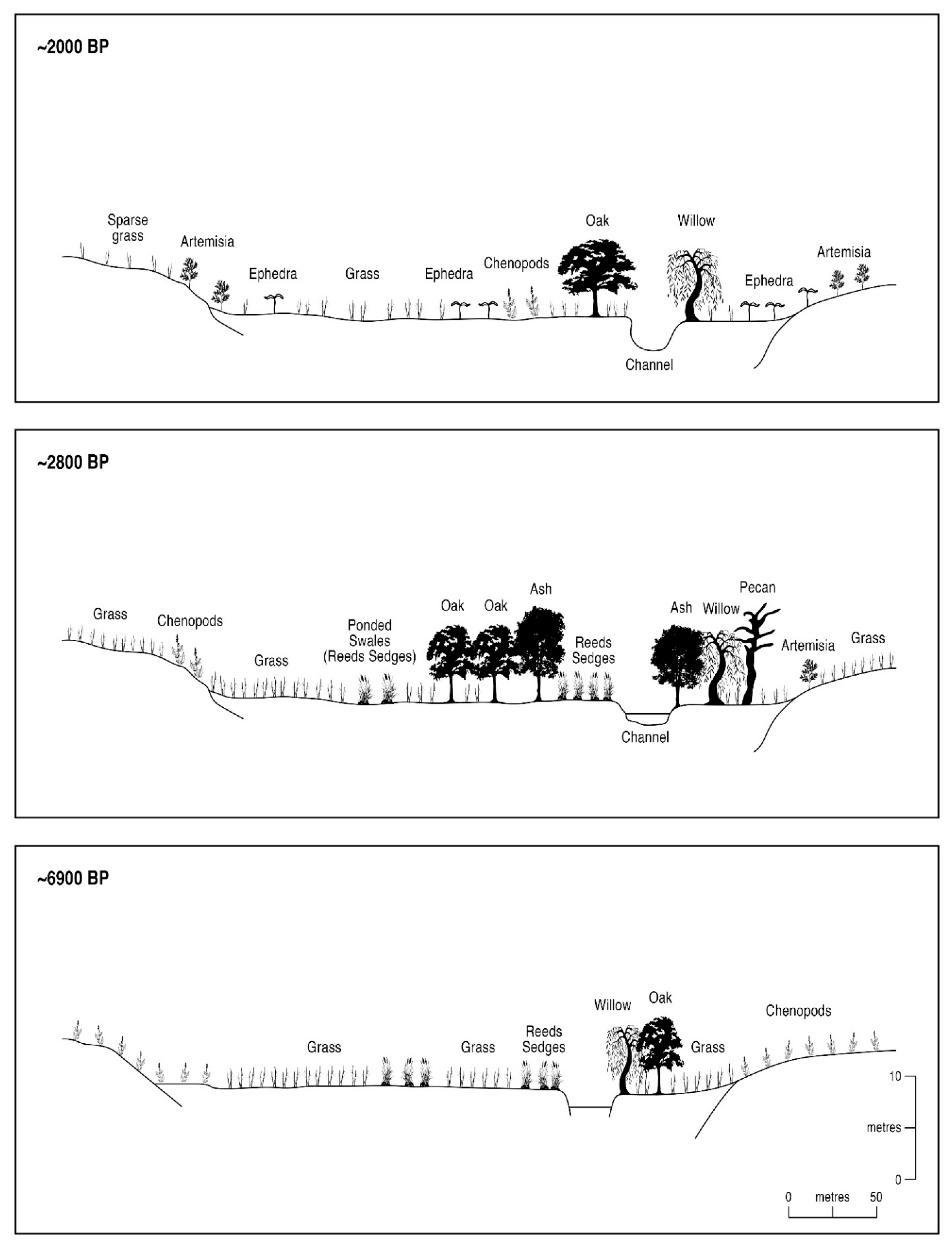
6.1. Zonal vs. Azonal Floral Changes at Upper Arroyo
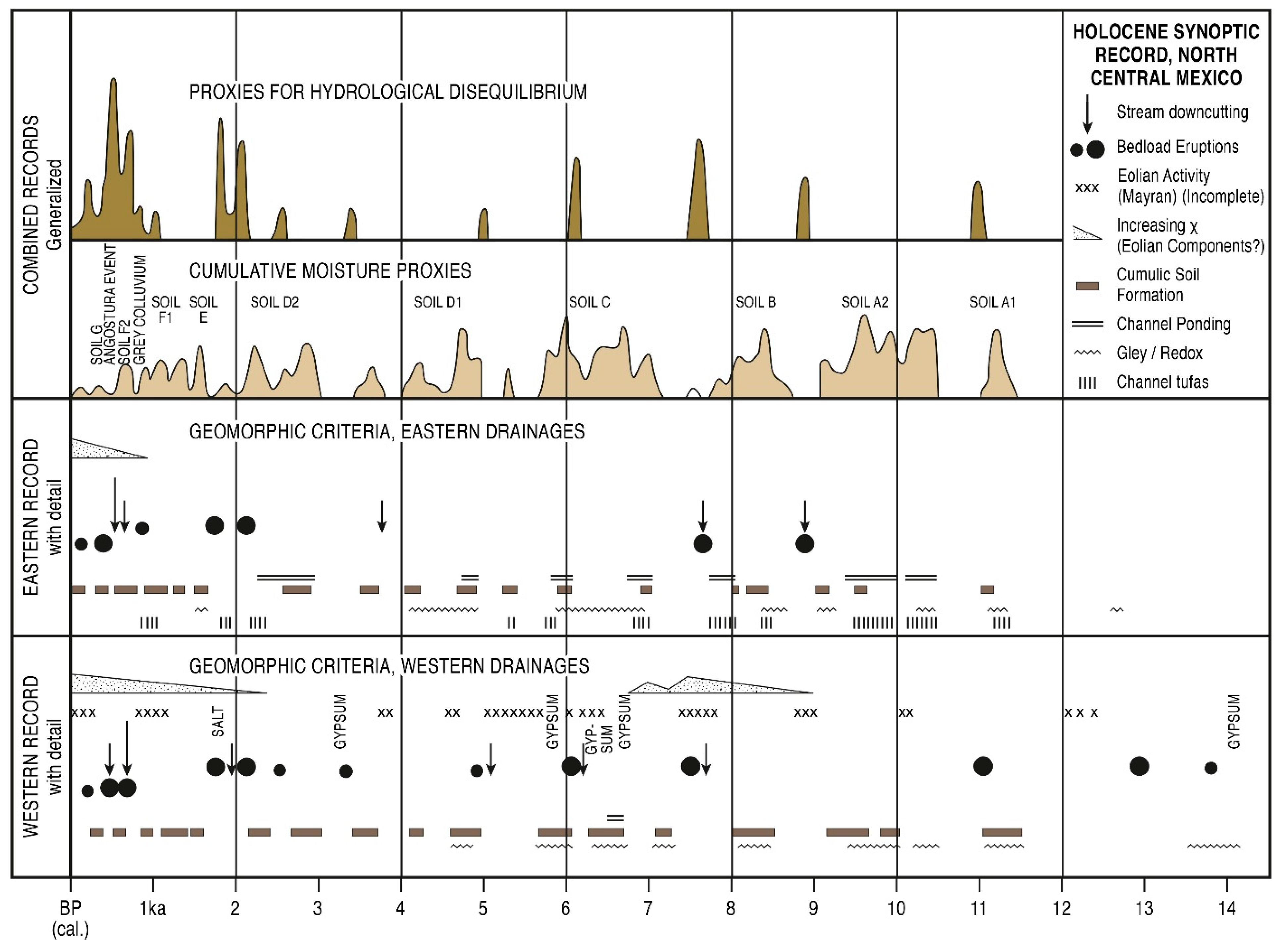
6.2. Comparative Environmental Records in North and Central Mexico
7. Conclusions
7.1. History of Desert Flora and Associated Sedimentation Processes
7.2. Causes of Desertification—Climatic and Physiographic Factors
7.3. Azonal Desertification Processes and Endemism of Upland Flora
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sosa, V.; De-Nova, J.A.; Vásquez-Cruz, M. Evolutionary history of the flora of Mexico: Dry forests cradles and museums of endemism. J. Syst. Evol. 2018, 56, 523–536. [Google Scholar] [CrossRef]
- de León, R.M.; Castellanos-Morales, G.; Moreno-Letelier, A. Incipient speciation, high genetic diversity, and ecological divergence in the alligator bark juniper suggest complex demographic changes during the Pleistocene. Peerj 2022, 10, e13802. [Google Scholar] [CrossRef] [PubMed]
- Buxbaum, C.A.Z.; Vanderbilt, K. Soil heterogeneity and the distribution of desert and steppe plant species across a desert-grassland ecotone. J. Arid. Environ. 2007, 69, 617–632. [Google Scholar] [CrossRef]
- Wainwright, J.; Parsons, A.J.; Abrahams, A.D. Plot-scale studies of vegetation, overland flow and erosion interactions: Case studies from Arizona and New Mexico. Hydrol. Process. 2000, 14, 2921–2943. [Google Scholar] [CrossRef]
- Li, J.; Okin, G.S.; Alvarez, L.; Epstein, H. Quantitative effects of vegetation cover on wind erosion and soil nutrient loss in a desert grassland of southern New Mexico, USA. Biogeochemistry 2007, 85, 317–332. [Google Scholar] [CrossRef]
- Castro-López, V.; Domínguez-Vázquez, G.; Islebe, G.A.; Priego-Santander, A.G.; Velázquez, A. Modern pollen-vegetation relationships across a landscape mosaic in central México. Rev. Palaeobot. Palynol. 2021, 289, 104362. [Google Scholar] [CrossRef]
- Brown, A.G.; Carpenter, R.G.; Walling, D.E. Monitoring fluvial pollen transport, its relationship to catchment vegetation and implications for palaeoenvironmental studies. Rev. Palaeobot. Palynol. 2007, 147, 60–76. [Google Scholar] [CrossRef]
- Liu, D.; Ma, J.; Wu, P.; Pan, Y. A new indicator for dividing sedimentary rhythms in alluvial deposits: A pollen-based method. Catena 2020, 189, 104500. [Google Scholar] [CrossRef]
- Albert, B.M. Holocene aridification, vegetation change, sedimentation regime and limits of carbon isotope data as indicated by the alluvial pollen sites of Arroyo Grande and El Molino in North-Central Mexico. Quat. Int. 2015, 377, 2–17. [Google Scholar] [CrossRef]
- Albert, B.M. Acidification and Pine Expansion in East Texas According to Pollen Evidence from Dual Cores in Alluvium. Castanea 2011, 76, 164–177. [Google Scholar] [CrossRef]
- Albert, B.; Innes, J.; Krementskiy, K.; Millard, A.R.; Gaydarska, B.; Nebbia, M.; Chapman, J. What was the ecological impact of a Trypillia megasite occupation? Multi-proxy palaeo-environmental investigations at Nebelivka, Ukraine. Veg. Hist. Archaeobot. 2019, 29, 15–34. [Google Scholar] [CrossRef]
- Albert, B.; Pokorný, P. Pollen Taphonomy and Hydrology at Vranský potok versus Zahájí Alluvial Pollen sites: Methodological Implications for Cultural Landscape Reconstruction in the Peruc Sandstone Area, Czech Republic. Interdiscip. Archaeol.—Nat. Sci. Archaeol. 2012, III, 85–101. [Google Scholar] [CrossRef]
- Butzer, K.W.; Abbott, J.T.; Frederick, C.D.; Lehman, P.H.; Cordova, C.E.; Oswald, J.F. Soil-geomorphology and “wet” cycles in the Holocene record of North-Central Mexico. Geomorphology 2008, 101, 237–277. [Google Scholar] [CrossRef]
- Muller, C.H. Plants as Indicators of Climate in Northeast Mexico. Am. Midl. Nat. 1937, 18, 986. [Google Scholar] [CrossRef]
- Holmes, P.L. Differential transport of spores and pollen: A laboratory study. Rev. Palaeobot. Palynol. 1990, 64, 289–296. [Google Scholar] [CrossRef]
- Jacobsen, G.L.; Bradshaw, R.H. The selection of sites for palaeoecological studies. Quat. Res. 1981, 16, 80–96. [Google Scholar] [CrossRef]
- Ortega-Rosas, C.I.; Peñalba, M.C.; Guiot, J. Holocene altitudinal shifts in vegetation belts and environmental changes in the Sierra Madre Occidental, Northwestern Mexico, based on modern and fossil pollen data. Rev. Palaeobot. Palynol. 2008, 151, 1–20. [Google Scholar] [CrossRef]
- Meyer, E.R. Late-Quarternary Paleoecology of the Cuatro Cienegas Basin, Coahuila, Mexico. Ecology 1973, 54, 982–995. [Google Scholar] [CrossRef]
- Felstead, N.J.; Gonzalez, S.; Huddart, D.; Noble, S.R.; Hoffmann, D.L.; Metcalfe, S.E.; Leng, M.J.; Albert, B.M.; Pike, A.W.; Gonzalez-Gonzalez, A.; et al. Holocene-aged human footprints from the Cuatrociénegas Basin, NE Mexico. J. Archaeol. Sci. 2014, 42, 250–259. [Google Scholar] [CrossRef]
- Ortega-Rosas, C.I.; Peñalba, M.C.; Guiot, J. The Lateglacial interstadial at the southeastern limit of the Sonoran Desert, Mexico: Vegetation and climate reconstruction based on pollen sequences from Ciénega San Marcial and comparison with the subrecent record. Boreas 2016, 45, 773–789. [Google Scholar] [CrossRef]
- Albert, B.M. Past vegetation influence on the hydrological cycle in the Chihuahuan Desert, North Mexico as indicated by data of four pollen sites. Quat. Int. 2015, 374, 168–181. [Google Scholar] [CrossRef]
- Albert, B.M. Prehistoric and Colonial land-use in the Chihuahuan Desert as inferred from pollen and sedimentary data from Coahuila, Mexico. Quat. Int. 2015, 377, 38–51. [Google Scholar] [CrossRef]
- Caballero-Rodríguez, D.; Correa-Metrio, A.; Lozano-García, S.; Sosa-Nájera, S.; Ortega, B.; Sanchez-Dzib, Y.; Aguirre-Navarro, K.; Sandoval-Montaño, A. Late-Quaternary spatiotemporal dynamics of vegetation in Central Mexico. Rev. Palaeobot. Palynol. 2018, 250, 44–52. [Google Scholar] [CrossRef]
- Ashworth, J.; Keyes, D.; Kirk, R.; Lessard, R. Standard Procedure in the Hydrometer Method For Particle Size Analysis. Commun. Soil Sci. Plant Anal. 2001, 32, 633–642. [Google Scholar] [CrossRef]
- Faegri, K.; Kaland, P.E.; Krzywinski, K. Textbook of Pollen Analysis, 4th ed.; John Wiley & Sons Ltd.: Chichester, UK, 1989. [Google Scholar]
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- England, J.; Dyke, A.S.; Coulthard, R.D.; Mcneely, R.; Aitken, A. The exaggerated radiocarbon age of deposit-feeding molluscs in calcareous environments. Boreas 2012, 42, 362–373. [Google Scholar] [CrossRef]
- Trachsel, M.; Telford, R.J. All age–Depth models are wrong, but are getting better. Holocene 2017, 27, 860–869. [Google Scholar] [CrossRef]
- Chávez-Lara, C.M.; Holtvoeth, J.; Roy, P.D.; Pancost, R.D. 27cal ka biomarker-based record of ecosystem changes from lacustrine sediments of the Chihuahua Desert of Mexico. Quat. Sci. Rev. 2018, 191, 132–143. [Google Scholar] [CrossRef]
- Park, J.; Byrne, R.; Böhnel, H.; Garza, R.M.; Conserva, M. Holocene climate change and human impact, central Mexico: A record based on maar lake pollen and sediment chemistry. Quat. Sci. Rev. 2010, 29, 618–632. [Google Scholar] [CrossRef]
- Sycheva, S.A.; Pogosyan, L.; Sedov, S.; Solleiro-Rebolledo, E.; Golyeva, A.A.; Barceinas-Cruz, H.; Abrosimov, K.N.; Romanenko, K.A. Evidence for stages of landscape evolution in Central Mexico during the late Quaternary from paleosol-pedosediment sequences. Quat. Res. 2022, 107, 159–177. [Google Scholar] [CrossRef]
- Ortega, B.; Vázquez, G.; Caballero, M.; Israde, I.; Lozano-García, S.; Schaaf, P.; Torres, E. Late Pleistocene: Holocene record of environmental changes in Lake Zirahuen, Central Mexico. J. Paleolimnol. 2010, 44, 745–760. [Google Scholar] [CrossRef]
- Ortega-Guerrero, B.; Lozano-García, S.; Herrera-Hernández, D.; Caballero, M.; Beramendi-Orosco, L.; Bernal, J.P.; Torres-Rodríguez, E.; Avendaño-Villeda, D. Lithostratigraphy and physical properties of lacustrine sediments of the last ca. 150 kyr from Chalco basin, central México. J. S. Am. Earth Sci. 2017, 79, 507–524. [Google Scholar] [CrossRef]
- Lamb, A.L.; Gonzalez, S.; Huddart, D.; Metcalfe, S.E.; Vane, C.H.; Pike, A.W. Tepexpan Palaeoindian site, Basin of Mexico: Multi-proxy evidence for environmental change during the late Pleistocene–Late Holocene. Quat. Sci. Rev. 2009, 28, 2000–2016. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology, 4th ed.; Academic Press: New York, NY, USA, 2005. [Google Scholar]
- Roy, P.D.; Jonathan, M.P.; Pérez-Cruz, L.L.; Sánchez-Córdova, M.M.; Quiroz-Jiménez, J.D.; Romero, F.M. A millennial-scale L ate Pleistocene–Holocene palaeoclimatic record from the western Chihuahua Desert, Mexico. Boreas 2012, 41, 707–718. [Google Scholar] [CrossRef]
- Romero-Manzanares, A.; Flores-Flores, J.L.; Luna-Cavazos, M.; Garcìa-Moya, E. Effect of Slope and Aspect on the Associated Flora of Pinyon Pines in Central Mexico. Southwest. Nat. 2012, 57, 452–456. [Google Scholar] [CrossRef]
- Stahle, D.W.; Cook, E.R.; Burnette, D.J.; Villanueva, J.; Cerano, J.; Burns, J.N.; Griffin, D.; Cook, B.I.; Acuña, R.; Torbenson, M.C.; et al. The Mexican Drought Atlas: Tree-ring reconstructions of the soil moisture balance during the late pre-Hispanic, colonial, and modern eras. Quat. Sci. Rev. 2016, 149, 34–60. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit and Depth | 14C Age | Cal. Age | Median Age | Delta 13C | Lab Number | Material |
|---|---|---|---|---|---|---|
| B5-595 cm | 7640 ± 45 | 8520–8360 | 8440 cal. BP | −22.1 | AA 40612 | Humus (soil) |
| B4-630 cm | 7410 ± 65 | 8370–8150 | 8250 cal. BP | −12.1 | OS 17841 | Charcoal |
| B3-800 cm | 8990 ± 60 | 10,250–10,110 | 10,160 cal. BP | −24.0 | (OS 17058) | Charcoal |
| B2-880 cm | 8820 ± 110 | 10,180–9570 | 9890 cal. BP | −23.1 | (AA 33477) | Charcoal |
| A6-920 cm | 9065 ± 80 | 10,430–10,120 | 10,230 cal. BP | −24.0 | (AA 33478) | Charcoal |
| A6-940 cm | 9240 ± 50 | 10,550–10,260 | 10,410 cal. BP | −24.1 | (OS 15416) | Charcoal |
| A6-960 cm | 9100 ± 75 | 10,510–10,150 | 10,270 cal. BP | −23.7 | (AA 33479) | Charcoal |
| A5-9800 cm | 8605 ± 55 | 9680–9480 | 9570 cal. BP | −22.8 | AA 40251 | Humus (soil) |
| A5-1000 cm | 8675 ± 65 | 9830–9530 | 9640 cal. BP | −24.0 | AA 33480 | Charcoal |
| Unit and Depth | 14C Age | Cal. Age | Median Age | Delta 13C | Lab Number | Material |
|---|---|---|---|---|---|---|
| C.6-20 cm | 4205 ± 120 | 5060–4400 | 4740 cal. BP | −19.3 | A 11128 | Humus (soil) |
| C.6-30 cm | 7600 ± 130” | 8660–8150 | 8410 cal. BP | −18.8 | (OS 16100) | Charcoal |
| C.3-125 cm | 5840 ± 150 | 7030–6330 | 6660 cal. BP | −24.5 | A 11122 | Charcoal |
| C.2-240 cm | 5670 ± 60 | 6570–6310 | 6700 cal. BP | −24.6 | OS 14143 | Humus (soil) |
| C.2-250 cm | 5880 ± 60 | 6860–6540 | 6700 cal. BP | −24.6 | OS 14143 | Charcoal |
| Unit and Depth | 14C Age | Cal. Age | Median Age | Delta 13C | Lab Number | Material |
|---|---|---|---|---|---|---|
| G1-30 cm | 370 ± 55 | 510–310 | 420 cal. BP | −25.3 | AA 33471 | Charcoal |
| G1-45 cm | 565 ± 45 | 650–520 | 590 cal. BP | −24.9 | OS 15517 | Charcoal |
| F2-60 cm | 610 ± 80 | 710–510 | 610 cal. BP | −23.5 | A 11116 | Humus (soil) |
| F2-80 cm | 1140 ± 35 | 1160–970 | 1040 cal. BP | −25.1 | OS 15412 | Charcoal |
| F2-95 cm | 1210 ± 50 | 1240–1030 | 1140 cal. BP | −22.6 | TX 9153 | Humus (soil) |
| F1-155 cm | 1880 ± 75 | 1990–1680 | 1820 cal. BP | −24.0 | (0S 15413) | Charcoal |
| E5-170 cm | 1620 ± 65 | 1660–1370 | 1520 cal. BP | −24.3 | A 11117 | Humus (soil) |
| E5-235 cm | 1670 ± 75 | 1760–1410 | 1580 cal. BP | −24.9 | A 11118 | Humus (soil) |
| E.1-295 cm | 1550 ± 40 | 1530–1360 | 1440 cal. BP | −23.9 | (TX 9170) | Humus (soil) |
| D11-320 cm | 2280 ± 60 | 2410–2130 | 2300 cal. BP | −23.1 | TX 9150 | Humus (soil) |
| D5-390 cm | 2795± 40 | 3000–2820 | 2900 cal. BP | −25.2 | AA 40250 | Humus (soil) |
| D3-470 cm | 2820 ± 60 | 3110–2810 | 2940 cal. BP | −24.3 | TX 9154 | Humus (soil) |
| DO-515 cm | 3800 ± 60 | 4370–4010 | 4190 cal. BP | −17.9 | TX 9151 | Humus (soil) |
| Y. BP | Zonal Vegetation Elements | Azonal Vegetation Elements |
|---|---|---|
| 10,000–8875 | Mixed-oak mesophytic woodland in lowlands and piedmont, and downslope migration of pine from uplands | Perennial aquatics in the channel, and ash and elm in floodplain |
| 8860–7900 | Xerophytes in lowlands, mixed-oak scrub woodland in piedmont, and pine uplands | Dry floodplain with ash and elm at riparian verges |
| 6900–6150 | Xerophytes in lowlands and piedmont (mixed with scrub woodland), and pine uplands | Dry floodplain with grasses and forbs |
| 5750–5650 | Xerophytes in lowlands, mixed-oak scrub woodland in piedmont, and downslope migration of pine | Telmatic-influenced floodplain with cattails |
| 5250–4600 | Xerophytes in lowlands and piedmont (mixed with scrub woodland), and pine uplands | Dry floodplain with grasses and forbs, minor telmatics influence |
| ~4190 | Xerophytes in lowlands and piedmont (mixed with scrub woodland), and pine uplands | Dry floodplain with composites |
| ~2900 | Mixed-oak scrub woodland in lowlands and piedmont, and downslope migration of pine | Minor telmatics in dry floodplain with alder and pecan |
| ~2800 | Mixed-oak scrub woodland in lowlands and piedmont, and pine uplands | Telmatic-influenced floodplain with ash and elm at riparian verges |
| 2800–2600 | Xerophytes emergent in lowlands, mixed-oak scrub woodland in piedmont, and pine uplands | Dry floodplain with ash, elm, and pecan, and extensive Ephedra |
| 2600–2150 | Xerophytes in lowlands, mixed-oak scrub woodland in piedmont, and pine uplands | Dry floodplain with composites |
| 1900–1450 | Xerophytes with Artemisia in lowlands, mixed-oak scrub woodland in piedmont, and pinyon pine uplands | Extensive Ephedra on gravels and dry floodplain |
| 400–0 | Xerophytes in lowlands, mixed-oak scrub woodland in piedmont, and pinyon pine uplands | Dry channel (Spanish Colonial dam construction) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albert, B.M. Chihuahuan Desert Vegetation Development during the Past 10,000 Years According to Pollen and Sediment Data at Upper Arroyo, Saltillo, Mexico. Quaternary 2023, 6, 15. https://doi.org/10.3390/quat6010015
Albert BM. Chihuahuan Desert Vegetation Development during the Past 10,000 Years According to Pollen and Sediment Data at Upper Arroyo, Saltillo, Mexico. Quaternary. 2023; 6(1):15. https://doi.org/10.3390/quat6010015
Chicago/Turabian StyleAlbert, Bruce M. 2023. "Chihuahuan Desert Vegetation Development during the Past 10,000 Years According to Pollen and Sediment Data at Upper Arroyo, Saltillo, Mexico" Quaternary 6, no. 1: 15. https://doi.org/10.3390/quat6010015
APA StyleAlbert, B. M. (2023). Chihuahuan Desert Vegetation Development during the Past 10,000 Years According to Pollen and Sediment Data at Upper Arroyo, Saltillo, Mexico. Quaternary, 6(1), 15. https://doi.org/10.3390/quat6010015